
Lithium Chloride Protects against Sepsis-Induced Skeletal Muscle Atrophy and Cancer Cachexia


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Collection of Conditioned Media
2.4. MTT Assay for Cell Proliferation
2.5. Flow cytometry
2.6. siRNA-Mediated Gene Knockdown
2.7. Real-Time Quantitative PCR
2.8. In Vitro Model of Cancer Cachexia and Morphological Analysis of Myotubes
2.9. Immunoblotting
2.10. Assessment of Pro-Cachexia Cytokine Induction in Macrophages
2.11. Animal Studies
2.12. Animal Model of Sepsis-Induced Muscle Wasting
2.13. Animal Model of Cancer Cachexia
2.14. Histological Analysis
2.15. Statistics
3. Results
3.1. LiCl Enhanced Myogenic Differentiation
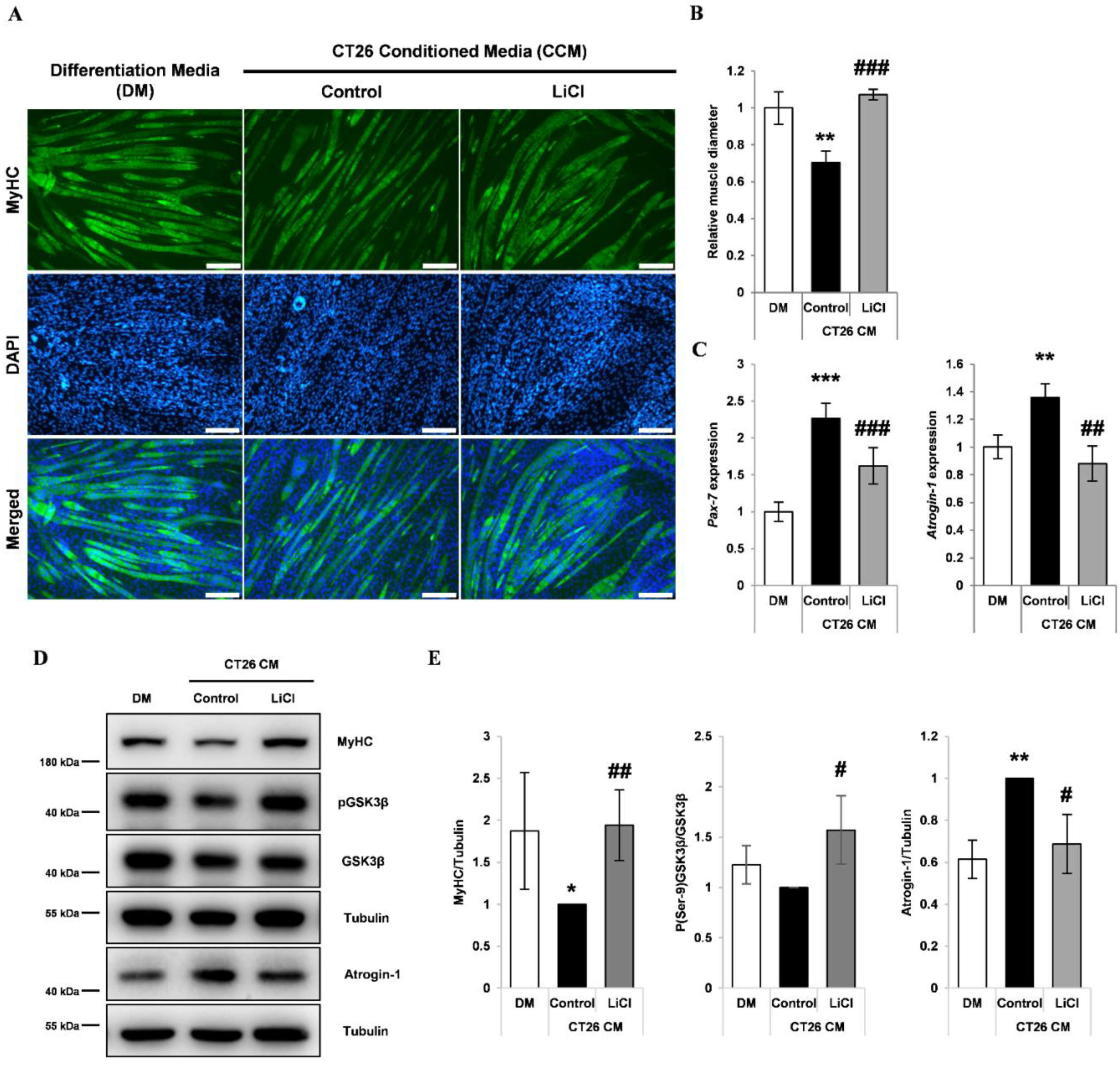
3.2. LiCl Increased Myh2 Expression and Reduced Pax-7 Expression in Differentiating Myoblasts Treated with CCM
3.3. LiCl Prevented CCM-Induced Myotube Wasting
3.4. Lithium Mimetic Ebselen Did Not Prevent Myotube Wasting Induced by CCM
3.5. LiCl Inhibited LPS-Induced Inflammatory Cytokine Production
3.6. LiCl Prevented Muscle Wasting in a Mouse Model of Septic Cachexia
3.7. LiCl Treatment Attenuated Muscle Wasting in Cancer Cachexia
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mourkioti, F.; Rosenthal, N. NF-kappaB signaling in skeletal muscle: Prospects for intervention in muscle diseases. J. Mol. Med. 2008, 86, 747–759. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Liu, Z.; Ding, H.; Zhou, Y.; Doan, H.A.; Sin, K.W.T.; Zhu, Z.J.; Flores, R.; Wen, Y.; Gong, X.; et al. Tumor induces muscle wasting in mice through releasing extracellular Hsp70 and Hsp90. Nat. Commun. 2017, 8, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadi, A.M.; Mouchaers, K.T.; Schalij, I.; Grunberg, K.; Meijer, G.A.; Vonk-Noordegraaf, A.; van der Laarse, W.J.; Belien, J.A. Rapid quantification of myocardial fibrosis: A new macro-based automated analysis. Anal. Cell. Pathol. 2010, 33, 257–269. [Google Scholar] [CrossRef]
- Stephens, N.A.; Skipworth, R.J.; Fearon, K.C. Cachexia, survival and the acute phase response. Curr. Opin. Support. Palliat. Care 2008, 2, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Hahn, A.; Kny, M.; Pablo-Tortola, C.; Todiras, M.; Willenbrock, M.; Schmidt, S.; Schmoeckel, K.; Jorde, I.; Nowak, M.; Jarosch, E.; et al. Serum amyloid A1 mediates myotube atrophy via Toll-like receptors. J. Cachexia Sarcopenia Muscle 2020, 11, 103–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhamarneh, O.; Agada, F.; Madden, L.; Stafford, N.; Greenman, J. Serum IL10 and circulating CD4(+) CD25(high) regulatory T cell numbers as predictors of clinical outcome and survival in patients with head and neck squamous cell carcinoma. Head Neck 2011, 33, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.A.; O’Brien, J.M., Jr.; Hoffmann, S.P.; Phillips, G.; Garland, A.; Finley, J.C.; Almoosa, K.; Hejal, R.; Wolf, K.M.; Lemeshow, S.; et al. Acquired weakness, handgrip strength, and mortality in critically ill patients. Am. J. Respir. Critcal Care Med. 2008, 178, 261–268. [Google Scholar] [CrossRef]
