
Sperm Selection for ICSI: Do We Have a Winner?
by
 , ,
, ,
Domenico Baldini
1,* ,
,
Daniele Ferri
1,
Giorgio Maria Baldini
1,
Dario Lot
1,
Assunta Catino
2,
Damiano Vizziello
3 and
Giovanni Vizziello
1 1
MOMO Fertilife–IVF Center, 76011 Bisceglie, Italy
2
Department of Biology, University of Bari, 70125 Bari, Italy
3
Department of Urology, Policlinico San Donato Milano, 20100 Milan, Italy
*
Author to whom correspondence should be addressed.
Cells 2021, 10(12), 3566; https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123566
Submission received: 4 November 2021
/
Revised: 14 December 2021
/
Accepted: 15 December 2021
/
Published: 17 December 2021
(This article belongs to the Section Intracellular and Plasma Membranes)
Abstract
:In assisted reproductive technology (ART), the aim of sperm cells’ preparation is to select competent spermatozoa with the highest fertilization potential and in this context, the intracytoplasmic sperm injection (ICSI) represents the most applied technique for fertilization. This makes the process of identifying the perfect spermatozoa extremely important. A number of methods have now been developed to mimic some of the natural selection processes that exist in the female reproductive tract. Although many studies have been conducted to identify the election technique, many doubts and disagreements still remain. In this review, we will discuss all the sperm cell selection techniques currently available for ICSI, starting from the most basic methodologies and continuing with those techniques suitable for sperm cells with reduced motility. Furthermore, different techniques that exploit some sperm membrane characteristics and the most advanced strategy for sperm selection based on microfluidics, will be examined. Finally, a new sperm selection method based on a micro swim-up directly on the ICSI dish will be analyzed. Eventually, advantages and disadvantages of each technique will be debated, trying to draw reasonable conclusions on their efficacy in order to establish the gold standard method.
1. Introduction
Intracytoplasmic sperm injection (ICSI) is the method that has definitely revolutionized the field of ART since normal fertilization and ongoing pregnancies can be achieved even with low quality sperm samples and affected spermatozoa. By injecting a single sperm cell in the oocyte, the technique bypasses several biological barriers that naturally select the gametes to achieve optimal embryonic and fetal development.
ICSI has been proven to show the same efficiencies as in vitro fertilization (IVF), a simpler technique where the oocytes, after pick-up, are left with the prepared sperm sample overnight and fertilization occurs following a sort of natural sperm selection [1]. Nowadays, ICSI is the most used technique infertility treatments in developed countries, replacing IVF as the first choice [2,3].
In fact, since ICSI allows the oocytes to be fertilized by sperm cells that may not be competent, concerns about male inheritance patterns are growing [4].
It has been estimated that about 50% of couples with fertility issues are affected by low sperm cell quality [5] and the reproductive outcome is generally influenced by the paternal genome condition. Some authors [6] suggest that paternal age might impact the quality of the seminal fluid.
This is widely confirmed by many studies based on animal models: when exposed to low doses of chemotherapeutic agents providing an amount of reactive oxygen species (ROS), animals produce sperm cells with epigenetic defects, which lead to deformity in the offspring and are often inherited in the following generations [7]. Human epidemiological studies [8] have shown a correlation between paternal age and correct neurological development. Some studies have shown that the lack of acrosomal reaction and the persistence of the perinuclear theca on the sperm cell head is associated with a delay in de-condensation of the genetic material at the time of fertilization [9]. Other studies have shown that male subjects born from ICSI generate seminal fluid with a low number of cells and with reduced motility when compared to subjects born spontaneously [10]. Further, ICSI has been associated with an increased risk for many health issues, ranging from premature births and diverse metabolic disorders in the offspring to more severe complications, such as abortions, congenital malformations, and imprinting disorders [11]. It has been suggested that one of the reasons for the relatively low efficiency of ART is that we currently lack an effective methodology to separate this specific sperm subpopulation for its use in ARTs [12,13].
In this perspective, effort to identify the eligible technique to select competent sperm cells for ICSI seems mandatory.
2. Natural Sperm Selection
In nature, the process that leads to the selection of the best sperm cells, capable of fertilizing the oocyte in the female genital tract, is very selective and it involves high morphologic qualities and dynamic features. Leaving the seminal plasma in the vagina, sperm cells perform the first step that leads to this process.
Only a small portion of the sperm cells that swim from the uterus to the oviduct can be collected in niches, where cells interact with unknown receptors to form a sperm reserve, as in some animal species the sperm cells can stay in the fallopian tubes even for months before ovulation. However, in the human species, the fertilization window is reduced, and the sperm cells can survive no longer than 5/6 days [14]. This process allows the sperm to reach complete capacitation, which is based on plasma membrane cholesterol ultrastructure changes; it leads to an increase in concentration of intracellular ions, such as Ca2+, that switch the motility patterns of tails to hyper-activation. Moreover, phosphorylation on tyrosine residues determines the preparation for the acrosomal reaction event [15]; subsequent interactions with molecules, such as secreted protein or hormones, from the female reproductive tract, modulate the swim, by a chemotactic and thermotactic effect, towards the oocyte.
Most chemoattractants are still unknown, but progesterone released from cumulus cells certainly plays a key role, allowing the interaction with oocyte receptors and preparing for acrosome reaction [16].
One of the main goals of artificial reproductive technology (ART) is to simulate the strict selection process that takes place in the female tract to improve the reproductive outcome.
Unfortunately, classic methods have given unsatisfactory results, as they rely on separation mechanisms dependent on morphology and motility, such as swim up (SU) and density gradient centrifugation (DGC) [17]. To be adopted in the ART routine procedures, sperm cell preparation techniques should be simple, cheap and fast, allowing a highly efficient selection that differentiates motile and morphologically normal spermatozoa from other cell species, leukocytes or bacteria and toxic substances, avoiding the production of ROS [18].
All current methodologies improve sperm cells quality, but none have been associated with a significant increase in clinical results [17]. Methods for sperm cells’ selection are categorized into classic techniques based on sperm motility or density, and advanced methods that rely on membrane surface charge, high-resolution morphology, and nuclear or membrane integrity. Nowadays, the most used techniques in the ART lab are SU and DGC [18]. Here below the Table 1 represent a summary of all the sperm cell preparation techniques described in the review with advantages and disadvantages of each of them.
3. Classic Methods
3.1. Swim-Up
Lopata and Patullo [19] first described in 1984 the basic principle of recovery motile spermatozoa that manage to migrate through a medium placed on top of the seminal plasma.
The swim-up (Figure 1) is known as the most elementary method for sperm cells’ preparation. The sample is centrifuged with a medium consisting of polysiloxane microparticles to remove the seminal serum and subsequently left for 60 min in an incubator at 37 °C, with a 45° inclination and an overlapped layer of medium. The principle of the technique is based on the attraction of sperm cells to the most enriched area of the liquid. These conditions should promote capacitation and migration: cells with progressive motility can rise from the pellet through the medium and they can be recovered from the surface [64]. There are also several variations of the technique that avoid the centrifugation process where the sperm cells can pass directly from the seminal fluid to the media surface, which is simply placed over the sample [23].
This method is applied both for in vitro fertilization (IVF) and ICSI, and is recommended when motile spermatozoa in the sample are in low percentage [23]. Although there is centrifugation, swim-up is a very soft technique, it produces a small load of ROS, but it recruits only mature activated sperm cells [20]. On the other hand, swim-up has some drawbacks. In the case of samples with high volume or elevated concentration, the method requires a distribution between several tubes of the sample to emphasize the area of contact to the culture medium; this subdivision leads to a burden of workload. Another weak point is the small portion of spermatic cells retrieved; only a maximum of 10% [24] of the total sperm cells population becomes available for further use. If samples are not centrifuged gently, many motile cells from pellets could potentially become anchored to the bottom of the tube, and never reach the culture media layer.
3.2. DGC
In DGC [25] (Figure 1), sperm cells are placed over a continuous or discontinuous density gradient and then centrifuged. Separation occurs as a function of density and motility; the fastest spermatozoa will migrate to the bottom of the tube.
When left on top of the solution with a higher density, the sample is centrifuged for about half an hour. All sperm cells pass through gradients, but motile spermatozoa swim vigorously instead of remaining subjected to the kinetics of centrifugation. Quicker sperm cells form a silky pellet at the bottom of the tube while immotile cells and debris remain between gradients.
The substance used to constitute the gradients should not be toxic for spermatozoa and stable in solution, to avoid any pH alteration or osmolarity.
More specifically, we find hydrophilic organic molecules, such as sucrose or sucrose copolymers (Ficoll) [26], which are less toxic than mixed compounds, such as colloidal silica coated with polyvinylpyrrolidone (Percoll), producing reduced negative effects on the ultra-sperm structure [27]. This method allows the isolation of motile spermatozoa even from a sample with low sperm motility. However, morphology and motility parameters vary considerably among different studies in the literature. The reason for this bias may be caused by different initial conditions, such as concentration, volume, number of layers, g-force applied and centrifugation time. Furthermore, the technique takes longer. DGC is low-yielding when the viscosity of semen is altered, and it produces a higher amount of ROS due to mechanical stress caused by centrifugation. When samples are excessively concentrated, clusters of different types of cells prevent movements through gradients. Unfortunately, density gradient centrifugation is even more expensive than the classic swim-up [24].
Several studies have tried to clarify which one is the ideal method for sperm cells preparation, inspecting morphological and dynamical parameters. Nevertheless, the results available in the literature to date are contradictory.
Since the studies published during the 1990s [65], it has been demonstrated that the SU method selects sperm cells with better motility, vitality and morphology than DGC. Moreover, the latter is less efficient than SU in selecting sperm cells with intact DNA [66]. While these theories have been reinforced by more recent studies [67], in contrast, other authors have shown that DGC is better than SU to select a higher percentage of capacitated sperm [68] with hyper-activated motion even in cases of subfertility [69]. Other scientists suggest that neither method provides a cell population with acceptable levels of intact DNA. Centrifugations related to both techniques might produce an increase of ROS harmful for the spermatic nuclear material [70]. Both methodologies are regularly exploited in the ART lab. Unfortunately, by relying only on motility, these techniques cannot be associated with an optimal quality of spermatozoa [71]. Further studies [21] have shown that by using both methods in combination, better results can be obtained in separating sperm cells with damaged DNA, removing them from the rest of the sample [22]. It has also been observed that spermatozoa with less nuclear fragmentation are correlated with an improvement of the embryo quality and a reduced probability of miscarriage [72]. In performing ICSI, it is crucial to choose sperm cells with optimal characteristics, and using these combined methods, a higher fertilization rate can be achieved even in cases where patients have a normal spermatozoa density but reduced motility.
