
HOPX Plays a Critical Role in Antiretroviral Drugs Induced Epigenetic Modification and Cardiac Hypertrophy

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells Culture and ARVs Treatment
2.2. Global Transcriptomic Profiling
2.3. Functional Annotation
2.4. RNA Isolation and Expression Analysis by Quantitative Real-Time PCR
2.5. Protein Extraction and Western Blot Analysis
2.6. siRNA-Mediated Knockdown of HOPX
2.7. Immunofluorescence Staining
2.8. Immunohistochemistry
2.9. HDAC Activity Assay
2.10. Data Analysis and Statistical Procedures
3. Results
3.1. ARVs Treatment Cause Pathological Gene Expression in Cardiomyocytes
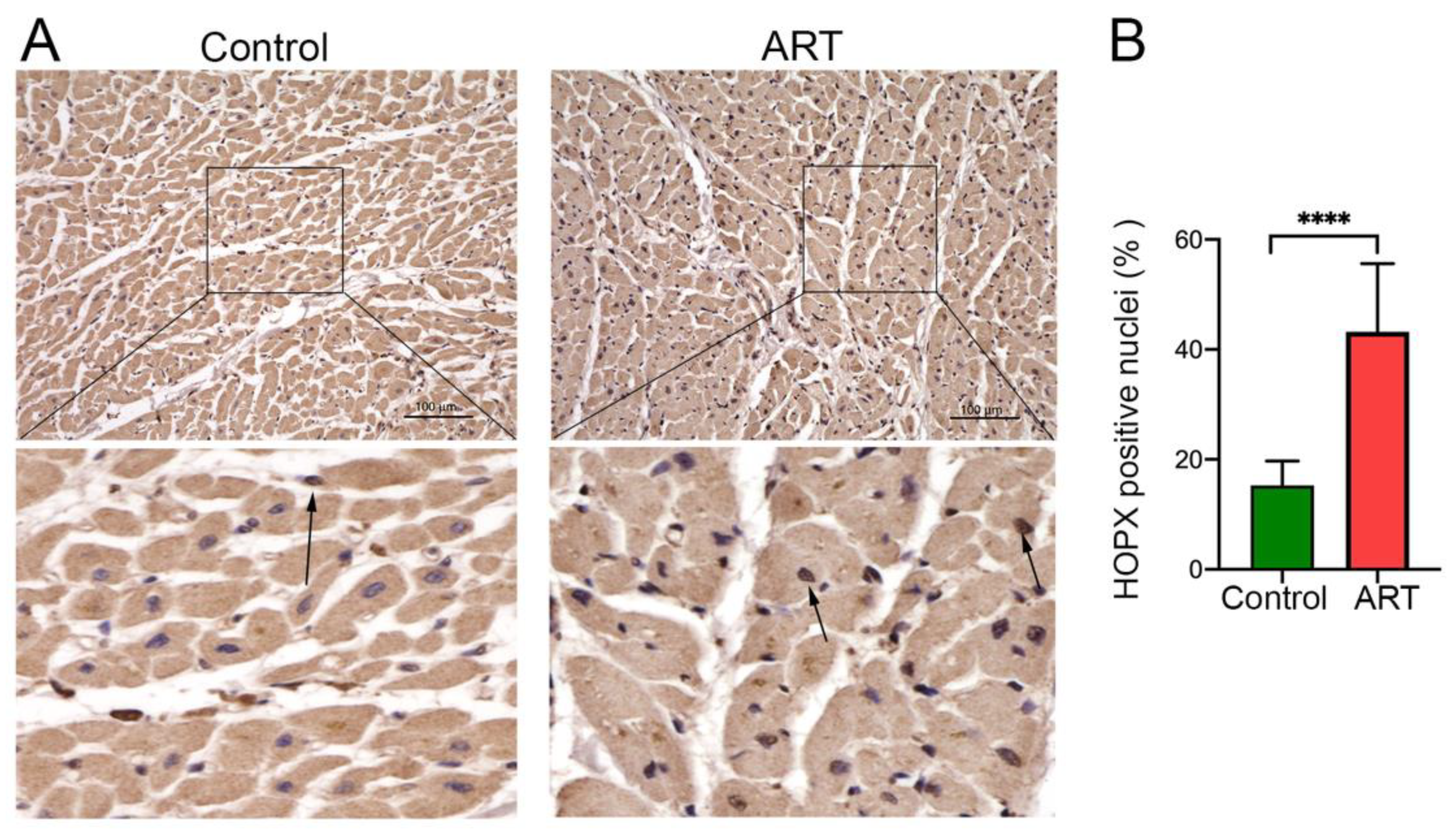
3.2. HOPX Is Upregulated in Cardiomyocytes and in Cardiac Tissue of HIV Patients after ARV Treatment
3.3. HOPX Differentially Localized in the ART-Treated Cardiac Tissue
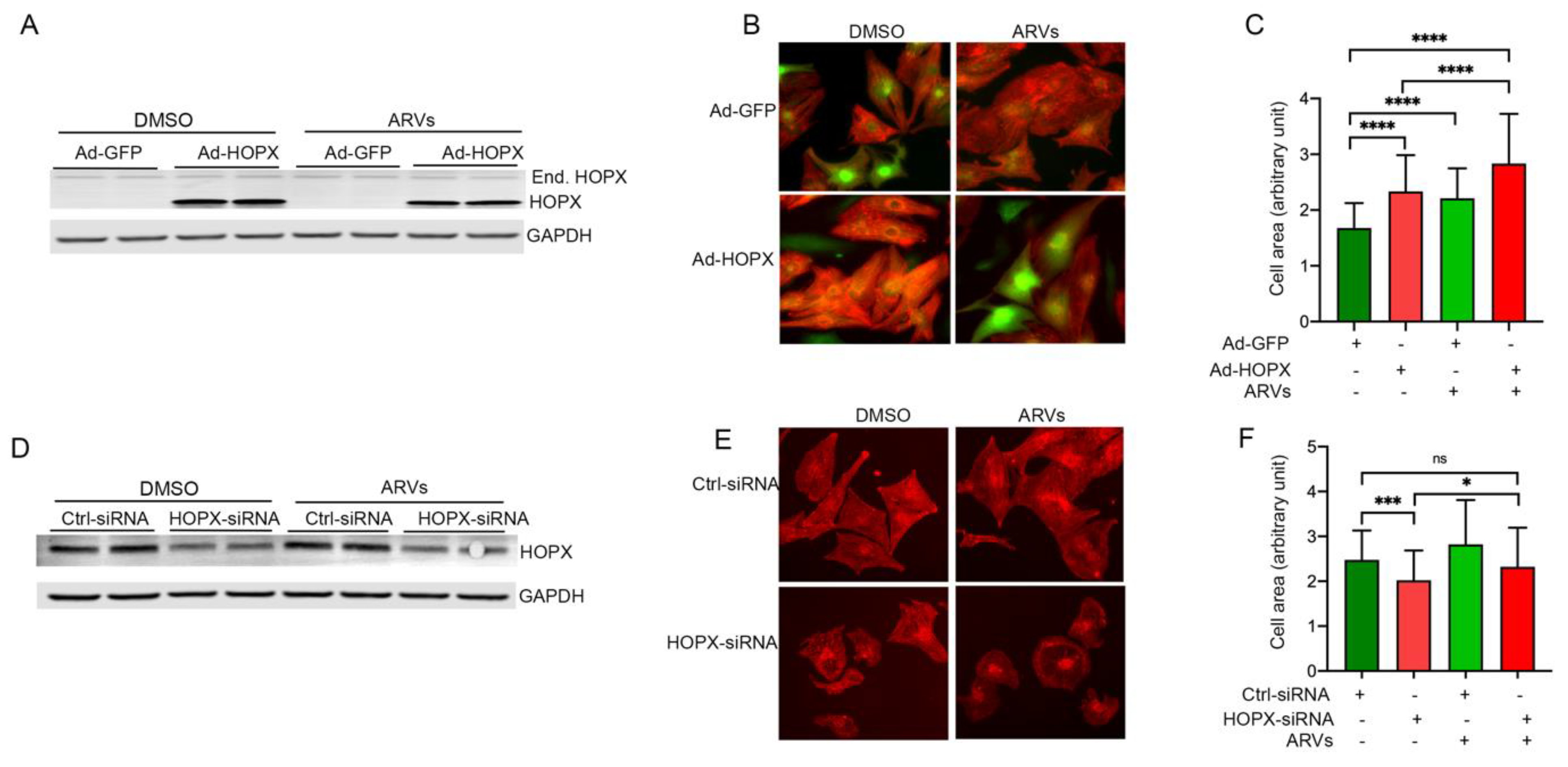
3.4. HOPX Regulates Cardiomyocyte Hypertrophy
3.5. HOPX Is Critically Involved in Epigenetic Modification of Histone 3 during ARV Treatment
3.6. HOPX Is Important to Maintain an Optimum Acetylation Level in the Histone 3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. The Global HIV/AIDS Epidemic. 2021. Available online: https://www.hiv.gov/hiv-basics/overview/data-and-trends/global-statistics (accessed on 1 November 2021).
- WHO. Consolidated Guidelines on the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection: Recommendations for a Public Health Approach; WHO Guidelines Approved by the Guidelines Review Committee: Geneva, Switzerland, 2013. [Google Scholar]
- Buregyeya, E.; Naigino, R.; Mukose, A.; Makumbi, F.; Esiru, G.; Arinaitwe, J.; Musinguzi, J.; Wanyenze, R.K. Facilitators and barriers to uptake and adherence to lifelong antiretroviral therapy among HIV infected pregnant women in Uganda: A qualitative study. BMC Pregnancy Childbirth 2017, 17, 94. [Google Scholar] [CrossRef] [Green Version]