- Penet, M.F.; Bhujwalla, Z.M. Cancer cachexia, recent advances, and future directions. Cancer J. 2015, 21, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Argiles, J.M.; Busquets, S.; Lopez-Soriano, F.J.; Costelli, P.; Penna, F. Are there any benefits of exercise training in cancer cachexia? J. Cachexia Sarcopenia Muscle 2012, 3, 73–76. [Google Scholar] [CrossRef] [Green Version]
- De Jonghe, B.; Sharshar, T.; Lefaucheur, J.P.; Authier, F.J.; Durand-Zaleski, I.; Boussarsar, M.; Cerf, C.; Renaud, E.; Mesrati, F.; Carlet, J.; et al. Paresis acquired in the intensive care unit: A prospective multicenter study. JAMA 2002, 288, 2859–2867. [Google Scholar] [CrossRef] [Green Version]
- Won, E.; Kim, Y.K. An Oldie but Goodie: Lithium in the Treatment of Bipolar Disorder through Neuroprotective and Neurotrophic Mechanisms. Int. J. Mol. Sci. 2017, 18, 2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shorter, E. The history of lithium therapy. Bipolar Disord. 2009, 11 (Suppl. S2), 4–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKnight, R.F.; Chesney, E.; Amit, B.H.; Geddes, J.; Cipriani, A. Lithium for acute mania. Cochrane Database Syst. Rev. 2019, 6, CD004048. [Google Scholar] [CrossRef]
- Xu, M.; Wang, S.L.; Zhu, L.; Wu, P.Y.; Dai, W.B.; Rakesh, K.P. Structure-activity relationship (SAR) studies of synthetic glycogen synthase kinase-3beta inhibitors: A critical review. Eur. J. Med. Chem. 2019, 164, 448–470. [Google Scholar] [CrossRef]
- Malhi, G.S.; Tanious, M.; Das, P.; Coulston, C.M.; Berk, M. Potential mechanisms of action of lithium in bipolar disorder. Current understanding. CNS Drugs 2013, 27, 135–153. [Google Scholar] [CrossRef]
- Maurer, I.C.; Schippel, P.; Volz, H.P. Lithium-induced enhancement of mitochondrial oxidative phosphorylation in human brain tissue. Bipolar Disord. 2009, 11, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Carli, M.; Anand-Srivastava, M.B.; Molina-Holgado, E.; Dewar, K.M.; Reader, T.A. Effects of chronic lithium treatments on central dopaminergic receptor systems: G proteins as possible targets. Neurochem. Int. 1994, 24, 13–22. [Google Scholar] [CrossRef]
- Hokin, L.E.; Dixon, J.F.; Los, G.V. A novel action of lithium: Stimulation of glutamate release and inositol 1,4,5 trisphosphate accumulation via activation of the N-methyl D-aspartate receptor in monkey and mouse cerebral cortex slices. Adv. Enzym. Regul. 1996, 36, 229–244. [Google Scholar] [CrossRef]