4. Advanced Methods
4.1. Selection Methods for Sperm Cells with Reduced Motility
When the sperm sample is retrieved by testicular aspiration (TESE) [73] or epididymal aspiration (MESA) [74], the spermatozoa might appear immotile due to the lack of complete maturation that takes place in the final tract of the epididymis [75].
It is important to underline that in these cases, the SU and DGC techniques, that exploit dynamic characteristics for separation, are inadequate.
Marques De Oliveira [76] proposed the technique of mechanical touch (MTT) for the selection of immotile spermatozoa as potentially suitable for ICSI. Briefly, if the tail is still flexible and recovers its original position being touched by an ICSI injection needle, it is considered viable. Instead, rigidity and incapacity to recover the initial tail position is thought as a sign of non-viability [77]. The success and reliability of this technique depends mainly on the expertise of the biologist, and this could be considered as the only current drawback of the latter [76].
An alternative selection technique is the hypo-osmotic swelling test (HOST) [28]. This method (Figure 2) assumes that the tails of viable spermatozoa swell and bend if they are introduced into a hypoosmotic environment, due to the activity of the osmo-sensitive calcium membrane channels [78,79]. HOST can be used to estimate the percentage of integrity in chromatin. Seven distinct patterns of swelling are related to different levels of chromatin integrity. This allows the recognition, through microscopy, of sperm cells with better nuclear material for ICSI treatments [29,30]. According to the World Health Organization laboratory manual for the examination and processing of human semen [23], HOST is advised in cases of asthenozoospermia to collect suitable spermatozoa for ICSI. Some studies suggested that HOST should be employed in a subsequent ICSI cycle following total fertilization failure [80].
Some controversial studies instead suggest that the use of the HOST is associated with a low fertilization rate, and non-viability of sperm after 30 min of incubation in hypo-osmotic solution [31]. Moreover, biologists must work with extremely limited quantities of testicular sperm in many cases, and the HOST test involves dilution of the sample.
Hence, this method should be prerogative depending on the specific ART laboratory settings and on the amount of sperm sample available [32].
Using polarized light implemented in the microscope (Figure 3) is another possible strategy for the selection of immotile sperm cells [33]. Some research teams have proposed its use to verify the birefringence of sperm cell head as an index of suitability. As a matter of fact, the sub-acrosomal protein filaments extend themselves longitudinally, giving a typical pattern of birefringence to the sperm cell head [34]. In some published studies, the selection of non-motile spermatozoa with birefringent heads resulted in an increase in clinical pregnancy and implantation rate (58% vs. 9% and 42% vs. 12%), compared to a control group with immotile spermatozoa (where polarized light was not used) [81], and an increased implantation rate when the HOST technique was used instead as selection method (45% vs. 11%) [82].
Unfortunately, even this technique presents some disadvantages: the polarized light implemented in the microscope is expensive and there is a lack of data regarding the integrity of the sperm DNA. Furthermore, the success of the latter is based largely on the experience of the operator.
In this perspective, it is fundamental to underline that sperm DNA integrity is essential for fertilization and early stages of embryo development [83]. It is known that sperm cells are not able to repair the DNA after spermiogenesis since they do not express a DNA repair mechanism. [84,85]. Due to the sperm physiology and the environment that they are exposed to, DNA damage is inevitable, but post-fertilization it can be repaired at least partially. When the damage exceeds the oocyte’s repair capacity, it might result in absence of fertilization [86]. Studies have shown that the chance of both natural and artificial conceptions is reduced if the DNA fragmentation index is more than 30% [84,87,88]. Hence, a wide range of techniques have been developed in order to assess the sperm chromatin and DNA integrity, despite standardization or guidelines to uniformize results remain absent [89].
A group of researchers has shown another potential method to recover immotile but suitable sperms for ICSI. It exploits the chemical inducers of motility (Figure 4), that belong to the class of phosphodiesterase inhibitors, such as pentoxifylline (PTF), dimethylxanthines and papaverines [90], allowing the motility reactivation of spermatozoa recovered from the testicular and epididymal level [91]. Using this motile fraction, it was possible to obtain fertilization, normal pregnancies, and an increase in birth rate through ICSI [92,93,94]. Some doubts about the technique concern the intrinsic toxicity of these molecules towards the genetic material, but results from studies suggest that in vitro treatment adequately improves motility of immotile sperm, without leading to acrosome reaction, DNA damage, and viability loss [95,96]. Comparing this technique with HOST, researchers have obtained a higher fertilization and pregnancy rate (32% vs. 16%) [97].
Alternatively, motility can be further reactivated by using a laser incorporated in the magnification system that is shot at the immotile sperm cell tail (LAISS laser-assisted immotile sperm selection) (Figure 5) [35]. Laser irradiation causes the release in cytosol of second messengers, such as Ca2+ or ROS, and an increase in the synthesis of ATP, which could generate a slight movement of the tail [98]. The sperm cell is then considered viable when its tail is coiled after the laser shot [36]. On the other hand, some authors claim that high laser doses cause an excess of potentially toxic ROS [39]. Furthermore, more Ca2+ influx induces the hyperactivity of Ca2+-ATPase calcium channels and exhausts the ATP reserves of the cell. This process could lead to depletion of cell channels activity, correlated with an increase in internal osmotic pressure causing the swelling of the sperm cell and subsequently the rupture of its plasma membrane [40]. Contrary, other authors suggest that this technique does not damage the spermatic membrane and does not affect the percentage of fragmentation of the genetic material [99]. LAISS is also an alternative to the chemicals’ motility activators, which could be potentially toxic [100]. In addition, it significantly increases the embryonic segmentation and post ICSI birth rate, using testicular or ejaculated spermatozoa, when compared to control groups [101]. LAISS is the recommended technique in case patients suffer from primary ciliary dyskinesia [37] or Kartagener’s syndrome [38]. Despite the high potential of the method, the complexity and cost are the main reasons why it is not adopted in routine practice in ART laboratories.
It has also been demonstrated that the use of ATP/MgSO4 generates an increase in motility in testicular seminal samples, generating a weak contraction of the flagellum (Figure 6). The results of the study have shown that using ATP as a solute, the induced motility increases significantly, and viable sperm cells can be captured and used for injection through ICSI [102].
Alternatively, the myoinositol can be used to isolate viable sperm cells for ICSI. It represents the most abundant stereoisomer of the inositol class modulating the intracellular concentration of Ca2+ [103]. This is synthesized in two steps by two enzymes, myo-1-phosphate synthase and myo-monophosphatase-1, located in high concentration in the testicular mesenchymal tissue [104,105]. By incubating sperm cells frozen and then thawed from oligoasthenospermic patients with myoinositol, some research groups were able to recover a portion of sperm cells with significantly increased motility [101]. Myoinositol generates an increase in mitochondrial membrane potential, resulting in an intracellular increase of Ca2+ [106]. The research group performed a series of ICSI by pre-incubating sperm cells from oligoasthenospermic patients in solutions containing myo-inositol [107]. The increase in motility and fertilization rate has generated encouraging results. Unfortunately, one of the key objectives for ART laboratories is to mimic the culture conditions in vivo and myoinositol, as the ATP/MgSO4, could generate an uncontrolled increase in the mitochondrial membrane potential which could lead to the occurrence of toxic effects. Further studies should be conducted to incorporate this practice into clinical routines [108].
4.2. Sperm Cells Selection by Membrane Characteristics
The outer sperm membrane is critical for their functionality since it is firstly involved in many aspects of the fertilization such as capacitation, oocyte binding and acrosome reaction [109,110]. In order to select high-quality sperm cells, methods that gain benefits from the characteristics of their membrane have been studied.
The zona pellucida binding assay (ZBA) is a technique that allows the selection of mature and competent spermatozoa. The zona pellucida has specific receptors for the latter [111]. An immature oocyte, recovered from the cohort obtained from a patient’s oocyte retrieval, is incubated with the sperm cells of interest, previously selected with DGC. Sperm cells that bind the zona pellucida are retrieved through a microinjection needle, subsequently used to perform the ICSI. From some studies, it emerges that sperm selection with this method leads to improved embryo quality and an increase in implantation rates [112]. Although there is no routine application in the IVF laboratories, probably due to the heavy workload, the technique is recommended for cases of repeated fertilization failure [113].
A further method that takes advantage of the membrane characteristics is the magnetic activated cell sorting (MACS). This methodology allows the selection of the non-apoptotic portion from a sample of interest [114]. It involves the use of magnetic microspheres conjugated to Annexin V (AV-MACS) (Figure 7) [115], that have a high affinity for phosphatidyl-serine. The latter is normally exposed on the outer side of the membrane when the sperm cells are in an apoptotic state [116]. The seminal sample of interest passes through a column containing microspheres, to which the annexin has adhered. The non-viable spermatozoa remain trapped inside the column, while the viable fraction is eluted, improving the vitality characteristics of the starting sample [41,42].
Several studies have shown that the MACS method is efficient in cases of high nuclear fragmentation, idiopathic infertility, and patients with varicocele [43,44]. When this technique was performed in combination with classical procedures, such as swim-up or density gradient centrifugation, the recovered spermatozoa exhibited an even lower percentage of fragmentation [45,117]. A study conducted on a group of oligoasthenzoospermic patients reported that the seminal sample subjected to MACS led to an increase in embryonic segmentation and pregnancy rate when compared to simple centrifugation on a density gradient [118]. Unfortunately, also in this case, the literature remains incomplete on the percentage of live births, and the method does not allow discrimination between the type of motile spermatozoa selected.
Hyaluronic acid is the main component of the extracellular matrix surrounding the cumulus-oocyte complex [119]. Only the sperm cells that have successfully completed spermatogenesis and maturation are able to show the polysaccharide-binding receptors on their outer membrane [46,47]. Moreover, they are usually characterized by a normal morphology and a low percentage of fragmentation of the nuclear material [48]. Exploiting these characteristics, a methodology for sperm cell selection has been devised where spermatozoa are incubated in plates or in media containing hyaluronic acid; only the sperm cells able to bind the molecule are then used to perform a modified version of ICSI: physiological intracytoplasmic sperm injection (PICSI) [120,121]. The selection should result in an increased fertilization rate, but in this case, the clinical data are conflicting. Some studies suggest that both the fertilization rate and the percentage of top-quality embryos benefit from the technique (92 vs. 86; 36 vs. 24%) [122], but other research groups do not confirm these improvements [49]. Studies show that a negative charge is present on the sperm cell membrane [123]. The Zeta method (Figure 8) uses this feature to separate sperm cells containing the Y chromosome from those containing the X chromosome [50]. Two distinct research groups have developed two different methodologies, one using a positively charged centrifuge tube [54], the other using migration in an electrophoretic field [55], which allow the collection of live spermatozoa with normal morphology and a high percentage of the integrity of the genetic material [51,52,124,125]. To date, only a randomized study has been conducted on spermatozoa selected with the Zeta method and subjected to ICSI [53]. This showed a significant increase in the percentage of top-quality embryos (45.83 ± 3.11% vs. 35.38 ± 4.64% (p = 0.04)) and in the pregnancy rate (39.2 vs. 21.8 (p = 0.009) as compared to the method of density gradient centrifugation.
