
Ascorbic Acid/Retinol and/or Inflammatory Stimuli’s Effect on Proliferation/Differentiation Properties and Transcriptomics of Gingival Stem/Progenitor Cells


 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. G-MSCs’ Isolation, Characterization and Multilineage Differentiation
2.2. Experimental Groups
2.3. G-MSCs’ mRNA Expression
2.4. ELISA
2.5. mRNA Next-Generation Sequencing
2.6. CFUs and Cellular Proliferation
2.7. Multilineage Potential of Stimulated G-MSCs
2.8. Statistical Analysis
3. Results
3.1. Characterization of G-MSCs
3.2. Stemness Markers’ Expression
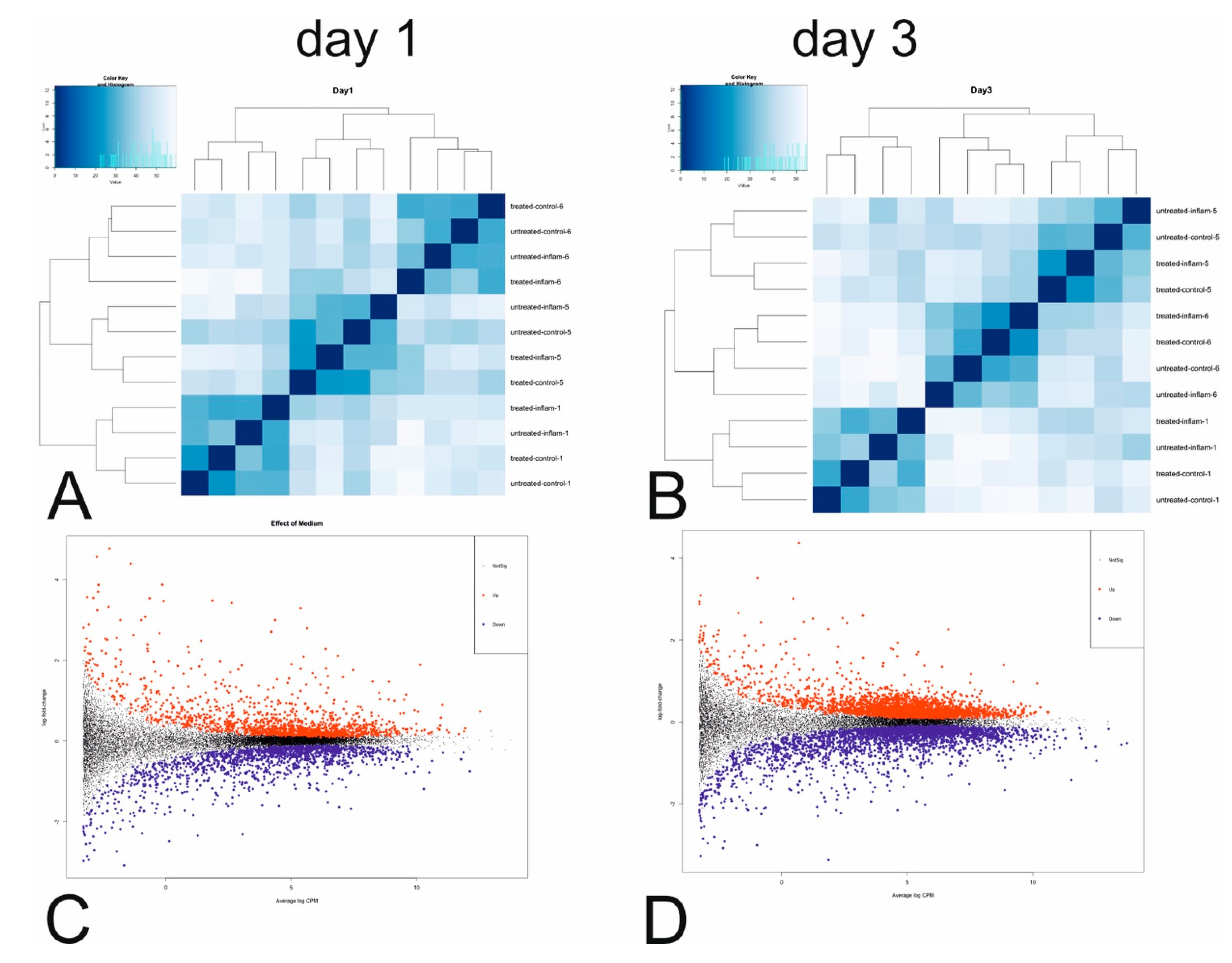
3.3. mRNA Next Generation Sequencing
3.4. Intracellular β-Catenin
3.5. CFUs and Cellular Proliferation
3.6. Stimulated G-MCSs’ Multilineage Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, S.; Yu, N.; Arce, R.M. Periodontal inflammation: Integrating genes and dysbiosis. Periodontology 2020, 82, 129–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papapanou, P.N.; Susin, C. Periodontitis epidemiology: Is periodontitis under-recognized, over-diagnosed, or both? Periodontology 2017, 75, 45–51. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, K.M.F.; Elahmady, M.; Adawi, Z.; Aboushadi, N.; Elnaggar, A.; Eid, M.; Hamdy, N.; Sanaa, D.; Dörfer, C.E. The periodontal stem/progenitor cell inflammatory-regenerative cross talk: A new perspective. J. Periodontal Res. 2019, 54, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Shi, S.; Liu, Y.; Uyanne, J.; Shi, Y.; Shi, S.; Le, A.D. Mesenchymal Stem Cells Derived from Human Gingiva Are Capable of Immunomodulatory Functions and Ameliorate Inflammation-Related Tissue Destruction in Experimental Colitis. J. Immunol. 2009, 183, 7787–7798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawzy-El-Sayed, K.; Mekhemar, M.; Adam-Klages, S.; Kabelitz, D.; Dorfer, C. TlR expression profile of human gingival margin-derived stem progenitor cells. Med. Oral Patol. Oral Cir. Bucal. 2016, 21, e30–e38. [Google Scholar] [CrossRef]
- Mekhemar, M.K.; Adam-Klages, S.; Kabelitz, D.; Dörfer, C.E.; El-Sayed, K.F. TLR-induced immunomodulatory cytokine expression by human gingival stem/progenitor cells. Cell. Immunol. 2018, 326, 60–67. [Google Scholar] [CrossRef]
- Zhou, L.; Dörfer, C.E.; Chen, L.; El-Sayed, K.M.F. Porphyromonas gingivalislipopolysaccharides affect gingival stem/progenitor cells attributes through NF-κB, but not Wnt/β-catenin, pathway. J. Clin. Periodontol. 2017, 44, 1112–1122. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Si, M.; Wang, H.; Mekhemar, M.K.; Dörfer, C.E.; El-Sayed, K.M.F. IL-1/TNF-αInflammatory and Anti-Inflammatory Synchronization Affects Gingival Stem/Progenitor Cells’ Regenerative Attributes. Stem Cells Int. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-A.; Lee, J.-H.; Jin, B.-H.; Bae, K.-H. Association of Some Vitamins and Minerals with Periodontitis in a Nationally Representative Sample of Korean Young Adults. Biol. Trace Element Res. 2017, 178, 171–179. [Google Scholar] [CrossRef]