- Brunello, N.; Tascedda, F. Cellular mechanisms and second messengers: Relevance to the psychopharmacology of bipolar disorders. Int. J. Neuropsychopharmacol. 2003, 6, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Halliday, A.C.; Thomas, J.M.; Kuznetsova, O.V.; Baldwin, R.; Woon, E.C.; Aley, P.K.; Antoniadou, I.; Sharp, T.; Vasudevan, S.R.; et al. A safe lithium mimetic for bipolar disorder. Nat. Commun. 2013, 4, 1332. [Google Scholar] [CrossRef] [Green Version]
- Alda, M. Lithium in the treatment of bipolar disorder: Pharmacology and pharmacogenetics. Mol. Psychiatry 2015, 20, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Kazemi, H.; Noori-Zadeh, A.; Darabi, S.; Rajaei, F. Lithium prevents cell apoptosis through autophagy induction. Bratisl. Lek. Listy 2018, 119, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.C.; Shih, S.R.; Chang, T.Y.; Fang, M.Y.; Hsu, J.T. The combination effects of licl and the active leflunomide metabolite, A771726, on viral-induced interleukin 6 production and EV-A71 replication. PLoS ONE 2014, 9, e111331. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.L.; Yasmin, H.; Varone, A.; Wiles, A.; Poole, C.A.; Knight, M.M. Lithium chloride prevents interleukin-1beta induced cartilage degradation and loss of mechanical properties. J. Orthop. Res. 2015, 33, 1552–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttuso, T., Jr. High lithium levels in tobacco may account for reduced incidences of both Parkinson’s disease and melanoma in smokers through enhanced beta-catenin-mediated activity. Med. Hypotheses 2019, 131, 109302. [Google Scholar] [CrossRef]
- Verma, Y.K.; Singh, A.K.; Gurudutta, G.U. Survival genes expression analysis following ionizing radiation to LiCl treated KG1a cells. Int. J. Radiat. Biol. 2020, 96, 671–688. [Google Scholar] [CrossRef]
- Kurgan, N.; Whitley, K.C.; Maddalena, L.A.; Moradi, F.; Stoikos, J.; Hamstra, S.I.; Rubie, E.A.; Kumar, M.; Roy, B.D.; Woodgett, J.R.; et al. A Low-Therapeutic Dose of Lithium Inhibits GSK3 and Enhances Myoblast Fusion in C2C12 Cells. Cells 2019, 8, 1340. [Google Scholar] [CrossRef] [Green Version]
- Whitley, K.C.; Hamstra, S.I.; Baranowski, R.W.; Watson, C.J.F.; MacPherson, R.E.K.; MacNeil, A.J.; Roy, B.D.; Vandenboom, R.; Fajardo, V.A. GSK3 inhibition with low dose lithium supplementation augments murine muscle fatigue resistance and specific force production. Physiol. Rep. 2020, 8, e14517. [Google Scholar] [CrossRef]
- Findlay, A.R.; Bengoechea, R.; Pittman, S.K.; Chou, T.F.; True, H.L.; Weihl, C.C. Lithium chloride corrects weakness and myopathology in a preclinical model of LGMD1D. Neurol. Genet. 2019, 5, e318. [Google Scholar] [CrossRef] [Green Version]