4.3. Selection Based on Morphology–IMSI
The analysis of semen quality has for a long time been associated with the morphological evaluation of spermatozoa. The introduction of digital microscopy has made its possible to analyze the ultrastructural characteristics of motile spermatozoa (MSOME; motile sperm organelle morphology examination) [126]. The integration of the MSOME into the ICSI method allows a high-magnificence micro-injection (IMSI) (Figure 9) [127]. The selection of motile spermatozoa with few vacuoles and normal nuclear morphology is possible through a 6000× magnification system integrated with the micromanipulation system [128]. Again, the scientific data are promising but conflicting. Using this method, some studies [56] have revealed that the presence of large vacuoles in the sperm nucleus is associated with a high level of nuclear fragmentation. In case of repeated fertilization failure with the ICSI method, the IMSI method has been suggested as preferential [57]. On the other hand, some authors [58] did not find significant differences in terms of fragmentation or pregnancy rates between ICSI and IMSI. Recently, in a study conducted on a large sample of semen analyzed with MSOME, no correlation was found between the presence of vacuoles in the nucleus and DNA fragmentation [59]. The authors argue that vacuoles are physiologically present in the sperm head, and they do not affect their functions. Unfortunately, due to lack of literature, the timing of the analysis and the cost of the equipment, the IMSI method has not been fully adopted in the routine practices of ART clinics.
4.4. Dynamic Selection of Spermatozoa
The three main mechanisms that control the movements of spermatozoa within the oviducts, rheotaxis, chemotaxis and thermotaxis, have been exploited as methods for sperm cell selection, to improve the ART outcomes:
- Human spermatozoa orient their motion, by rheotaxis, against the fluid stream directed towards them [129]. Rheotaxis is defined as the tendency of certain living beings to move in response to the mechanical stimulus of a current of water. Some studies have confirmed the presence of a flow-directed towards the uterus in the oviductal region, capable of increasing its intensity after intercourse, which is able to attract the sperm cells [130]. In the technique developed by Nagata et al. [131], using bull sperm as a mammalian model, a flow is created through a series of microchannels towards a well where the sperm sample is delivered. In response to the flow, the spermatozoa swim towards it passing through the microchannels and finally reaching a receptive well where they can be collected for ART applications. Another study was conducted on normospermic patients [132] to select spermatozoa using rheotaxis: compared to an untreated sample and a sample subjected to density gradient centrifugation, the recovered spermatozoa showed higher chromatin compactness (99% vs. 71% vs. 83%). In recent years, attempts have been made to include the concept of rheotaxis in microfluidics [133], but despite the promising data, an analysis on the quality of selected spermatozoa has not been conducted and the passive nature of rheotaxis correlates to the inability to discern the different types of motility.
- Progesterone plays the main role as a chemo-attractor for the navigation of spermatozoa through the environment surrounding the cumulus-oocyte complex [134]. Only capacitated spermatozoa possess the receptors to recognize and bind this type of molecule [134]. Some sperm cell selection techniques exploit this feature to distinguish capacitated spermatozoa from non-capacitated ones. Several studies have been conducted on different types of sperm populations, using a device where a progesterone concentration gradient is constituted (sperm selection assay) [135]. In this method, two wells are connected by a 2 mm length per 2.5 mm diameter tube. One of the wells is filled with a media containing the chemoattractant molecule in solution (progesterone for instance) and the other well is filled with the human sperm sample. The chemoattractant diffuses through the tube generating a gradient and the spermatozoa respond by moving towards the higher concentration and accumulating in the initial well free of cells where they can be used for ART applications. The recovered spermatozoa from this technique exhibit better morphology, less DNA fragmentation and a reduced rate of apoptosis as compared to those selected with density gradient centrifugation [136]. Although the results are also promising in this case, further studies need to be conducted to confirm the improvements in the clinical field.
- The spermatozoa can direct their motion according to the variation in temperature, moving from cold areas to warmer areas [137]. Several studies show that this mechanism underlies the movement of sperm cells from the fallopian tubes to the ampulla [138,139]. Even in this case, however, only the capacitated spermatozoa can respond to the temperature gradient, making this motion a mechanism for the selection of sperm cells with better fertilizing characteristics. The method of sperm selection by thermotaxis was developed by Pérez-Cerezales et al. [140]. Spermatozoa are placed in a drop of medium, connected by a capillary to a second drop free of cells. A temperature gradient is generated between both drops, as the highest temperature is maintained in the drop free of cells. Spermatozoa respond by thermotaxis migrating towards the warmer temperature and accumulating in the second drop where they can be collected for downstream applications.
The purpose was to select spermatozoa with high nuclear integrity, compared to those isolated by swim-up. Unfortunately, even in this case, the literature does not offer a variety of studies that fully confirm the improvements for ART efficacy in the human clinic.
4.5. Microfluidics Applied to Sperm Selection
Technologies related to microfluidics are rapidly growing within ART laboratories. Among the first experiments with this methodology, Smith and Takayama [60] have published a series of articles demonstrating the efficiency of the method for the selection of high-quality spermatozoa.
By controlling fluid dynamics (Figure 10), within millimeter diameter capillaries, it is possible to mimic the physiological conditions of pH and temperature of the female genital tract [141]. Hence, we could potentially select spermatozoa with increased motility through flows [142], chemical gradients [143] or electrophoretic fields [144]. The method of Smith and Takayama [60] uses two parallel laminar flow channels. While the motile spermatozoa can move through the flows and be eluted separately, the debris and immotile cells are passively transported from the entrance to the exit of the capillary canal. Compared to a classic density gradient centrifugation, the morphology and motility of the selected spermatozoa are significantly increased (98% and 22%). Parrella et al. [61] report in a study performed on a small pool of couples undergoing ICSI, that spermatozoa selected using these devices could be linked to an increase in pregnancy rate (71%).
In the near future, microfluidics based on morpho-dynamic characteristics will be replaced by further innovations, including integrating chemo-attracting selection systems [145,146], cumulus cells in the selection chamber or even layers of oviductal cells [147].
The application of optical systems to microfluidics could further improve the selection system [148]. Raman spectroscopy allows the discernment of sperm cells with high nuclear integrity from sperm cells with fragmented DNA [149]. By coupling the three-dimensional imaging to the selected channels, it is possible to accurately identify the different types of flagellar movement using a digital sensor, consisting of a semi-conductive material, which can virtually reconstruct the cell volume by recording the differences in the types of interference between a reference light wave and a scatter from the sample [150]. Using this method, De Wagenaar B. et al., were able to trace the profile of the flagellar beat of hyper-activated spermatozoa [151].
Among the advantages that microfluidics certainly offer are automation, scalability and reduction in diagnosis and preparation times. Unfortunately, the high cost of instrumentation and poor yield of the sample in terms of volume represent the main limitations of the method.
4.6. Horizontal Sperm Migration
The method developed by our research group [62] called “horizontal sperm migration” involves the preparation of a modified ICSI plate, including three additional drops of a solution containing HEPES (G-mops® Vitrolife, Göteborg, Sweden). The drops are connected through a medium bridge shaped by swiping the denuding pipette across them. In relation to concentration and motility, observed from an analysis previously performed under the microscope, 1 to 5 µL of ejaculate are loaded into the proximal portion of the medium strip, about ten minutes before performing the oocyte injection. During ICSI, an adequate number of spermatozoa reaches the distal edge of the furthest drop; some of these can be recovered from the injection needle, to be moved to the PVP and carefully selected for fertilization (Figure 11).
The technique allows the recovery of spermatozoa with high motility, normal morphology and minimal damage to the DNA, using a fast, safe, and economical procedure. Comparing the clinical results obtained from this new selection method with the classic swim-up technique, no significant differences were found in terms of fertilization and implantation rate. On the other hand, segmentation and blastocyst rates are higher in the horizontal swim-up, suggesting that this process that generates a lower quantity of harmful oxygen reactants is correlated with better seminal quality and a lower DNA fragmentation. In line with our observations, clinical and ongoing pregnancy rates are numerically better in the horizontal swim-up than in conventional methods although there is no significant statistical difference. By comparing a variant of horizontal migration with density gradient centrifugation, a previous study [63] found out that fertilization rates between the two methodologies are similar, but blastocyst and pregnancy rates results improved in favor of the innovative technology.
In terms of timing, while for the swim-up and the centrifugation on the density gradient it takes between 50 and 80 min for the preparation of samples, with this innovative method it is possible to perform the oocyte injection 10 minutes after placing the migrating semen sample. Furthermore, the procedure is much less expensive since there is no necessity to use culture media required for preparation in other methods. Moreover, by avoiding the use of several tubes of media, the absence of bacterial contamination is also ensured. Furthermore, it is also possible to improve the recognition system and avoid mismatch errors in patient identification.
5. Conclusions
Each method has shown advantages and disadvantages, but none seem to be superior at this time (Table 1). Despite the countless number of methods, none have shown outstanding quality or results in increasing pregnancy rates. To date, there are many new methods that present encouraging results, but we still do not have a randomized controlled trial with sufficient data to demonstrate that one of these could be eligible as a gold standard technique. Considering this, all the methods listed in this paper offer applicability in different contexts with similar clinical results. More specifically, our methodology meets the demands of an ART clinic, where reduced timing and work quality must coexist. It is fast, cheap, easy to learn, highly repeatable, and results in pregnancy rates are comparable to the most innovative methods.