- Linden, G.J.; McClean, K.M.; Woodside, J.V.; Patterson, C.C.; Evans, A.; Young, I.S.; Kee, F. Antioxidants and periodontitis in 60-70-year-old men. J. Clin. Periodontol. 2009, 36, 843–849. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Jin, Y.; Gao, H.; Lin, X. Oral Administration of All-TransRetinoic Acid Suppresses Experimental Periodontitis by Modulating the Th17/Treg Imbalance. J. Periodontol. 2014, 85, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Grossi, S.G.; Dunford, R.G.; Ho, A.W.; Trevisan, M.; Genco, R.J. Dietary Vitamin C and the Risk for Periodontal Disease. J. Periodontol. 2000, 71, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Chitsazi, M.; Faramarzie, M.; Sadighi, M.; Shirmohammadi, A.; Hashemzadeh, A. Effects of adjective use of melatonin and vitamin C in the treatment of chronic periodontitis: A randomized clinical trial. J. Dent. Res. Dent. Clin. Dent. Prospect. 2017, 11, 236–240. [Google Scholar]
- Sulaiman, A.E.A.; Shehadeh, R.M. Assessment of Total Antioxidant Capacity and the Use of Vitamin C in the Treatment of Non-Smokers with Chronic Periodontitis. J. Periodontol. 2010, 81, 1547–1554. [Google Scholar] [CrossRef]
- Bi, Y.; Gong, M.; Zhang, X.; Zhang, X.; Jiang, W.; Zhang, Y.; Chen, J.; Liu, Y.; He, T.-C.; Li, T. Pre-activation of retinoid signaling facilitates neuronal differentiation of mesenchymal stem cells. Dev. Growth Differ. 2010, 52, 419–431. [Google Scholar] [CrossRef]
- Yu, J.; Tu, Y.-K.; Tang, Y.-B.; Cheng, N.-C. Stemness and transdifferentiation of adipose-derived stem cells using l-ascorbic acid 2-phosphate-induced cell sheet formation. Biomaterials 2014, 35, 3516–3526. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-J.; Sun, L.-Y.; Pang, C.-Y. Synergistic Protection of N-Acetylcysteine and Ascorbic Acid 2-Phosphate on Human Mesenchymal Stem Cells Against Mitoptosis, Necroptosis and Apoptosis. Sci. Rep. 2015, 5, 9819. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Han, Z.; Li, Q.; Wu, Y.; Shi, X.; Ai, Z.; Du, J.; Li, W.; Guo, Z.; Zhang, Y. Vitamin C induces a pluripotent state in mouse embryonic stem cells by modulating microRNA expression. FEBS J. 2015, 282, 685–699. [Google Scholar] [CrossRef]
- Esteban, M.A.; Pei, D. Vitamin C improves the quality of somatic cell reprogramming. Nat. Genet. 2012, 44, 366–367. [Google Scholar] [CrossRef]
- Hore, T.A. Modulating epigenetic memory through vitamins and TET: Implications for regenerative medicine and cancer treatment. Epigenomics 2017, 9, 863–871. [Google Scholar] [CrossRef]