- Bertsch, S.; Lang, C.H.; Vary, T.C. Inhibition of glycogen synthase kinase 3[beta] activity with lithium in vitro attenuates sepsis-induced changes in muscle protein turnover. Shock 2011, 35, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Sonni, P.V.; Iannuzzi, S.; Aversa, Z.; Tommasi, V.; Frascaria, T.; Costelli, P.; Fanelli, F.R.; Muscaritoli, M. Effect of lithium administration on muscle and body weight loss in experimental cancer cachexia. Eur. J. Intern. Med. 2008, 19, S16. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Cho, H.; Seo, Y.; Loke, K.M.; Kim, S.W.; Oh, S.M.; Kim, J.H.; Soh, J.; Kim, H.S.; Lee, H.; Kim, J.; et al. Cancer-Stimulated CAFs Enhance Monocyte Differentiation and Protumoral TAM Activation via IL6 and GM-CSF Secretion. Clin. Cancer Res. 2018, 24, 5407–5421. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.X.; Rudnicki, M.A. Satellite cells, the engines of muscle repair. Nat. Rev. Mol. Cell Biol. 2011, 13, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Rommel, C.; Bodine, S.C.; Clarke, B.A.; Rossman, R.; Nunez, L.; Stitt, T.N.; Yancopoulos, G.D.; Glass, D.J. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nat. Cell Biol. 2001, 3, 1009–1013. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.D.; Lecker, S.H.; Jagoe, R.T.; Navon, A.; Goldberg, A.L. Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. Proc. Nat. Acad. Sci. USA 2001, 98, 14440–14445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Hung, Y.L.; Fang, S.H.; Wang, S.C.; Cheng, W.C.; Liu, P.L.; Su, C.C.; Chen, C.S.; Huang, M.Y.; Hua, K.F.; Shen, K.H.; et al. Corylin protects LPS-induced sepsis and attenuates LPS-induced inflammatory response. Sci. Rep. 2017, 7, 46299. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.; Im, M.; Gu, M.J.; Ma, J.Y. Citrus unshiu peel extract alleviates cancer-induced weight loss in mice bearing CT-26 adenocarcinoma. Sci. Rep. 2016, 6, 24214. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Kishimoto, H.; Chua, H.L.; Badve, S.; Miller, K.D.; Bigsby, R.M.; Nakshatri, H. Interleukin-1 alpha promotes tumor growth and cachexia in MCF-7 xenograft model of breast cancer. Am. J. Pathol. 2003, 163, 2531–2541. [Google Scholar] [CrossRef]
- Narsale, A.A.; Carson, J.A. Role of interleukin-6 in cachexia: Therapeutic implications. Curr. Opin. Support. Palliat. Care 2014, 8, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, D.T.; Ma, J.F.; Marco, S.D.; Gallouzi, I.E. Inducible nitric oxide synthase (iNOS) in muscle wasting syndrome, sarcopenia, and cachexia. Aging 2011, 3, 702–715. [Google Scholar] [CrossRef] [Green Version]
- Braun, T.P.; Grossberg, A.J.; Krasnow, S.M.; Levasseur, P.R.; Szumowski, M.; Zhu, X.X.; Maxson, J.E.; Knoll, J.G.; Barnes, A.P.; Marks, D.L. Cancer- and endotoxin-induced cachexia require intact glucocorticoid signaling in skeletal muscle. FASEB J. 2013, 27, 3572–3582. [Google Scholar] [CrossRef] [Green Version]