- Feinstein, M.J.; Bahiru, E.; Achenbach, C.; Longenecker, C.T.; Hsue, P.; So-Armah, K.; Freiberg, M.S.; Lloyd-Jones, D. Patterns of Cardiovascular Mortality for HIV-Infected Adults in the United States: 1999 to 2013. Am. J. Cardiol. 2016, 117, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Palella, F.J., Jr.; Baker, R.K.; Moorman, A.C.; Chmiel, J.S.; Wood, K.C.; Brooks, J.T.; Holmberg, S.D. Mortality in the highly active antiretroviral therapy era: Changing causes of death and disease in the HIV outpatient study. J. Acquir Immun. Defic. Syndr. 2006, 43, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.S.; Stelzle, D.; Lee, K.K.; Beck, E.J.; Alam, S.; Clifford, S.; Longenecker, C.T.; Strachan, F.; Bagchi, S.; Whiteley, W.; et al. Global Burden of Atherosclerotic Cardiovascular Disease in People Living with HIV: Systematic Review and Meta-Analysis. Circulation 2018, 138, 1100–1112. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.M.; Wu, J.; Jansson, J.; Wilson, D.P. Relative risk of cardiovascular disease among people living with HIV: A systematic review and meta-analysis. HIV Med. 2012, 13, 453–468. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Lewin, S.R.; Havlir, D.V. The end of AIDS: HIV infection as a chronic disease. Lancet 2013, 382, 1525–1533. [Google Scholar] [CrossRef] [Green Version]
- Hsue, P.Y.; Waters, D.D. HIV infection and coronary heart disease: Mechanisms and management. Nat. Rev. Cardiol. 2019, 16, 745–759. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.L.; Whitlock, G.; Milinkovic, A.; Moyle, G. HIV integrase inhibitors: A new era in the treatment of HIV. Expert Opin. Pharmacother. 2015, 16, 1313–1324. [Google Scholar] [CrossRef]