Author Contributions
We declare that all the authors have made substantial contributions to all the aspects of the manuscript. We summarized the specific contributions of each author just below: D.B. conception and design of the review, final approval of the version to be submitted; D.V. drafting the article or revising it critically for important intellectual content; G.V. contributed to the design and implementation of the review; D.F. drafting the article and revising it critically for important intellectual content; final approval of the version to be submitted; A.C. references research and revising for intellectual content; D.L. references research and revising for intellectual content; G.M.B. drafted the manuscript and designed the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schwarze, J.E.; Jeria, R.; Crosby, J.; Villa, S.; Ortega, C.; Pommer, R. Is there a reason to perform ICSI in the absence of male factor? Lessons from the Latin American Registry of ART. Hum. Reprod. Open 2017, 2017, hox013. [Google Scholar] [CrossRef] [Green Version]
- CDC. 2016—National Summary Report—Assisted Reproductive Technology; US Department of Health. Human Service: Washington, DC, USA, 2018. [Google Scholar]
- De Geyter, C.; Calhaz-Jorge, C.; Kupka, M.S.; Wyns, C.; Mocanu, E.; Motrenko, T.; Scaravelli, G.; Smeenk, J.; Vidakovic, S.; Goossens, V. European IVF-monitoring Consortium (EIM) for the European Society of Human Reproduction and Embryology (ESHRE). ART in Europe, 2014: Results generated from European registries by ESHRE: The European IVF-monitoring Consortium (EIM) for the European Society of Human Reproduction and Embryology (ESHRE). Hum. Reprod. 2018, 33, 1586–1601. [Google Scholar] [CrossRef]
- Henningsen, A.A.; Gissler, M.; Rasmussen, S.; Opdahl, S.; Wennerholm, U.B.; Spangsmose, A.L.; Tiitinen, A.; Bergh, C.; Romundstad, L.B.; Laivuori, H.; et al. Imprinting disorders in children born after ART: A nordic study from the CoNARTaS group. Hum. Reprod. 2020, 35, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Levine, H.; Jorgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef]
- Nilsson, E.E.; Sadler-Riggleman, I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenet. 2018, 4, dvy016. [Google Scholar] [CrossRef] [Green Version]
- Janecka, M.; Mill, J.; Basson, M.A.; Goriely, A.; Spiers, H.; Reichenberg, A.; Schalkwyk, L.; Fernandes, C. Advanced paternal age effects in neurodevelopmental disorders—review of potential underlying mechanisms. Transl. Psychiatry 2017, 7, e1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Onofrio, B.M.; Rickert, M.E.; Frans, E.; Kuja-Halkola, R.; Almqvist, C.; Sjolander, A.; Larsson, H.; Lichtenstein, P. Paternal age at childbearing and offspring psychiatric and academic morbidity. JAMA Psychiatry 2014, 71, 432–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belva, F.; Bonduelle, M.; Roelants, M.; Michielsen, D.; Van Steirteghem, A.; Verheyen, G.; Tournaye, H. Semen quality of young adult ICSI offspring: The first results. Hum. Reprod. 2016, 31, 2811–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Calabuig, M.J.; López-Cardona, A.P.; Fernández-González, R.; Ramos-Ibeas, P.; Fonseca Balvís, N.; Laguna-Barraza, R.; Pericuesta, E.; Gutiérrez-Adán, A.; Bermejo-Álvarez, P. Potential Health Risks Associated to ICSI: Insights from Animal Models and Strategies for a Safe Procedure. Front. Public Health 2014, 2, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakkas, D.; Ramalingam, M.; Garrido, N.; Barratt, C.L. Sperm selection in natural conception: What can we learn from Mother Nature to improve assisted reproduction outcomes? Hum. Reprod. Update 2015, 21, 711–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Cerezales, S.; Ramos-Ibeas, P.; Acuña, O.S.; Avilés, M.; Coy, P.; Rizos, D.; Gutiérrez-Adán, A. The oviduct: From sperm selection to the epigenetic landscape of the embryo. Biol. Reprod. 2018, 98, 262–276. [Google Scholar] [CrossRef] [Green Version]
- Suarez, S.S.; Pacey, A.A. Sperm transport in the female reproductive tract. Hum. Reprod. Update 2006, 12, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Puga Molina, L.C.; Luque, G.M.; Balestrini, P.A.; Marín-Briggiler, C.I.; Romarowski, A.; Buffone, M.G. Molecular Basis of Human Sperm Capacitation. Front. Cell Dev. Biol. 2018, 6, 72. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Silva, G.; Chirinos, M. Proteins from male and female reproductive tracts involved in sperm function regulation. Zygote 2019, 27, 5–16. [Google Scholar] [CrossRef]
- ESHRE Guideline Group on Good Practice in IVF Labs; De los Santos, M.J.; Apter, S.; Coticchio, G.; Debrock, S.; Lundin, K.; Vermeulen, N. Revised guidelines for good practice in IVF laboratories. Hum. Reprod. 2015, 31, 685–686. [Google Scholar]
- Vaughan, D.A.; Sakkas, D. Sperm selection methods in the 21st century. Biol. Reprod. 2019, 101, 1076–1082. [Google Scholar] [CrossRef]
- Lopata, A.; Patullo, M.J.; Chang, A.; James, B. A method collecting motile spermatozoa from human semen. Fertil. Steril. 1976, 27, 677. [Google Scholar] [CrossRef]
- Bui, A.D.; Sharma, R.; Henkel, R.; Agarwal, A. Reactive oxygen species impact on sperm DNA and its role in male infertility. Andrologia 2018, 50, e13012. [Google Scholar] [CrossRef]
- Ghumman, S.; Adiga, S.K.; Upadhya, D.; Kalthur, G.; Jayaraman, V.; Rao, S.B.; Kumar, P. Combination of swim-up and density gradient separation methods effectively eliminate DNA damaged sperm. J. Turk. Ger. Gynecol. Assoc. 2011, 12, 148–152. [Google Scholar] [CrossRef]
- Jayaraman, V.; Upadhya, D.; Narayan, P.K.; Adiga, S.K. Sperm processing by swim-up and density gradient is effective in elimination of sperm with DNA damage. J. Assist. Reprod. Genet. 2012, 29, 557–563. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Henkel, R.R.; Schill, W.B. Sperm preparation for ART. Reprod. Biol. Endocrinol. 2003, 1, 108. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.A. A highly efficient method for washing mammalian spermatozoa. J. Reprod. Fertil. 1976, 48, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Jee, B.C. Evaluation of normal morphology, DNA fragmentation, and hyaluronic acid binding ability of human spermatozoa after using four different commercial media for density gradient centrifugation. Clin. Exp. Reprod. Med. 2019, 46, 8–13. [Google Scholar] [CrossRef]
- Strehler, E.; Baccetti, B.; Sterzik, K.; Capitani, S.; Collodel, G.; De Santo, M.; Gambera, L.; Piomboni, P. Detrimental effects of polyvinylpyrrolidone on the ultrastructure of spermatozoa (Notulae seminologicae 13). Hum. Reprod. 1998, 13, 120–123. [Google Scholar] [CrossRef] [Green Version]
- Jeyendran, R.S.; Van der Ven, H.H.; Perez-Pelaez, M.; Crabo, B.G.; Zaneveld, L.J. Development of an assay to assess the functional integrity of the human sperm membrane and its relationship to other semen characteristics. J. Reprod. Fertil. 1984, 70, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Bassiri, F.; Tavalaee, M.; Shiravi, A.H.; Mansouri, S.; Nasr-Esfahani, M.H. Is there an association between HOST grades and sperm quality? Hum Reprod. 2012, 27, 2277–2284. [Google Scholar] [CrossRef] [Green Version]
- Bloch, A.; Rogers, E.J.; Nicolas, C.; Martin-Denavit, T.; Monteiro, M.; Thomas, D.; Morel, H.; Lévy, R.; Siffroi, J.P.; Dupont, C.; et al. Detailed cell-level analysis of sperm nuclear quality among the different hypo-osmotic swelling test (HOST) classes. J. Assist. Reprod. Genet. 2021, 38, 2491–2499. [Google Scholar] [CrossRef]
- Casper, R.F.; Meriano, J.S.; Jarvi, K.A.; Cowan, L.; Lucato, M.L. The hypo-osmotic swelling test for selection of viable sperm for intracytoplasmic injection in men with complete asthenozoospermia. Fertil. Steril. 1996, 65, 972–976. [Google Scholar] [CrossRef]
- Tsai, Y.L.; Liu, J.; Garcia, J.E.; Katz, E.; Compton, G.; Baramki, T.A. Establishment of optimal hypo-osmotic swelling test by examining single spermatozoa from different hypo-osmotic solutions. Hum. Reprod. 1997, 12, 11–113. [Google Scholar] [CrossRef] [Green Version]
- Vermey, B.G.; Chapman, M.G.; Cooke, S.; Kilani, S. The relationship between sperm head retardance using polarized light microscopy and clinical outcomes. Reprod. Biomed. Online 2015, 30, 67–73. [Google Scholar] [CrossRef]
- Baccetti, B. Microscopical advances in assisted reproduction. J. Submicrosc. Cytol. Pathol. 2004, 36, 333–339. [Google Scholar]
- Aktan, T.M.; Montag, M.; Duman, S.; Gorkemli, H.; Rink, K.; Yurdakul, T. Use of a laser to detect viable but immotile spermatozoa. Andrologia 2004, 36, 366–369. [Google Scholar] [CrossRef]
- Birowo, P.; Tendi, W.; Rasyid, N.; Turek, P.J.; Sini, I.R.; Rizal, M. Successful Targeted Testicular Sperm Extraction Using Microsurgical Technique (microTESE) Following Fine Needle Aspiration (FNA) Mapping in a Non-Obstructive Azoospermia (NOA) Patient: A Case Report. J. Reprod. Infertil. 2021, 22, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.A.; Kruse, R.; Hirchenhain, J.; Krüssel, J.S.; Neumann, N.J. Pregnancy after laser-assisted selection of viable spermatozoa before intracytoplasmatic sperm injection in a couple with male primary cilia dyskinesia. Fertil. Steril. 2008, 89, e9–e12. [Google Scholar] [CrossRef]
- Ozkavukcu, S.; Celik-Ozenci, C.; Konuk, E.; Atabekoglu, C. Live birth after Laser Assisted Viability Assessment (LAVA) to detect pentoxifylline resistant ejaculated immotile spermatozoa during ICSI in a couple with male Kartagener’s syndrome. Reprod. Biol. Endocrinol. 2018, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Salman Yazdi, R.; Bakhshi, S.; Jannat Alipoor, F.; Akhoond, M.R.; Borhani, S.; Farrahi, F.; Lotfi Panah, M.; Sadighi Gilani, M.A. Effect of 830-nm diode laser irradiation on human sperm motility. Lasers Med. Sci. 2014, 29, 97–104. [Google Scholar] [CrossRef]