- Can, A.; Blackwell, R.A.; Piantadosi, S.C.; Dao, D.T.; O’Donnell, K.C.; Gould, T.D. Antidepressant-like responses to lithium in genetically diverse mouse strains. Genes Brain Behav. 2011, 10, 434–443. [Google Scholar] [CrossRef] [Green Version]
- Bao, H.; Ge, Y.; Wang, Z.; Zhuang, S.; Dworkin, L.; Peng, A.; Gong, R. Delayed administration of a single dose of lithium promotes recovery from AKI. J. Am. Soc. Nephrol. 2014, 25, 488–500. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Huang, W.C.; Wang, C.Y.; Tsai, C.C.; Chen, C.L.; Chang, Y.T.; Kai, J.I.; Lin, C.F. Inhibiting glycogen synthase kinase-3 reduces endotoxaemic acute renal failure by down-regulating inflammation and renal cell apoptosis. Br. J. Pharmacol. 2009, 157, 1004–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazlovitskaya, E.M.; Edwards, E.; Thotala, D.; Fu, A.; Osusky, K.L.; Whetsell, W.O., Jr.; Boone, B.; Shinohara, E.T.; Hallahan, D.E. Lithium treatment prevents neurocognitive deficit resulting from cranial irradiation. Cancer Res. 2006, 66, 11179–11186. [Google Scholar] [CrossRef] [Green Version]
- Buck, M.; Chojkier, M. Muscle wasting and dedifferentiation induced by oxidative stress in a murine model of cachexia is prevented by inhibitors of nitric oxide synthesis and antioxidants. EMBO J. 1996, 15, 1753–1765. [Google Scholar] [CrossRef] [PubMed]
- Hall, D.T.; Griss, T.; Ma, J.F.; Sanchez, B.J.; Sadek, J.; Tremblay, A.M.K.; Mubaid, S.; Omer, A.; Ford, R.J.; Bedard, N.; et al. The AMPK agonist 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), but not metformin, prevents inflammation-associated cachectic muscle wasting. EMBO Mol. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Miyazaki, H.; Takeda, Y.; Takeo, S. Detection of serum cytokine levels in experimental cancer cachexia of colon 26 adenocarcinoma-bearing mice. Cancer Lett. 1993, 72, 65–70. [Google Scholar] [CrossRef]
- Evans, W.J.; Morley, J.E.; Argiles, J.; Bales, C.; Baracos, V.; Guttridge, D.; Jatoi, A.; Kalantar-Zadeh, K.; Lochs, H.; Mantovani, G.; et al. Cachexia: A new definition. Clin. Nutr. 2008, 27, 793–799. [Google Scholar] [CrossRef]
- Strassmann, G.; Fong, M.; Kenney, J.S.; Jacob, C.O. Evidence for the involvement of interleukin 6 in experimental cancer cachexia. J. Clin. Investig. 1992, 89, 1681–1684. [Google Scholar] [CrossRef] [Green Version]
- Strassmann, G.; Fong, M.; Freter, C.E.; Windsor, S.; D’Alessandro, F.; Nordan, R.P. Suramin interferes with interleukin-6 receptor binding in vitro and inhibits colon-26-mediated experimental cancer cachexia in vivo. J. Clin. Investig. 1993, 92, 2152–2159. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Von Haehling, S.; Springer, J. Promising models for cancer-induced cachexia drug discovery. Expert Opin. Drug Discov. 2020, 15, 627–637. [Google Scholar] [CrossRef]