- Van Pham, P.; Tran, N.Y.; Phan, N.L.-C.; Vu, N.B.; Phan, N.K. Vitamin C stimulates human gingival stem cell proliferation and expression of pluripotent markers. Vitr. Cell. Dev. Biol. Anim. 2015, 52, 218–227. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, K.M.F.; Paris, S.; Graetz, C.; Kassem, N.; Mekhemar, M.; Ungefroren, H.; Fändrich, F.; E Dorfer, C. Isolation and characterisation of human gingival margin-derived STRO-1/MACS+ and MACS− cell populations. Int. J. Oral Sci. 2015, 7, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidney, L.E.; Kirkham, G.R.; Buttery, L.D. Comparison of Osteogenic Differentiation of Embryonic Stem Cells and Primary Osteoblasts Revealed by Responses to IL-1β, TNF-α, and IFN-γ. Stem Cells Dev. 2014, 23, 605–617. [Google Scholar] [CrossRef] [Green Version]
- Fawzy-El-Sayed, K.M.; Klingebiel, P.; Dörfer, C.E. Toll-like Receptor Expression Profile of Human Dental Pulp Stem/Progenitor Cells. J. Endod. 2016, 42, 413–417. [Google Scholar] [CrossRef]
- Fehrmann, C.; Dörfer, C.E.; El-Sayed, K.M.F. Toll-like Receptor Expression Profile of Human Stem/Progenitor Cells Form the Apical Papilla. J. Endod. 2020, 46, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, X.; Hu, Y.; Wu, J.; Cao, Q.; Chen, S.; Gao, Y. All-trans retinoic acid modulates Wnt3A-induced osteogenic differentiation of mesenchymal stem cells via activating the PI3K/AKT/GSK3β signalling pathway. Mol. Cell. Endocrinol. 2016, 422, 243–253. [Google Scholar] [CrossRef]
- Ewels, P.A.; Peltzer, A.; Fillinger, S.; Patel, H.; Alneberg, J.; Wilm, A.; Garcia, M.U.; Di Tommaso, P.; Nahnsen, S. The nf-core framework for community-curated bioinformatics pipelines. Nat. Biotechnol. 2020, 38, 276–278. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Mahmood, M.; Li, Z.; Casciano, D.; Khodakovskaya, M.V.; Chen, T.; Karmakar, A.; Dervishi, E.; Xu, Y.; Mustafa, T.; Watanabe, F.; et al. Nanostructural materials increase mineralization in bone cells and affect gene expression through miRNA regulation. J. Cell. Mol. Med. 2011, 15, 2297–2306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Ye, X.; Xie, M.; Ye, J. Induction of triglyceride accumulation and mitochondrial maintenance in muscle cells by lactate. Sci. Rep. 2016, 6, 33732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekhemar, M.; Tölle, J.; Dörfer, C.; El-Sayed, K.F. TLR3 ligation affects differentiation and stemness properties of gingival mesenchymal stem/progenitor cells. J. Clin. Periodontol. 2020, 47, 991–1005. [Google Scholar] [CrossRef]
- El-Sayed, K.M.F.; Dörfer, C.E. Animal Models for Periodontal Tissue Engineering: A Knowledge-Generating Process. Tissue Eng. Part C: Methods 2017, 23, 900–925. [Google Scholar] [CrossRef]