- Montag, M.; Rink, K.; Delacrétaz, G.; van der Ven, H. Laser-induced immobilization and plasma membrane permeabilization in human spermatozoa. Hum. Reprod. 2000, 15, 846–852. [Google Scholar] [CrossRef] [Green Version]
- Hoogendijk, C.F.; Kruger, T.F.; Bouic, P.J.; Henkel, R.R. A novel approach for the selection of human sperm using annexin V-binding and flow cytometry. Fertil. Steril. 2009, 91, 1285–1292. [Google Scholar] [CrossRef]
- Grunewald, S.; Paasch, U.; Glander, H.J. Enrichment of non-apoptotic human spermatozoa after cryopreservation by immunomagnetic cell sorting. Cell Tissue Bank. 2001, 2, 127–133. [Google Scholar] [CrossRef]
- Lee, T.H.; Liu, C.H.; Shih, Y.T.; Tsao, H.M.; Huang, C.C.; Chen, H.H.; Lee, M.S. Magnetic-activated cell sorting for sperm preparation reduces spermatozoa with apoptotic markers and improves the acrosome reaction in couples with unexplained infertility. Hum. Reprod. 2010, 25, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Degheidy, T.; Abdelfattah, H.; Seif, A.; Albuz, F.K.; Gazi, S.; Abbas, S. Magnetic activated cell sorting: An effective method for reduction of sperm DNA fragmentation in varicocele men prior to assisted reproductive techniques. Andrologia 2015, 47, 892–896. [Google Scholar] [CrossRef]
- Nadalini, M.; Tarozzi, N.; Di Santo, M.; Borini, A. Annexin V magnetic-activated cell sorting versus swim-up for the selection of human sperm in ART: Is the new approach better than the traditional one? J. Assist. Reprod. Genet. 2014, 31, 1045–1051. [Google Scholar] [CrossRef] [Green Version]
- Cayli, S.; Jakab, A.; Ovari, L.; Delpiano, E.; Celik-Ozenci, C.; Sakkas, D.; Ward, D.; Huszar, G. Biochemical markers of sperm function: Male fertility and sperm selection for ICSI. Reprod. Biomed. Online. 2003, 7, 462–468. [Google Scholar] [CrossRef]
- Huszar, G.; Ozkavukcu, S.; Jakab, A.; Celik-Ozenci, C.; Sati, G.L.; Cayli, S. Hyaluronic acid binding ability of human sperm reflects cellular maturity and fertilizing potential: Selection of sperm for intracytoplasmic sperm injection. Curr. Opin. Obstet. Gynecol. 2006, 18, 260–267. [Google Scholar] [CrossRef]
- Jakab, A.; Sakkas, D.; Delpiano, E.; Cayli, S.; Kovanci, E.; Ward, D.; Revelli, A.; Huszar, G. Intracytoplasmic sperm injection: A novel selection method for sperm with normal frequency of chromosomal aneuploidies. Fertil. Steril. 2005, 84, 1665–1673. [Google Scholar] [CrossRef]
- Van Den Bergh, M.J.; Fahy-Deshe, M.; Hohl, M.K. Pronuclear zygote score following intracytoplasmic injection of hyaluronan-bound spermatozoa: A prospective randomized study. Reprod. Biomed. Online 2009, 19, 796–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelmann, U.; Krassnigg, F.; Schatz, H.; Schill, W.B. Separation of human X and Y spermatozoa by free-flow electrophoresis. Gamete Res. 1988, 19, 151–160. [Google Scholar] [CrossRef]
- Kheirollahi-Kouhestani, M.; Razavi, S.; Tavalaee, M.; Deemeh, M.R.; Mardani, M.; Moshtaghian, J.; Nasr-Esfahani, M.H. Selection of sperm based on combined density gradient and Zeta method may improve ICSI outcome. Hum. Reprod. 2009, 24, 2409–2416. [Google Scholar] [CrossRef] [Green Version]
- Razavi, S.H.; Nasr-Esfahani, M.H.; Deemeh, M.R.; Shayesteh, M.; Tavalaee, M. Evaluation of zeta and HA-binding methods for selection of spermatozoa with normal morphology, protamine content and DNA integrity. Andrologia 2010, 42, 13–19. [Google Scholar] [CrossRef]
- Nasr Esfahani, M.H.; Deemeh, M.R.; Tavalaee, M.; Sekhavati, M.H.; Gourabi, H. Zeta Sperm Selection Improves Pregnancy Rate and Alters Sex Ratio in Male Factor Infertility Patients: A Double-Blind, Randomized Clinical Trial. Int. J. Fertil. Steril. 2016, 10, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.J.; Jacobson, J.D.; Corselli, J.U.; Patton, W.C. A simple zeta method for sperm selection based on membrane charge. Fertil. Steril. 2006, 85, 481–486. [Google Scholar] [CrossRef]
- Ainsworth, C.; Nixon, B.; Aitken, R.J. Development of a novel electrophoretic system for the isolation of human spermatozoa. Hum. Reprod. 2005, 20, 2261–2270. [Google Scholar] [CrossRef] [Green Version]
- Shalom-Paz, E.; Anabusi, S.; Michaeli, M.; Karchovsky-Shoshan, E.; Rothfarb, N.; Shavit, T.; Ellenbogen, A. Can intra cytoplasmatic morphologically selected sperm injection (IMSI) technique improve outcome in patients with repeated IVF-ICSI failure? a comparative study. Gynecol. Endocrinol. 2015, 31, 247–251. [Google Scholar] [CrossRef]
- Leandri, R.D.; Gachet, A.; Pfeffer, J.; Celebi, C.; Rives, N.; Carre-Pigeon, F.; Kulski, O.; Mitchell, V.; Parinaud, J. Is intracytoplasmic morphologically selected sperm injection (IMSI) beneficial in the first ART cycle? a multicentric randomized controlled trial. Andrology 2013, 1, 692–697. [Google Scholar] [CrossRef]
- Teixeira, D.M.; Hadyme Miyague, A.; Barbosa, M.A.; Navarro, P.A.; Raine-Fenning, N.; Nastri, C.O.; Martins, W.P. Regular (ICSI) versus ultra-high magnification (IMSI) sperm selection for assisted reproduction. Cochrane Database Syst. Rev. 2020, 2, CD010167. [Google Scholar] [CrossRef]
- Fortunato, A.; Boni, R.; Leo, R.; Nacchia, G.; Liguori, F.; Casale, S.; Bonassisa, P.; Tosti, E. Vacuoles in sperm head are not associated with head morphology, DNA damage and reproductive success. Reprod. Biomed. Online 2016, 32, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.D.; Takayama, S. Application of microfluidic technologies to human assisted reproduction. Mol. Hum. Reprod. 2017, 23, 257–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrella, A.C.D.; Keating, D.; Rosenwaks, Z.; Palermo, G.D. A microfluidic devide for selecting the most progressively motile spermatozoa yields a higher rate of euploid embryos. Fertil. Steril. 2018, 110, e342. [Google Scholar] [CrossRef] [Green Version]
- Baldini, D.; Baldini, A.; Silvestris, E.; Vizziello, G.; Ferri, D.; Vizziello, D. A fast and safe technique for sperm preparation in ICSI treatments within a randomized controlled trial (RCT). Reprod. Biol. Endocrinol. 2020, 18, 88. [Google Scholar] [CrossRef] [PubMed]
- Palini, S.; Stefani, S.; Primiterra, M.; Benedetti, S.; Barone, S.; Carli, L.; Vaccari, E.; Murat, U.; Feichtinger, W. Comparison of in vitro fertilization outcomes in ICSI cycles after human sperm preparation by density gradient centrifugation and direct micro swim-up without centrifugation. JBRA Assist. Reprod. 2017, 21, 89–93. [Google Scholar] [CrossRef]
- Mahadevan, M.; Baker, G. Assessment and Preparation of Semen for in Vitro Fertilization Clinical in Vitro Fertilization, and A. Trounson; Springer: London, UK, 1984; pp. 83–97. [Google Scholar]
- Sanchez, R.; Stalf, T.; Khanaga, O.; Turley, H.; Gips, H.; Schill, W.B. Sperm selection methods for intracytoplasmic sperm injection (ICSI) in andrological patients. J. Assist. Reprod. Genet. 1996, 13, 228–233. [Google Scholar] [CrossRef]
- Zini, A.; Finelli, A.; Phang, D.; Jarvi, K. Influence of semen processing technique on human sperm DNA integrity. Urology 2000, 56, 1081–1084. [Google Scholar] [CrossRef]
- Raad, G.; Bakos, H.W.; Bazzi, M.; Mourad, Y.; Fakih, F.; Shayya, S.; Mchantaf, L.; Fakih, C. Differential impact of four sperm preparation techniques on sperm motility, morphology, DNA fragmentation, acrosome status, oxidative stress, and mitochondrial activity: A prospective study. Andrology 2021, 9, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Silva, G.; López-Torres, A.S.; Maldonado-Rosas, I.; Mata-Martínez, E.; Larrea, F.; Torres-Flores, V.; Treviño, C.L.; Chirinos, M. Effects of Semen Processing on Sperm Function: Differences between Swim-Up and Density Gradient Centrifugation. World J. Mens. Health 2021, 39, 740–749. [Google Scholar] [CrossRef]
- Xue, X.; Wang, W.S.; Shi, J.Z.; Zhang, S.L.; Zhao, W.Q.; Shi, W.H.; Guo, B.Z.; Qin, Z. Efficacy of swim-up versus density gradient centrifugation in improving sperm deformity rate and DNA fragmentation index in semen samples from teratozoospermic patients. J. Assist. Reprod. Genet. 2014, 31, 1161–1166. [Google Scholar] [CrossRef] [Green Version]
- Muratori, M.; Tarozzi, N.; Carpentiero, F.; Danti, S.; Perrone, F.M.; Cambi, M.; Casini, A.; Azzari, C.; Boni, L.; Maggi, M.; et al. Sperm selection with density gradient centrifugation and swim up: Effect on DNA fragmentation in viable spermatozoa. Sci. Rep. 2019, 9, 7492. [Google Scholar] [CrossRef]
- De Martin, H.; Miranda, E.P.; Cocuzza, M.S.; Monteleone, P.A.A. Density gradient centrifugation and swim-up for ICSI: Useful, unsafe, or just unsuitable? J. Assist. Reprod. Genet. 2019, 36, 2421–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avendaño, C.; Oehninger, S. DNA fragmentation in morphologically normal spermatozoa: How much should we be concerned in the ICSI era? J. Androl. 2011, 32, 356–363. [Google Scholar] [CrossRef]
- Ezeh, U.I.; Moore, H.D.; Cooke, I.D. A prospective study of multiple needle biopsies versus a single open biopsy for testicular sperm extraction in men with non-obstructive azoospermia. Hum. Reprod. 1998, 13, 3075–3080. [Google Scholar] [CrossRef] [Green Version]
- Girardi, S.K.; Schlegel, P.N. Microsurgical epididymal sperm aspiration: Review of techniques, preoperative considerations, and results. J. Androl. 1996, 17, 5–9. [Google Scholar]