- Cho, D.S.; Schmitt, R.E.; Dasgupta, A.; Ducharme, A.M.; Doles, J.D. Single-cell deconstruction of post-sepsis skeletal muscle and adipose tissue microenvironments. J. Cachexia Sarcopenia Muscle 2020, 11, 1351–1363. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Zhong, Z.; Zheng, Z.; Shi, X.M.; Zhang, W. Inhibition of glycogen synthase kinase-3beta attenuates glucocorticoid-induced suppression of myogenic differentiation in vitro. PLoS ONE 2014, 9, e105528. [Google Scholar] [CrossRef]
- Abu-Baker, A.; Laganiere, J.; Gaudet, R.; Rochefort, D.; Brais, B.; Neri, C.; Dion, P.A.; Rouleau, G.A. Lithium chloride attenuates cell death in oculopharyngeal muscular dystrophy by perturbing Wnt/beta-catenin pathway. Cell Death Dis. 2013, 4, e821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Li, S.; Duan, X.; Gu, Z.; Ma, Z.; Yuan, X.; Feng, X.; Wang, H. Inhibition of glycogen synthase kinase 3 beta (GSK3beta) suppresses the progression of esophageal squamous cell carcinoma by modifying STAT3 activity. Mol. Carcinog. 2017, 56, 2301–2316. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Chen, Y.; Niu, Y.; Chen, W.; Wang, Q.; Xiao, S.; Li, A.; Xie, Y.; Li, J.; Zhao, X.; et al. A deep investigation into the adipogenesis mechanism: Profile of microRNAs regulating adipogenesis by modulating the canonical Wnt/beta-catenin signaling pathway. BMC Genom. 2010, 11, 320. [Google Scholar] [CrossRef] [Green Version]
- Eldar-Finkelman, H.; Schreyer, S.A.; Shinohara, M.M.; LeBoeuf, R.C.; Krebs, E.G. Increased glycogen synthase kinase-3 activity in diabetes- and obesity-prone C57BL/6J mice. Diabetes 1999, 48, 1662–1666. [Google Scholar] [CrossRef]
- Tomasin, R.; Martin, A.; Cominetti, M.R. Metastasis and cachexia: Alongside in clinics, but not so in animal models. J. Cachexia Sarcopenia Muscle 2019, 10, 1183–1194. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, W.T.; Klein, P.S. Validating GSK3 as an in vivo target of lithium action. Biochem. Soc. Trans. 2009, 37, 1133–1138. [Google Scholar] [CrossRef] [Green Version]
- Belmaker, R.H.; Bersudsky, Y.; Agam, G.; Levine, J.; Kofman, O. How does lithium work on manic depression? Clinical and psychological correlates of the inositol theory. Annu. Rev. Med. 1996, 47, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Khasraw, M.; Ashley, D.; Wheeler, G.; Berk, M. Using lithium as a neuroprotective agent in patients with cancer. BMC Med. 2012, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Van der Werf, A.; van Bokhorst, Q.N.E.; de van der Schueren, M.A.E.; Verheul, H.M.W.; Langius, J.A.E. Cancer Cachexia: Identification by Clinical Assessment versus International Consensus Criteria in Patients with Metastatic Colorectal Cancer. Nutr. Cancer 2018, 70, 1322–1329. [Google Scholar] [CrossRef] [PubMed]