- Silva, T.; Garlet, G.; Fukada, S.; Silva, J.S.; Cunha, F.Q. Chemokines in Oral Inflammatory Diseases: Apical Periodontitis and Periodontal Disease. J. Dent. Res. 2007, 86, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Bin Lee, J.; Cha, J.; Choi, E.; Park, S.; Cho, K.S.; Kim, C. Chemokine in inflamed periodontal tissues activates healthy periodontal-ligament stem cell migration. J. Clin. Periodontol. 2017, 44, 530–539. [Google Scholar] [CrossRef]
- Rubin, M.B. Vitamins Wound Healing. Plast. Surg. Nurs. 1984, 4, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Elbehwashy, M.T.; Hosny, M.M.; Elfana, A.; Nawar, A.; El-Sayed, K.F. Clinical and radiographic effects of ascorbic acid-augmented platelet-rich fibrin versus platelet-rich fibrin alone in intra-osseous defects of stage-III periodontitis patients: A randomized controlled clinical trial. Clin. Oral Investig. 2021, 25, 6309–6319. [Google Scholar] [CrossRef]
- Chapple, I.L.C. Reactive oxygen species and antioxidants in inflammatory diseases. J. Clin. Periodontol. 1997, 24, 287–296. [Google Scholar] [CrossRef]
- Alagl, A.; Bhat, S.G. Ascorbic acid: New role of an age-old micronutrient in the management of periodontal disease in older adults. Geriatr. Gerontol. Int. 2014, 15, 241–254. [Google Scholar] [CrossRef]
- Majewicz, J.; Rimbach, G.; Proteggente, A.R.; Lodge, J.K.; Kraemer, K.; Minihane, A.M. Dietary vitamin C down-regulates inflammatory gene expression in apoE4 smokers. Biochem. Biophys. Res. Commun. 2005, 338, 951–955. [Google Scholar] [CrossRef]
- Tomofuji, T.; Ekuni, D.; Sanbe, T.; Irie, K.; Azuma, T.; Maruyama, T.; Tamaki, N.; Murakami, J.; Kokeguchi, S.; Yamamoto, T. Effects of vitamin C intake on gingival oxidative stress in rat periodontitis. Free. Radic. Biol. Med. 2009, 46, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Erkelens, M.N.; Mebius, R.E. Retinoic Acid and Immune Homeostasis: A Balancing Act. Trends Immunol. 2017, 38, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, L.; Hussein, H.; Ghanem, M.; Eissa, N. Retinoic acid-mediated anti-inflammatory responses in equine immune cells stimulated by LPS and allogeneic mesenchymal stem cells. Res. Veter. Sci. 2017, 114, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, L.; Chen, L.; Wang, X.; Wu, H.; Ai, Z.; Du, J.; Liu, Y.; Shi, X.; Wu, Y.; et al. Vitamin C facilitates pluripotent stem cell maintenance by promoting pluripotency gene transcription. Biochimie 2013, 95, 2107–2113. [Google Scholar] [CrossRef] [PubMed]
- Tomar, G.B.; Srivastava, R.K.; Gupta, N.; Barhanpurkar, A.P.; Pote, S.T.; Jhaveri, H.M.; Mishra, G.C.; Wani, M.R. Human gingiva-derived mesenchymal stem cells are superior to bone marrow-derived mesenchymal stem cells for cell therapy in regenerative medicine. Biochem. Biophys. Res. Commun. 2010, 393, 377–383. [Google Scholar] [CrossRef]