- Ozkocer, S.E.; Konac, E. The current perspective on genetic and epigenetic factors in sperm maturation in the epididymis. Andrologia 2021, 53, e13989. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, N.M.; Vaca Sánchez, R.; Rodriguez Fiesta, S.; Lopez Salgado, T.; Rodríguez, R.; Bethencourt, J.C.; Blanes Zamora, R. Pregnancy with frozen-thawed and fresh testicular biopsy after motile and immotile sperm microinjection, using the mechanical touch technique to assess viability. Hum. Reprod. 2004, 19, 262–265. [Google Scholar] [CrossRef] [Green Version]
- Soares, J.B.; Glina, S.; Galuppo, A.G. Pregnancy with frozen-thawed and fresh testicular biopsy after motile and immotile sperm microinjection, using the mechanical touch technique to assess viability. Hum. Reprod. 2005, 20, 569. [Google Scholar] [CrossRef] [Green Version]
- Rossato, M.; Di Virgilio, F.; Foresta, C. Involvement of osmo-sensitive calcium influx in human sperm activation. Mol. Hum. Reprod. 1996, 2, 903–909. [Google Scholar] [CrossRef] [Green Version]
- Nowicka-Bauer, K.; Szymczak-Cendlak, M. Structure and Function of Ion Channels Regulating Sperm Motility—An Overview. Int. J. Mol. Sci. 2021, 22, 3259. [Google Scholar] [CrossRef] [PubMed]
- Aydos, K.; Aydos, O.S. Sperm Selection Procedures for Optimizing the Outcome of ICSI in Patients with NOA. J. Clin. Med. 2021, 10, 2687. [Google Scholar] [CrossRef]
- Gianaroli, L.; Magli, M.C.; Collodel, G.; Moretti, E.; Ferraretti, A.P.; Baccetti, B. Sperm head’s birefringence: A new criterion for sperm selection. Fertil. Steril. 2008, 90, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Chattopadhyay, R.; Bose, G.; Ganesh, A.; Das, S.; Chakravarty, B.N. Selection of birefringent spermatozoa under Polscope: Effect on intracytoplasmic sperm injection outcome. Andrologia 2012, 44 (Suppl. 1), 734–738. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Role of sperm chromatin abnormalities and DNA damage in male infertility. Hum. Reprod. Update 2003, 9, 331–345. [Google Scholar] [CrossRef] [Green Version]
- González-Marín, C.; Gosálvez, J.; Roy, R. Types, causes, detection and repair of DNA fragmentation in animal and human sperm cells. Int. J. Mol. Sci. 2012, 13, 14026–14052. [Google Scholar] [CrossRef] [Green Version]
- Rappa, K.L.; Rodriguez, H.F.; Hakkarainen, G.C.; Anchan, R.M.; Mutter, G.L.; Asghar, W. Sperm processing for advanced reproductive technologies: Where are we today? Biotechnol. Adv. 2016, 34, 578–587. [Google Scholar] [CrossRef]
- Majzoub, A.; Agarwal, A.; Esteves, S.C. Clinical utility of sperm DNA damage in male infertility. Panminerva Med. 2019, 61, 118–127. [Google Scholar] [CrossRef]
- Oleszczuk, K.; Augustinsson, L.; Bayat, N.; Giwercman, A.; Bungum, M. Prevalence of high DNA fragmentation index in male partners of unexplained infertile couples. Andrology 2013, 1, 357–360. [Google Scholar] [CrossRef]
- Robinson, L.; Gallos, I.D.; Conner, S.J.; Rajkhowa, M.; Miller, D.; Lewis, S.; Kirkman-Brown, J.; Coomarasamy, A. The effect of sperm DNA fragmentation on miscarriage rates: A systematic review and meta-analysis. Hum. Reprod. 2012, 27, 2908–2917. [Google Scholar] [CrossRef] [Green Version]
- Pinto, S.; Carrageta, D.F.; Alves, M.G.; Rocha, A.; Agarwal, A.; Barros, A.; Oliveira, P.F. Sperm selection strategies and their impact on assisted reproductive technology outcomes. Andrologia 2021, 53, e13725. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Llanos, M.; Yovich, J.L.; Cummins, J.M.; Vigil, P. Pentoxifylline increases sperm penetration into zona-free hamster oocytes without increasing the acrosome reaction. Andrologia 1993, 25, 359–362. [Google Scholar] [CrossRef]
- Mahaldashtian, M.; Khalili, M.A.; Nottola, S.A.; Woodward, B.; Macchiarelli, G.; Miglietta, S. Does in vitro application of pentoxifylline have beneficial effects in assisted male reproduction? Andrologia 2021, 53, e13722. [Google Scholar] [CrossRef] [PubMed]
- Satish, M.; Kumari, S.; Deeksha, W.; Abhishek, S.; Nitin, K.; Adiga, S.K.; Hegde, P.; Dasappa, J.P.; Kalthur, G.; Rajakumara, E. Structure-based redesigning of pentoxifylline analogs against selective phosphodiesterases to modulate sperm functional competence for assisted reproductive technologies. Sci. Rep. 2021, 11, 12293. [Google Scholar] [CrossRef] [PubMed]
- Navas, P.; Paffoni, A.; Intra, G.; González-Utor, A.; Clavero, A.; Gonzalvo, M.C.; Díaz, R.; Peña, R.; Restelli, L.; Somigliana, E.; et al. Obstetric and neo-natal outcomes of ICSI cycles using pentoxifylline to identify viable spermatozoa in patients with immotile spermatozoa. Reprod. Biomed. Online 2017, 34, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.F.; Zhou, Q.W.; Zhang, S.P.; Lu, C.F.; Gong, F.; Shi, Y.; Lu, G.X.; Lin, G. The clinical and neonatal outcomes after stimulation of immotile spermatozoa using SperMagic medium. Andrologia 2018, 50, e13056. [Google Scholar] [CrossRef]
- Ibis, E.; Hayme, S.; Baysal, E.; Gul, N.; Ozkavukcu, S. Efficacy and safety of papaverine as an in vitro motility enhancer on human spermatozoa. J. Assist. Reprod. Genet. 2021, 38, 1523–1537. [Google Scholar] [CrossRef]
- Sandi-Monroy, N.L.; Musanovic, S.; Zhu, D.; Szabó, Z.; Vogl, A.; Reeka, N.; Eibner, K.; Bundschu, K.; Gagsteiger, F. Use of dimethylxanthine theophylline (SpermMobil®) does not affect clinical, obstetric or perinatal outcomes. Arch. Gynecol. Obstet. 2019, 300, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Mangoli, V.; Mangoli, R.; Dandekar, S.; Suri, K.; Desai, S. Selection of viable spermatozoa from testicular biopsies: A comparative study between pentoxifylline and hypoosmotic swelling test. Fertil. Steril. 2011, 95, 631–634. [Google Scholar] [CrossRef]
- Park, Y.J.; Pang, M.G. Mitochondrial Functionality in Male Fertility: From Spermatogenesis to Fertilization. Antioxidants 2021, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Ebner, T.; Moser, M.; Tews, G. Possible applications of a non-contact 1.48 microm wavelength diode laser in assisted reproduction technologies. Hum. Reprod. Update 2005, 11, 425–435. [Google Scholar] [CrossRef]
- Tarlatzis, B.C.; Kolibianakis, E.M.; Bontis, J.; Tousiou, M.; Lagos, S.; Mantalenakis, S. Effect of pentoxifylline on human sperm motility and fertilizing capacity. Arch. Androl. 1995, 34, 33–42. [Google Scholar] [CrossRef]
- Nordhoff, V.; Schüring, A.N.; Krallmann, C.; Zitzmann, M.; Schlatt, S.; Kiesel, L.; Kliesch, S. Optimizing TESE-ICSI by laser-assisted selection of immotile spermatozoa and polarization microscopy for selection of oocytes. Andrology 2013, 1, 67–74. [Google Scholar] [CrossRef]
- Neri, Q.V.; Lee, B.; Rosenwaks, Z.; Machaca, K.; Palermo, G.D. Understanding fertilization through intracytoplasmic sperm injection (ICSI). Cell Calcium. 2014, 55, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Guse, A.H.; Gil Montoya, D.C.; Diercks, B.P. Mechanisms and functions of calcium microdomains produced by ORAI channels, d-myo-inositol 1,4,5-trisphosphate receptors, or ryanodine receptors. Pharmacol. Ther. 2021, 223, 107804. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. The Inositol Trisphosphate/Calcium Signaling Pathway in Health and Disease. Physiol. Rev. 2016, 96, 1261–1296. [Google Scholar] [CrossRef] [Green Version]
- Ceyhan, Y.; Zhang, M.; Guo, J.; Sandoval, C.G.; Vacher, J.; Kaftanovskaya, E.M.; Agoulnik, A.I.; Agoulnik, I.U. Deletion of inositol polyphosphate 4-phosphatase type-II B affects spermatogenesis in mice. PLoS ONE 2020, 15, e0233163. [Google Scholar] [CrossRef]
- Abdolsamadi, M.; Mohammadi, F.; Nashtaei, M.S.; Teimouri, M.; Sardar, R.; Dayani, M.; Haghighi, M.; Ghasemi, S.; Vatannejad, A.; Zandieh, Z. Does myoinositol supplement improve sperm parameters and DNA integrity in patients with oligoasthenoteratozoospermia after the freezing-thawing process? Cell Tissue Bank. 2020, 21, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, R.A.; Barbagallo, F.; Calogero, A.E.; Cannarella, R.; Crafa, A.; La Vignera, S. D-Chiro-Inositol Improves Sperm Mitochondrial Membrane Potential: In Vitro Evidence. J. Clin. Med. 2020, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Rubino, P.; Palini, S.; Chigioni, S.; Carlomagno, G.; Quagliariello, A.; De Stefani, S.; Baglioni, A.; Bulletti, C. Improving fertilization rate in ICSI cycles by adding myoinositol to the semen preparation procedures: A prospective, bicentric, randomized trial on sibling oocytes. J. Assist. Reprod. Genet. 2015, 32, 387–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, S.; Xu, F.; Hirschfeld, M.; Brenig, B. Sperm Lipid Markers of Male Fertility in Mammals. Int. J. Mol. Sci. 2021, 22, 8767. [Google Scholar] [CrossRef]
- Hernández-Silva, G.; Fabián López-Araiza, J.E.; López-Torres, A.S.; Larrea, F.; Torres-Flores, V.; Chirinos, M. Proteomic characterization of human sperm plasma membrane-associated proteins and their role in capacitation. Andrology 2020, 8, 171–180. [Google Scholar] [CrossRef]
- Tumova, L.; Zigo, M.; Sutovsky, P.; Sedmikova, M.; Postlerova, P. Ligands and Receptors Involved in the Sperm-Zona Pellucida Interactions in Mammals. Cells 2021, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, M.; Nikkhoo, B.; Moradveisi, B.; Allahveisi, A. Effect of sperm selection methods on ICSI outcomes in patients with oligoteratozoospermia. Am. J. Clin. Exp. Urol. 2021, 9, 170–176. [Google Scholar]
- Izadi, M.; Khalili, M.A.; Salehi-Abargouei, A.; Rezvani, M.E.; Aflatoonian, B. Use of zona pellucida-bound spermatozoa as a natural selection in improvement of ICSI outcomes: A systematic review and meta-analysis. Andrologia 2021, 53, e14022. [Google Scholar] [CrossRef]