- Argiles, J.M.; Lopez-Soriano, F.J.; Stemmler, B.; Busquets, S. Therapeutic strategies against cancer cachexia. Eur. J. Transl. Myol. 2019, 29, 7960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsady, M.; Baumgarten, R.; Deen, P.M.; de Groot, T. Lithium in the Kidney: Friend and Foe? J. Am. Soc. Nephrol. 2016, 27, 1587–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinsson, L.; Westman, J.; Hallgren, J.; Osby, U.; Backlund, L. Lithium treatment and cancer incidence in bipolar disorder. Bipolar Disord. 2016, 18, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarse, K.; Terao, T.; Tian, J.; Iwata, N.; Ishii, N.; Ristow, M. Low-dose lithium uptake promotes longevity in humans and metazoans. Eur. J. Nutr. 2011, 50, 387–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Locus | Source | Primer Sequence | Size | |
|---|---|---|---|---|---|
| Myh2 | NM_001039545 | Mus musculus | Forward | GAAGAGCCGGGAGGTTCAC | 113 bp |
| Reverse | ACACAGGCGCATGACCAAA | ||||
| Pax-7 | NM_011039 | Mus musculus | Forward | CCCTTTCAAAGACCAAATGCA | 198 bp |
| Reverse | CCCTCACGGGCAGATCATTA | ||||
| Atrogin-1 | NM_026346 | Mus musculus | Forward | CAGAGAGCTGCTCCGTCTCA | 178 bp |
| Reverse | ACGTATCCCCCGCAGTTTC | ||||
| Murf-1 | NM_001039048 | Mus musculus | Forward | CCGAGTGCAGACGATCATCTC | 198 bp |
| Reverse | TGGAGGATCAGAGCCTCGAT | ||||
| Il-6 | NM_031168 | Mus musculus | Forward | GAGGATACCACTCCCAAC | 141 bp |
| Reverse | AAGTGCATCATCGTTGTT | ||||
| inos | NM_010927 | Mus musculus | Forward | CCCCTTCAATGGCTGGTACA | 64 bp |
| Reverse | GCGCTGGACGTCACAGAA | ||||
| Il-1β | NM_008361 | Mus musculus | Forward | TGCCACCTTTTGACAGTGATG | 135 bp |
| Reverse | TGTGCTGCTGCGAGATTTGA | ||||
| Gapdh | NM_001289726 | Mus musculus | Forward | CTCCACTCACGGCAAATTCA | 120 bp |
| Reverse | GCCTCACCCCATTTGATGTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-H.; Kim, S.-W.; Kim, J.-H.; Kim, H.-J.; Um, J.; Jung, D.-W.; Williams, D.R. Lithium Chloride Protects against Sepsis-Induced Skeletal Muscle Atrophy and Cancer Cachexia. Cells 2021, 10, 1017. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051017
Lee J-H, Kim S-W, Kim J-H, Kim H-J, Um J, Jung D-W, Williams DR. Lithium Chloride Protects against Sepsis-Induced Skeletal Muscle Atrophy and Cancer Cachexia. Cells. 2021; 10(5):1017. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051017
Chicago/Turabian StyleLee, Ji-Hyung, Seon-Wook Kim, Jun-Hyeong Kim, Hyun-Jun Kim, JungIn Um, Da-Woon Jung, and Darren R. Williams. 2021. "Lithium Chloride Protects against Sepsis-Induced Skeletal Muscle Atrophy and Cancer Cachexia" Cells 10, no. 5: 1017. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051017