- Iwuji, C.C.; Churchill, D.; Bremner, S.; Perry, N.; To, Y.; Lambert, D.; Bruce, C.; Waters, L.; Orkin, C.; Geretti, A.M. A phase IV Randomised, Open-Label Pilot Study to Evaluate Switching from Protease-Inhibitor Based Regimen to Bictegravir/Emtricitabine/Tenofovir Alafenamide Single Tablet Regimen in Integrase Inhibitor-Naive, Virologically Suppressed HIV-1 Infected Adults Harbouring Drug Resistance Mutations (PIBIK Study): Study Protocol for a Randomised Trial. BMC Infect. Dis. 2020, 20, 524. [Google Scholar]
- Reyskens, K.M.; Essop, M.F. HIV protease inhibitors and onset of cardiovascular diseases: A central role for oxidative stress and dysregulation of the ubiquitin-proteasome system. Biochim. Biophys. Acta. 2014, 1842, 256–268. [Google Scholar] [CrossRef] [Green Version]
- Stockdale, A.; Saunders, M.; Boyd, M.; Bonnett, L.; Johnston, V.; Wandeler, G.; Schoffelen, A.F.; Ciaffi, L.; Stafford, K.; Collier, A.C.; et al. Effectiveness of Protease Inhibitor/Nucleos(t)ide Reverse Transcriptase Inhibitor-Based Second-line Antiretroviral Therapy for the Treatment of Human Immunodeficiency Virus Type 1 Infection in Sub-Saharan Africa: A Systematic Review and Meta-analysis. Clin. Infect. Dis. 2018, 66, 1846–1857. [Google Scholar] [CrossRef] [Green Version]
- Smit, M.; Brinkman, K.; Geerlings, S.; Smit, C.; Thyagarajan, K.; van Sighem, A.; de Wolf, F.; Hallett, T. Future challenges for clinical care of an ageing population infected with HIV: A modelling study. Lancet Infect. Dis. 2015, 15, 810–818. [Google Scholar] [CrossRef] [Green Version]
- Friismoller, N.; Sabin, C.A.; Weber, R.S.; Monforte, A.D.; Elsadr, W.; Reiss, P.; Thiebaut, R.; Morfeldt, L.; De Wit, S.; Pradier, C.; et al. Combination antiretroviral therapy and the risk of myocardial infarction. N. Engl. J. Med. 2003, 349, 1993–2003. [Google Scholar]