- Plouffe, B.D.; Murthy, S.K.; Lewis, L.H. Fundamentals and application of magnetic particles in cell isolation and enrichment: A review. Rep. Prog. Phys. 2015, 78, 016601. [Google Scholar] [CrossRef] [PubMed]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutelingsperger, C. A novel assay for apoptosis. Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin, V. J. Immunol. Methods 1995, 184, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Arends, M.J.; Wyllie, A.H. Apoptosis: Mechanisms and roles in pathology. Int. Rev. Exp. Pathol. 1991, 32, 223–254. [Google Scholar] [CrossRef] [PubMed]
- Tavalaee, M.; Deemeh, M.R.; Arbabian, M.; Nasr-Esfahani, M.H. Density gradient centrifugation before or after magnetic-activated cell sorting: Which technique is more useful for clinical sperm selection? J. Assist. Reprod. Genet. 2012, 29, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Dirican, E.K.; Ozgün, O.D.; Akarsu, S.; Akin, K.O.; Ercan, O.; Uğurlu, M.; Camsari, C.; Kanyilmaz, O.; Kaya, A.; Unsal, A. Clinical outcome of magnetic activated cell sorting of non-apoptotic spermatozoa before density gradient centrifugation for assisted reproduction. J. Assist. Reprod. Genet. 2008, 25, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Dandekar, P.; Aggeler, J.; Talbot, P. Structure, distribution and composition of the extracellular matrix of human oocytes and cumulus masses. Hum. Reprod. 1992, 7, 391–398. [Google Scholar] [CrossRef]
- Parmegiani, L.; Cognigni, G.E.; Ciampaglia, W.; Pocognoli, P.; Marchi, F.; Filicori, M. Efficiency of hyaluronic acid (HA) sperm selection. J. Assist. Reprod. Genet. 2010, 27, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Barak, Y.; Menezo, Y.; Veiga, A.; Elder, K. A physiological replacement for polyvinylpyrrolidone (PVP) in assisted reproductive technology. Hum. Fertil. 2001, 4, 99–103. [Google Scholar] [CrossRef]
- Parmegiani, L.; Cognigni, G.E.; Bernardi, S.; Troilo, E.; Ciampaglia, W.; Filicori, M. “Physiologic ICSI”: Hyaluronic acid (HA) favors selection of spermatozoa without DNA fragmentation and with normal nucleus, resulting in improvement of embryo quality. Fertil. Steril. 2010, 93, 598–604. [Google Scholar] [CrossRef]
- Yanagimachi, R.; Noda, Y.D.; Fujimoto, M.; Nicolson, G.L. The distribution of negative surface charges on mammalian spermatozoa. Am. J. Anat. 1972, 135, 497–519. [Google Scholar] [CrossRef] [PubMed]
- Zarei-Kheirabadi, M.; Shayegan Nia, E.; Tavalaee, M.; Deemeh, M.R.; Arabi, M.; Forouzanfar, M.; Javadi, G.R.; Nasr-Esfahani, M.H. Evaluation of ubiquitin and annexin V in sperm population selected based on density gradient centrifugation and zeta potential (DGC-Zeta). J. Assist. Reprod. Genet. 2012, 29, 365–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahedi, A.; Tavalaee, M.; Deemeh, M.R.; Azadi, L.; Fazilati, M.; Nasr-Esfahani, M.H. Zeta potential vs apoptotic marker: Which is more suitable for ICSI sperm selection? J. Assist. Reprod. Genet. 2013, 30, 1181–1186. [Google Scholar] [CrossRef] [Green Version]
- Bartoov, B.; Berkovitz, A.; Eltes, F. Selection of spermatozoa with normal nuclei to improve the pregnancy rate with intracytoplasmic sperm injection. N. Engl. J. Med. 2001, 345, 1067–1068. [Google Scholar] [CrossRef]
- Lo Monte, G.; Murisier, F.; Piva, I.; Germond, M.; Marci, R. Focus on intracytoplasmic morphologically selected sperm injection (IMSI): A mini-review. Asian J. Androl. 2013, 15, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Franco, J.G., Jr.; Baruffi, R.L.; Mauri, A.L.; Petersen, C.G.; Oliveira, J.B.; Vagnini, L. Significance of large nuclear vacuoles in human spermatozoa: Implications for ICSI. Reprod. Biomed. Online 2008, 17, 42–45. [Google Scholar] [CrossRef]
- Miki, K.; Clapham, D.E. Rheotaxis guides mammalian sperm. Curr. Biol. 2013, 23, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hino, T.; Yanagimachi, R. Active peristaltic movements and fluid production of the mouse oviduct: Their roles in fluid and sperm transport and fertilization. Biol. Reprod. 2019, 101, 40–49. [Google Scholar] [CrossRef]
- Nagata, M.P.B.; Endo, K.; Ogata, K.; Yamanaka, K.; Egashira, J.; Katafuchi, N.; Yamanouchi, T.; Matsuda, H.; Goto, Y.; Sakatani, M.; et al. Live births from artificial insemination of microfluidic-sorted bovine spermatozoa characterized by trajectories correlated with fertility. Proc. Natl. Acad. Sci. USA 2018, 115, E3087–E3096. [Google Scholar] [CrossRef] [Green Version]
- De Martin, H.; Cocuzza, M.S.; Tiseo, B.C.; Wood, G.J.A.; Miranda, E.P.; Monteleone, P.A.A.; Soares, J.M., Jr.; Serafini, P.C.; Srougi, M.; Baracat, E.C. Positive rheotaxis extended drop: A one-step procedure to select and recover sperm with mature chromatin for intracytoplasmic sperm injection. J. Assist. Reprod. Genet. 2017, 34, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Zaferani, M.; Cheong, S.H.; Abbaspourrad, A. Rheotaxis-based separation of sperm with progressive motility using a microfluidic corral system. Proc. Natl. Acad. Sci. USA 2018, 115, 8272–8277. [Google Scholar] [CrossRef] [Green Version]
- Oren-Benaroya, R.; Orvieto, R.; Gakamsky, A.; Pinchasov, M.; Eisenbach, M. The sperm chemoattractant secreted from human cumulus cells is progesterone. Hum. Reprod. 2008, 23, 2339–2345. [Google Scholar] [CrossRef] [Green Version]
- Gatica, L.V.; Guidobaldi, H.A.; Montesinos, M.M.; Teves, M.E.; Moreno, A.I.; Uñates, D.R.; Molina, R.I.; Giojalas, L.C. Picomolar gradients of progesterone select functional human sperm even in subfertile samples. Mol. Hum. Reprod. 2013, 19, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, R.; Ni, Y.; Sun, P.; Liu, Y.; Zhang, D.; Huang, H. Novel distance-progesterone-combined selection approach improves human sperm quality. J. Transl. Med. 2018, 16, 203. [Google Scholar] [CrossRef]
- Bahat, A.; Eisenbach, M. Sperm thermotaxis. Mol. Cell Endocrinol. 2006, 252, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Bahat, A.; Tur-Kaspa, I.; Gakamsky, A.; Giojalas, L.C.; Breitbart, H.; Eisenbach, M. Thermotaxis of mammalian sperm cells: A potential navigation mechanism in the female genital tract. Nat Med. 2003, 9, 149–150. [Google Scholar] [CrossRef]
- Bahat, A.; Caplan, S.R.; Eisenbach, M. Thermotaxis of human sperm cells in extraordinarily shallow temperature gradients over a wide range. PLoS ONE 2012, 7, e41915. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Cerezales, S.; Laguna-Barraza, R.; de Castro, A.C.; Sánchez-Calabuig, M.J.; Cano-Oliva, E.; de Castro-Pita, F.J.; Montoro-Buils, L.; Pericuesta, E.; Fernández-González, R.; Gutiérrez-Adán, A. Sperm selection by thermotaxis improves ICSI outcome in mice. Sci. Rep. 2018, 8, 2902. [Google Scholar] [CrossRef]
- Sackmann, E.K.; Fulton, A.L.; Beebe, D.J. The present and future role of microfluidics in biomedical research. Nature 2014, 507, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Shirota, K.; Yotsumoto, F.; Itoh, H.; Obama, H.; Hidaka, N.; Nakajima, K.; Miyamoto, S. Separation efficiency of a microfluidic sperm sorter to minimize sperm DNA damage. Fertil. Steril. 2016, 105, 315–321.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.; Xie, L.; Han, C.; Su, K.; Qiu, T.; Wang, L.; Huang, G.; Xing, W.; Qiao, J.; Wang, J.; et al. In vitro fertilization on a single-oocyte positioning system integrated with motile sperm selection and early embryo development. Anal. Chem. 2011, 83, 2964–2970. [Google Scholar] [CrossRef]
- Simon, L.; Murphy, K.; Aston, K.I.; Emery, B.R.; Hotaling, J.M.; Carrell, D.T. Optimization of microelectrophoresis to select highly negatively charged sperm. J. Assist. Reprod. Genet. 2016, 33, 679–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, Y.J.; Maeng, J.H.; Hwang, S.Y.; Ahn, Y. Design, Fabrication, and Testing of a Microfluidic Device for Thermotaxis and Chemotaxis Assays of Sperm. SLAS Technol. 2018, 23, 507–515. [Google Scholar] [CrossRef]
- Bhagwat, S.; Sontakke, S.K.D.; Parte, P.; Jadhav, S. Chemotactic behavior of spermatozoa captured using a microfluidic chip. Biomicrofluidics 2018, 12, 024112. [Google Scholar] [CrossRef]
- Ferraz, M.A.M.M.; Rho, H.S.; Hemerich, D.; Henning, H.H.W.; van Tol, H.T.A.; Hölker, M.; Besenfelder, U.; Mokry, M.; Vos, P.L.A.M.; Stout, T.A.E.; et al. An oviduct-on-a-chip provides an enhanced in vitro environment for zygote genome reprogramming. Nat. Commun. 2018, 9, 4934. [Google Scholar] [CrossRef] [Green Version]
- Eravuchira, P.J.; Mirsky, S.K.; Barnea, I.; Levi, M.; Balberg, M.; Shaked, N.T. Individual sperm selection by microfluidics integrated with interferometric phase microscopy. Methods 2018, 136, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Amaral, S.; Da Costa, R.; Wübbeling, F.; Redmann, K.; Schlatt, S. Raman micro-spectroscopy analysis of different sperm regions: A species comparison. Mol. Hum. Reprod. 2018, 24, 185–202. [Google Scholar] [CrossRef]
- Di Caprio, G.; Ferrara, M.A.; Miccio, L.; Merola, F.; Memmolo, P.; Ferraro, P.; Coppola, G. Holographic imaging of unlabelled sperm cells for semen analysis: A review. J. Biophotonics 2015, 8, 779–789. [Google Scholar] [CrossRef] [Green Version]
- de Wagenaar, B.; Geijs, D.J.; de Boer, H.; Bomer, J.G.; Olthuis, W.; van den Berg, A.; Segerink, L.I. Spermometer: Electrical characterization of single boar sperm motility. Fertil. Steril. 2016, 106, 773–780.e6. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic representation of swim-up and density gradient centrifugation.