- Fournier, B.P.; Ferre, F.C.; Couty, L.; Lataillade, J.-J.; Gourven, M.; Naveau, A.; Coulomb, B.; Lafont, A.; Gogly, B. Multipotent Progenitor Cells in Gingival Connective Tissue. Tissue Eng. Part A 2010, 16, 2891–2899. [Google Scholar] [CrossRef]
- Jin, S.-H.; Lee, J.E.; Yun, J.-H.; Kim, I.; Ko, Y.; Park, J.B. Isolation and characterization of human mesenchymal stem cells from gingival connective tissue. J. Periodontal Res. 2014, 50, 461–467. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, G.; Jianliang, P.; Chen, X.; Pang, J.; Ke, J. Isolation and Multiple Differentiation Potential Assessment of Human Gingival Mesenchymal Stem Cells. Int. J. Mol. Sci. 2014, 15, 20982–20996. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- El-Sayed, K.M.F.; Hein, D.; Dörfer, C.E. Retinol/inflammation affect stemness and differentiation potential of gingival stem/progenitor cells via Wnt/β-catenin. J. Periodontal Res. 2019, 54, 413–423. [Google Scholar] [CrossRef]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, P.R.; Budnik, B.; Slavov, N.; Semrau, S. Dynamic post-transcriptional regulation during embryonic stem cell differentiation. bioRxiv 2017, 123497. [Google Scholar] [CrossRef] [Green Version]
- Reimegård, J.; Tarbier, M.; Danielsson, M.; Schuster, J.; Baskaran, S.; Panagiotou, S.; Dahl, N.; Friedländer, M.R.; Gallant, C.J. A combined approach for single-cell mRNA and intracellular protein expression analysis. Commun. Biol. 2021, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hore, T.A.; von Meyenn, F.; Ravichandran, M.; Bachman, M.; Ficz, G.; Oxley, D.; Santos, F.; Balasubramanian, S.; Jurkowski, T.P.; Reik, W. Retinol and ascorbate drive erasure of epigenetic memory and enhance reprogramming to naïve pluripotency by complementary mechanisms. Proc. Natl. Acad. Sci. USA 2016, 113, 12202–12207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, M.; Ortolano, N.A.; Romero-Morales, A.I.; Gama, V. Wnt Signaling and Its Impact on Mitochondrial and Cell Cycle Dynamics in Pluripotent Stem Cells. Genes 2018, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; Liu, Y.; Ye, R.; Zhu, B.; Zhu, Y.; Liu, X.; Hu, C.-H.; Luo, H.; Zhang, Y.; Ding, Y.; et al. GSK3β is a checkpoint for TNF-α-mediated impaired osteogenic differentiation of mesenchymal stem cells in inflammatory microenvironments. Biochim. Biophys. Acta (BBA) Gen. Subj. 2013, 1830, 5119–5129. [Google Scholar] [CrossRef]
- Lehnert, K.; Ni, J.; Leung, E.; Gough, S.; Morris, C.; Liu, D.; Wang, S.-X.; Langley, R.; Krissansen, G. The integrin α10 subunit: Expression pattern, partial gene structure, and chromosomal localization. Cytogenet. Genome Res. 1999, 87, 238–244. [Google Scholar] [CrossRef]
- Bosman, F.T.; Stamenkovic, I. Functional structure and composition of the extracellular matrix. J. Pathol. 2003, 200, 423–428. [Google Scholar] [CrossRef]