- Grinspoon, S.; Carr, A. Cardiovascular risk and body-fat abnormalities in HIV-infected adults. N. Engl. J. Med. 2005, 352, 48–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondy, K.E.; Gottdiener, J.; Overton, E.T.; Henry, K.; Bush, T.; Conley, L.; Hammer, J.; Carpenter, C.C.; Kojic, E.; Patel, P.; et al. High Prevalence of Echocardiographic Abnormalities among HIV-infected Persons in the Era of Highly Active Antiretroviral Therapy. Clin. Infect. Dis. 2011, 52, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Zareba, K.M.; Lavigne, J.E.; Lipshultz, S.E. Cardiovascular effects of HAART in infants and children of HIV-infected mothers. Cardiovasc. Toxicol. 2004, 4, 271–279. [Google Scholar] [CrossRef]
- Butler, J.; Kalogeropoulos, A.P.; Anstrom, K.J.; Hsue, P.Y.; Kim, R.J.; Scherzer, R.; Shah, S.J.; Shah, S.H.; Velazquez, E.J.; Hernandez, A.F.; et al. Diastolic Dysfunction in Individuals With Human Immunodeficiency Virus Infection: Literature Review, Rationale and Design of the Characterizing Heart Function on Antiretroviral Therapy (CHART) Study. J. Card. Fail. 2018, 24, 255–265. [Google Scholar] [CrossRef]
- Arceci, R.J.; King, A.A.; Simon, M.C.; Orkin, S.H.; Wilson, D.B. Mouse GATA-4: A retinoic acid-inducible GATA-binding transcription factor expressed in endodermally derived tissues and heart. Mol. Cell. Biol. 1993, 13, 2235–2246. [Google Scholar] [PubMed] [Green Version]
- Kelley, C.; Blumberg, H.; Zon, L.I.; Evans, T. GATA-4 is a novel transcription factor expressed in endocardium of the developing heart. Development 1993, 118, 817–827. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Asanoma, K.; Takao, T.; Kato, K.; Wake, N. Homeobox gene HOPX is epigenetically silenced in human uterine endometrial cancer and suppresses estrogen-stimulated proliferation of cancer cells by inhibiting serum response factor. Int. J. Cancer 2009, 124, 2577–2588. [Google Scholar] [CrossRef] [PubMed]
- Haberland, M.; Montgomery, R.L.; Olson, E.N. The many roles of histone deacetylases in development and physiology: Implications for disease and therapy. Nat. Rev. Genet. 2009, 10, 32–42. [Google Scholar] [CrossRef]