Figure 2.
Schematic representation Hypo-osmotic swelling Test. In this figure, we observe the difference between the abnormal sperm with not swelled tail and the normal sperm with swelled tail after treatment with a hypo-osmotic solution.
Figure 2.
Schematic representation Hypo-osmotic swelling Test. In this figure, we observe the difference between the abnormal sperm with not swelled tail and the normal sperm with swelled tail after treatment with a hypo-osmotic solution.

Figure 3.
Microscope implemented with polarized light. The birefringence of the heads is clear in the viable sperm (yellow heads) compared to the not viable one where the birefringence is absent (dark heads).
Figure 3.
Microscope implemented with polarized light. The birefringence of the heads is clear in the viable sperm (yellow heads) compared to the not viable one where the birefringence is absent (dark heads).

Figure 4.
Schematic representation of chemical inducers of motility. On the left we can recognize the initial immotile sperms and on the right side the viable motile sperms activated by the chemical inducer.
Figure 4.
Schematic representation of chemical inducers of motility. On the left we can recognize the initial immotile sperms and on the right side the viable motile sperms activated by the chemical inducer.

Figure 5.
Schematic representation of LAISS (laser-assisted immotile sperm selection). The laser irradiation generates a slight movement of the tail in those viable sperms initially immotile.
Figure 5.
Schematic representation of LAISS (laser-assisted immotile sperm selection). The laser irradiation generates a slight movement of the tail in those viable sperms initially immotile.

Figure 6.
Sperm motility enhanced by ATP/MgSo4 or myo-inositol. Briefly in the picture we observe the immotile sperm before the treatment with the chemical inducer (left side) and the motile sperm cells after the chemical exposure (right side).
Figure 6.
Sperm motility enhanced by ATP/MgSo4 or myo-inositol. Briefly in the picture we observe the immotile sperm before the treatment with the chemical inducer (left side) and the motile sperm cells after the chemical exposure (right side).

Figure 7.
Sperm cell passing through column with annexin V. The sperm cells able to pass through the column represent the viable one (green heads) while the apoptotic fraction remain trapped inside the column (yellow heads).
Figure 7.
Sperm cell passing through column with annexin V. The sperm cells able to pass through the column represent the viable one (green heads) while the apoptotic fraction remain trapped inside the column (yellow heads).

Figure 8.
Schematic representation of the Zeta method. The high negative charge of the mature sperm cell membrane reacts with the positive charge of the tube while the immature fraction is detached form the tube.
Figure 8.
Schematic representation of the Zeta method. The high negative charge of the mature sperm cell membrane reacts with the positive charge of the tube while the immature fraction is detached form the tube.

Figure 9.
Sperm cell visualization with magnification system 6000×. Sperm cells with appropriate morphology are well selected through the high magnification.
Figure 9.
Sperm cell visualization with magnification system 6000×. Sperm cells with appropriate morphology are well selected through the high magnification.

Figure 10.
Schematic representation of the microfluidic system. The motile fraction of sperm cells is able to swim through the flow and be collected in separate chambers (blue chambers) while the immotile sperm cells (yellow heads) and debris reach the exit of the microfluidic system (dark chamber).
Figure 10.
Schematic representation of the microfluidic system. The motile fraction of sperm cells is able to swim through the flow and be collected in separate chambers (blue chambers) while the immotile sperm cells (yellow heads) and debris reach the exit of the microfluidic system (dark chamber).

Figure 11.
Top and Side viewing of the sperm cells horizontal migration from the first drop (where the cells are added) to the third drop (where the sperm cells are aspirated) through 2 bridges that link them.
Figure 11.
Top and Side viewing of the sperm cells horizontal migration from the first drop (where the cells are added) to the third drop (where the sperm cells are aspirated) through 2 bridges that link them.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
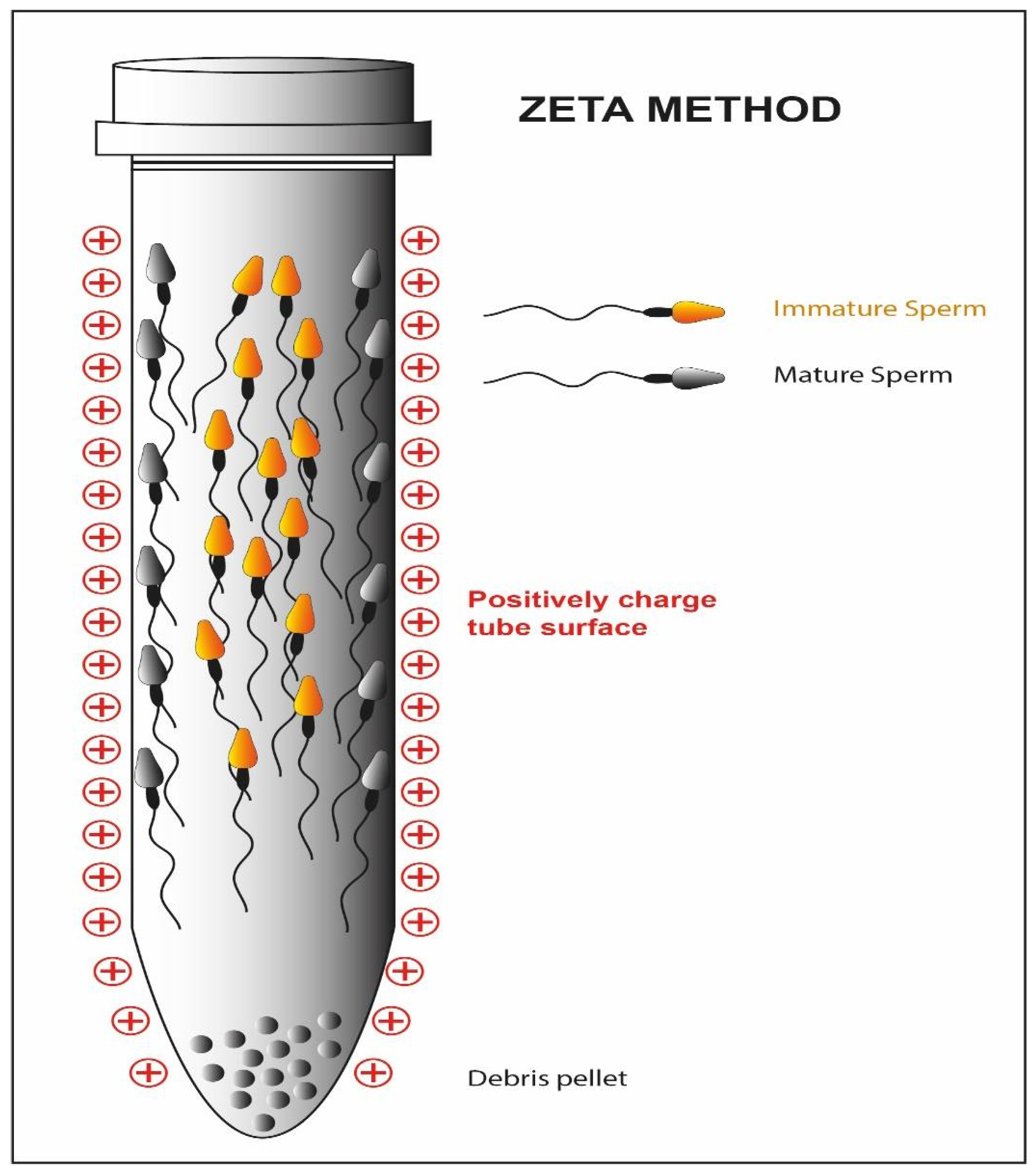{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Advantages and disadvantages of sperm cells’ selection techniques.
| Procedures | Advantages | Disadvantages |
|---|---|---|
| Swim-up | ||
| Centrifugation on density gradient | ||
| HOST | ||
| Polarization microscopy |
| |
| LAISS | ||
| MACS |
|
|
| PICSI and selection with hyaluronic acid |
|
|
| Zeta potential | ||
| IMSI |
| |
| Microfluidic separation |
| |
| Horizontal Sperm migration |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Baldini, D.; Ferri, D.; Baldini, G.M.; Lot, D.; Catino, A.; Vizziello, D.; Vizziello, G. Sperm Selection for ICSI: Do We Have a Winner? Cells 2021, 10, 3566. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123566
AMA Style
Baldini D, Ferri D, Baldini GM, Lot D, Catino A, Vizziello D, Vizziello G. Sperm Selection for ICSI: Do We Have a Winner? Cells. 2021; 10(12):3566. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123566
Chicago/Turabian StyleBaldini, Domenico, Daniele Ferri, Giorgio Maria Baldini, Dario Lot, Assunta Catino, Damiano Vizziello, and Giovanni Vizziello. 2021. "Sperm Selection for ICSI: Do We Have a Winner?" Cells 10, no. 12: 3566. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10123566
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.