- Stennard, F.; Holloway, A.F.; Hamilton, J.; West, A.K. Characterisation of six additional human metallothionein genes. Biochim. Biophys. Acta (BBA) Gene Struct. Expr. 1994, 1218, 357–365. [Google Scholar] [CrossRef]
- Petit, V.; Thiery, J.-P. Focal adhesions: Structure and dynamics. Biol. Cell 2000, 92, 477–494. [Google Scholar] [CrossRef]
- Yoo, H.G.; Chang, I.-Y.; Pyo, H.K.; Kang, Y.J.; Lee, S.H.; Kwon, O.; Cho, K.H.; Eun, H.C.; Kim, K.H. The Additive Effects of Minoxidil and Retinol on Human Hair Growth in Vitro. Biol. Pharm. Bull. 2007, 30, 21–26. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, K.M.F.; Nguyen, N.; Dörfer, C.E. Ascorbic Acid, Inflammatory Cytokines (IL-1β/TNF-α/IFN-γ), or Their Combination’s Effect on Stemness, Proliferation, and Differentiation of Gingival Mesenchymal Stem/Progenitor Cells. Stem Cells Int. 2020, 2020, 1–14. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, W.-K.; Sung, Y.K.; Kwack, M.H.; Song, S.Y.; Choi, J.-S.; Park, S.G.; Yi, T.; Lee, H.-J.; Kim, D.-D.; et al. The Molecular Mechanism Underlying the Proliferating and Preconditioning Effect of Vitamin C on Adipose-Derived Stem Cells. Stem Cells Dev. 2014, 23, 1364–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, L.; Kikuiri, T.; Akiyama, K.; Chen, C.; Xu, X.; Yang, R.; Chen, W.; Wang, S.; Shi, S. Mesenchymal stem cell–based tissue regeneration is governed by recipient T lymphocytes via IFN-γ and TNF-α. Nat. Med. 2011, 17, 1594–1601. [Google Scholar] [CrossRef]
- Carinci, F.; Pezzetti, F.; Spina, A.M.; Palmieri, A.; Laino, G.; De Rosa, A.; Farina, E.; Illiano, F.; Stabellini, G.; Perrotti, V.; et al. Effect of Vitamin C on pre-osteoblast gene expression. Arch. Oral Biol. 2005, 50, 481–496. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Iwasaki, K.; Komaki, M.; Ishikawa, I. Role of Ascorbic Acid in Periodontal Ligament Cell Differentiation. J. Periodontol. 2004, 75, 709–716. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor’s Number | Gender | Age |
|---|---|---|
| 1 | Male | 18 |
| 2 | Female | 20 |
| 3 | Female | 19 |
| 4 | Male | 22 |
| 5 | Male | 20 |
| Gene | Gene Symbol | Accession ID | Assay ID |
|---|---|---|---|
| RUNX2 | RUNX2 H. sapiens | ENST00000359524 | 113380 |
| ACAN | ACAN H. sapiens | ENST00000439576 | 138057 |
| ALP | ALP H. sapiens | ENST00000374840 | 103448 |
| LPL | LPL H. sapiens | ENST00000311322 | 113230 |
| NANOG | NANOG H. sapiens | ENST00000229307 | 148147 |
| OCT4A | OCT4 H. sapiens | ENST00000259915 | 113034 |
| PGK1 | PGK1 H. sapiens | ENST00000373316 | 102083 |
| PPARɣ | PPARɣ H. sapiens | ENST00000287820 | 110607 |
| SOX2 | SOX2 H. sapiens | ENST00000325404 | 111867 |
| Ensemble | Entrez ID | Gene Name | LogFC | p-Value | FDR | ||||
|---|---|---|---|---|---|---|---|---|---|
| Medium | Treatment | ||||||||