- Trivedi, C.M.; Zhu, W.; Wang, Q.; Jia, C.; Kee, H.J.; Li, L.; Hannenhalli, S.; Epstein, J.A. Hopx and Hdac2 interact to modulate Gata4 acetylation and embryonic cardiac myocyte proliferation. Dev. Cell. 2010, 19, 450–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haq, S.; Choukroun, G.; Kang, Z.B.; Ranu, H.; Matsui, T.; Rosenzweig, A.; Haq, S.; Choukroun, G.; Kang, Z.B.; Ranu, H.; et al. Glycogen synthase kinase-3beta is a negative regulator of cardiomyocyte hypertrophy. J. Cell Biol. 2000, 151, 117–130. [Google Scholar] [CrossRef]
- Minzi, O.; Mugoyela, V.; Gustafsson, L. Correlation between lamivudine plasma concentrations and patient self-reported adherence to antiretroviral treatment in experienced HIV patients. Ther. Clin. Risk Manag. 2011, 7, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. Clusterprofiler: An R package for comparing biological themes among gene clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Kashyap, S.; Mukker, A.; Gupta, D.; Datta, P.K.; Rappaport, J.; Jacobson, J.M.; Ebert, S.N.; Gupta, M.K. Antiretroviral Drugs Regulate Epigenetic Modification of Cardiac Cells Through Modulation of H3K9 and H3K27 Acetylation. Front. Cardiovasc. Med. 2021, 8, 634774. [Google Scholar] [CrossRef]
- Feinstein, M.J.; Steverson, A.B.; Ning, H.; Pawlowski, A.E.; Schneider, D.; Ahmad, F.S.; Sanders, J.M.; Sinha, A.; Nance, R.M.; Achenbach, C.J.; et al. Adjudicated Heart Failure in HIV-Infected and Uninfected Men and Women. J. Am. Heart Assoc. 2018, 7, e009985. [Google Scholar] [CrossRef] [Green Version]
- WHO. Consolidated Guidelines on the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection: Recommendations for a Public Health Approach—2nd ed. 2016. Available online: https://apps.who.int/iris (accessed on 1 November 2021).
- Takimoto, E.; Kass, D.A. Role of oxidative stress in cardiac hypertrophy and remodeling. Hypertension 2007, 49, 241–248. [Google Scholar] [CrossRef]
- Barbaro, G.; Barbarini, G.; Di Lorenzo, G. Early impairment of systolic and diastolic function in asymptomatic HIV-positive patients: A multicenter echocardiographic and echo-Doppler study. AIDS Res. Hum. Retrovir. 1996, 12, 1559–1563. [Google Scholar] [CrossRef] [PubMed]
- Lipshultz, S.E.; Easley, K.; Orav, E.J.; Kaplan, S.; Starc, T.J.; Bricker, J.T.; Lai, W.W.; Moodie, D.S.; McIntosh, K.; Schluchter, M.D.; et al. Left ventricular structure and function in children infected with human immunodeficiency virus: The prospective P2C2 HIV Multicenter Study. Pediatric Pulmonary and Cardiac Complications of Vertically Transmitted HIV Infection (P2C2 HIV) Study Group. Circulation 1998, 97, 1246–1256. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Garcia, T.; Sobrino, J.M.; Pujol, E.; Galvez, J.; Benitez, E.; Giron-Gonzalez, J.A. Ventricular mass and diastolic function in patients infected by the human immunodeficiency virus. Heart 2000, 84, 620–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugliese, A.; Isnardi, D.; Saini, A.; Scarabelli, T.; Raddino, R.; Torre, D. Impact of highly active antiretroviral therapy in HIV-positive patients with cardiac involvement. J. Infect. 2000, 40, 282–284. [Google Scholar] [CrossRef]
- Samaan, S.A.; Foster, A.; Raizada, V.; Schade, D.S.; Koster, F.; Crawford, M.H. Myocardial atrophy in acquired immunodeficiency syndrome--associated wasting. Am. Heart J. 1995, 130, 823–827. [Google Scholar] [CrossRef]
- Heineke, J.; Molkentin, J.D. Regulation of cardiac hypertrophy by intracellular signalling pathways. Nat. Rev. Mol. Cell Biol. 2006, 7, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Kook, H.; Milewski, R.; Gitler, A.D.; Lu, M.M.; Li, J.; Nazarian, R.; Schnepp, R.; Jen, K.-Y.; Biben, C.; et al. Hop is an unusual homeobox gene that modulates cardiac development. Cell 2002, 110, 713–723. [Google Scholar] [CrossRef] [Green Version]
- Shin, C.H.; Liu, Z.-P.; Passier, R.; Zhang, C.-L.; Wang, D.-Z.; Harris, T.M.; Yamagishi, H.; Richardson, J.A.; Childs, G.; Olson, E.N. Modulation of cardiac growth and development by HOP, an unusual homeodomain protein. Cell 2002, 110, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Kook, H.; Lepore, J.J.; Gitler, A.D.; Lu, M.M.; Yung, W.W.-M.; Mackay, J.; Zhou, R.; Ferrari, V.; Gruber, P.; Epstein, J.A. Cardiac hypertrophy and histone deacetylase-dependent transcriptional repression mediated by the atypical homeodomain protein Hop. J. Clin. Investig. 2003, 112, 863–871. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashyap, S.; Rabbani, M.; de Lima, I.; Kondrachuk, O.; Patel, R.; Shafiei, M.S.; Mukker, A.; Rajakumar, A.; Gupta, M.K. HOPX Plays a Critical Role in Antiretroviral Drugs Induced Epigenetic Modification and Cardiac Hypertrophy. Cells 2021, 10, 3458. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123458
Kashyap S, Rabbani M, de Lima I, Kondrachuk O, Patel R, Shafiei MS, Mukker A, Rajakumar A, Gupta MK. HOPX Plays a Critical Role in Antiretroviral Drugs Induced Epigenetic Modification and Cardiac Hypertrophy. Cells. 2021; 10(12):3458. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123458
Chicago/Turabian StyleKashyap, Shiridhar, Maryam Rabbani, Isabela de Lima, Olena Kondrachuk, Raj Patel, Mahnoush Sophia Shafiei, Avni Mukker, Aishwarya Rajakumar, and Manish Kumar Gupta. 2021. "HOPX Plays a Critical Role in Antiretroviral Drugs Induced Epigenetic Modification and Cardiac Hypertrophy" Cells 10, no. 12: 3458. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123458