| Day 1 | Treatment + medium | Gen 1 | ENSG00000105825 | 7980 | TFPI1 | 1.46 | 1.82 × 10−13 | 1.41 × 10−9 | |
| Gen 2 | ENSG00000134363 | 10468 | FST | −0.43 | 5.48 × 10−13 | 2.0 4× 10−9 | |||
| Gen 3 | ENSG00000096060 | 2289 | FKBP5 | 1.54 | 6.66 × 10−13 | 2.04 × 10−9 | |||
| Medium | Gen 1 | ENSG00000105825 | 7980 | TFPI2 | 1.98 | 5.85 × 10−14 | 9.06 × 10−10 | ||
| Gen 2 | ENSG00000163735 | 6374 | CXCL5 | 3.86 | 5.63 × 10−12 | 4.36 × 10−8 | |||
| Gen 3 | ENSG00000163131 | 1520 | CTSS | 2.27 | 8.82 × 10−12 | 4.56 × 10−8 | |||
| Treatment | Gen 1 | ENSG00000096060 | 2289 | FKBP5 | 2.00 | 1.29 × 10−13 | 2.00 × 10−9 | ||
| Gen 2 | ENSG00000134363 | 10468 | FST | −1.52 | 1.74 × 10−12 | 1.25 × 10−8 | |||
| Gen 3 | ENSG00000169715 | 4493 | MT1E | 1.33 | 2.42 × 10−12 | 1.25 × 10−8 | |||
| Day 3 | Treatment + medium | Gen 1 | ENSG00000096060 | 2289 | FKBP5 | 2.00 | 1.67 × 10−12 | 1.21 × 10−8 | |
| Gen 2 | ENSG00000135069 | 29968 | PSAT1 | 1.17 | 1.18 × 10−12 | 1.22 × 10−8 | |||
| Gen 3 | ENSG00000134363 | 10468 | ASNS | 0.38 | 2.39 × 10−11 | 1.22 × 10−8 | |||
| Medium | Gen 1 | ENSG00000019582 | 972 | CD74 | 1.89 | 3.20 × 10−12 | 4.88 × 10−8 | ||
| Gen 2 | ENSG00000163131 | 1520 | CTSS | 1.53 | 9.24 × 10−11 | 7.05 × 10−7 | |||
| Gen 3 | ENSG00000221852 | 83895 | KRTAP1-5 | 1.12 | 4.27 × 10−9 | 1.83 × 10−5 | |||
| Treatment | Gen 1 | ENSG00000096060 | 2289 | FKBP5 | 2.52 | 3.32 × 10−14 | 5.05 × 10−10 | ||
| Gen 2 | ENSG00000134363 | 10468 | FST | −1.83 | 2.04 × 10−11 | 1.55 × 10−7 | |||
| Gen 3 | ENSG00000116991 | 57568 | SIPA1L2 | −1.40 | 4.48 × 10−11 | 2.28 × 10−7 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fawzy El-Sayed, K.M.; Bittner, A.; Schlicht, K.; Mekhemar, M.; Enthammer, K.; Höppner, M.; Es-Souni, M.; Schulz, J.; Laudes, M.; Graetz, C.; et al. Ascorbic Acid/Retinol and/or Inflammatory Stimuli’s Effect on Proliferation/Differentiation Properties and Transcriptomics of Gingival Stem/Progenitor Cells. Cells 2021, 10, 3310. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123310
Fawzy El-Sayed KM, Bittner A, Schlicht K, Mekhemar M, Enthammer K, Höppner M, Es-Souni M, Schulz J, Laudes M, Graetz C, et al. Ascorbic Acid/Retinol and/or Inflammatory Stimuli’s Effect on Proliferation/Differentiation Properties and Transcriptomics of Gingival Stem/Progenitor Cells. Cells. 2021; 10(12):3310. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123310
Chicago/Turabian StyleFawzy El-Sayed, Karim M., Amira Bittner, Kristina Schlicht, Mohamed Mekhemar, Kim Enthammer, Marc Höppner, Martha Es-Souni, Juliane Schulz, Matthias Laudes, Christian Graetz, and et al. 2021. "Ascorbic Acid/Retinol and/or Inflammatory Stimuli’s Effect on Proliferation/Differentiation Properties and Transcriptomics of Gingival Stem/Progenitor Cells" Cells 10, no. 12: 3310. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123310