
High-Yield Mucosal Olfactory Ensheathing Cells Restore Loss of Function in Rat Dorsal Root Injury

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of High-Yield OECs from the Olfactory Mucosa (OM)
2.2. Lentiviral Transduction of ZsGreen Fluorescent Protein
2.3. Encapsulation of Modified Mucosal OECs (mMOECs) in Collagen
2.4. Surgery
2.4.1. Control—Dorsal Rhizotomy and Sham Surgery (n = 14)
2.4.2. Transplantation—Rhizotomy with Transplantation of mMOECs (n = 21)
2.4.3. Biotinylated Dextran Amine (BDA) Labelling (n = 11)
2.4.4. Postoperative Care
2.5. Function Analysis
2.5.1. Vertical Climbing Test
2.5.2. Adhesive Removal Test
2.6. Immunohistochemistry
2.6.1. Cell Culture Evaluation
2.6.2. For In Vivo Study
2.7. Statistical Analysis
3. Results
3.1. Modified Mucosal OECs Culture and Encapsulation in Collagen
3.2. Vertical Climbing Test
3.2.1. Rhizotomy Alone and Sham Surgery (Control Group; n = 14)
3.2.2. Rhizotomy with Transplantation of mMOECs (n = 21)
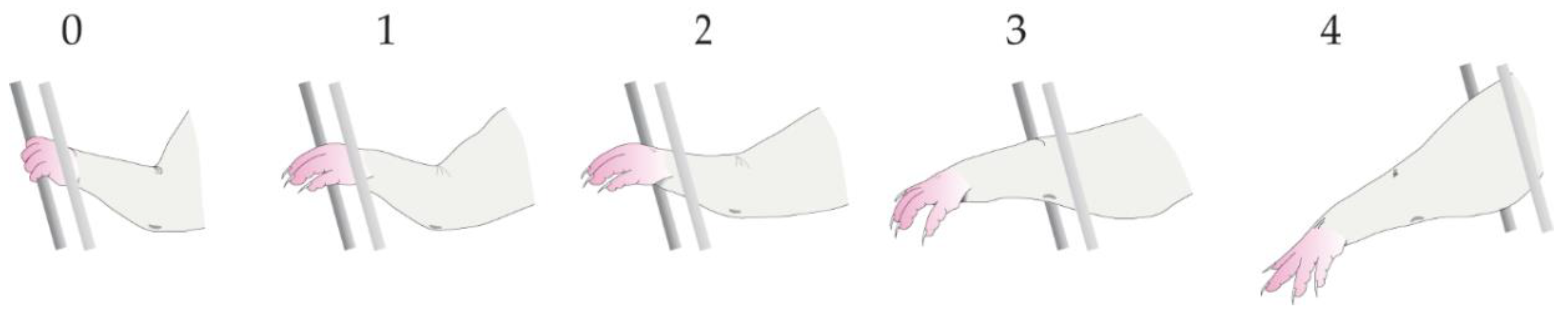
3.3. Forepaw Fault Score
3.3.1. Rhizotomy Alone (Control Group; n = 6)
3.3.2. Rhizotomy with Transplantation of mMOECs (n = 21)
3.4. Adhesive Removal Test (n = 14)
3.5. mMOECs Transplant
3.6. Immunostaining of Neurofilament (NF) and Biotinylated Dextran Amine (BDA) Tracing
3.7. Astrocyte Responses at the Dorsal Root Entry Zone
4. Discussion
4.1. Cell Sources
4.2. Culture Protocols on Mucosal OECs
4.3. Effect of Mucosal OECs Population
4.4. Advantage of Using Biomaterial
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graziadei, P.P.C.; Montigraziadei, G.A. Neurogenesis and neuron regeneration in the olfactory system of mammals. I. Morphological aspects of differentiation and structural organization of the olfactory sensory neurons. J. Neurocytol. 1979, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Schwob, J.E.; Youngentob, S.L.; Ring, G.; Iwema, C.L.; Mezza, R.C. Reinnervation of the rat olfactory bulb after methyl bromide-induced lesion: Timing and extent of reinnervation. J. Comp. Neurol. 1999, 412, 439–457. [Google Scholar] [CrossRef]
- Au, E.; Richter, M.W.; Vincent, A.J.; Tetzlaff, W.; Aebersold, R.; Sage, E.H.; Roskams, A.J. SPARC from olfactory ensheathing cells stimulates Schwann cells to promote neurite outgrowth and enhances spinal cord repair. J. Neurosci. 2007, 27, 7208–7221. [Google Scholar] [CrossRef] [PubMed]
- Ramon-Cueto, A.; Nieto-Sampedro, M. Regeneration into the spinal cord of transected dorsal root axons is promoted by ensheathing glia transplants. Exp. Neurol. 1994, 127, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Franklin, R.J.M.; Barnett, S.C. Olfactory ensheathing cells and CNS regeneration: The sweet smell of success? Neuron 2000, 28, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Field, P.M.; Raisman, G. Repair of adult rat corticospinal tract by transplants of olfactory ensheathing cells. Science 1997, 277, 2000–2002. [Google Scholar] [CrossRef] [PubMed]
- Keyvan-Fouladi, N.; Raisman, G.; Li, Y. Functional repair of the corticospinal tract by delayed transplantation of olfactory ensheathing cells in adult rats. J. Neurosci. 2003, 23, 9428–9434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Ashwell, K. Olfactory ensheathing cells: Their potential use for repairing the injured spinal cord. Spine 2002, 27, 887–892. [Google Scholar] [CrossRef] [PubMed]
- Khankan, R.R.; Griffis, K.G.; Haggerty-Skeans, J.R.; Zhong, H.; Roy, R.R.; Edgerton, V.R.; Phelps, P.E. Olfactory Ensheathing Cell Transplantation after a Complete Spinal Cord Transection Mediates Neuroprotective and Immunomodulatory Mechanisms to Facilitate Regeneration. J. Neurosci. 2016, 36, 6269–6286. [Google Scholar] [CrossRef]
- Wright, A.A.; Todorovic, M.; Tello-Velasquez, J.; Rayfield, A.J.; St John, J.A.; Ekberg, J.A. Enhancing the Therapeutic Potential of Olfactory Ensheathing Cells in Spinal Cord Repair Using Neurotrophins. Cell Transplant. 2018, 27, 867–878. [Google Scholar] [CrossRef] [Green Version]
- Lankford, K.L.; Sasaki, M.; Radtke, C.; Kocsis, J.D. Olfactory ensheathing cells exhibit unique migratory, phagocytic, and myelinating properties in the X-irradiated spinal cord not shared by Schwann cells. Glia 2008, 56, 1664–1678. [Google Scholar] [CrossRef] [PubMed]
- Ramon-Cueto, A.; Cordero, M.I.; Santos-Benito, F.F.; Avila, J. Functional recovery of paraplegic rats and motor axon regeneration in their spinal cords by olfactory ensheathing glia. Neuron 2000, 25, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Steward, O.; Sharp, K.; Selvan, G.; Hadden, A.; Hofstadter, M.; Au, E.; Roskams, J. A re-assessment of the consequences of delayed transplantation of olfactory lamina propria following complete spinal cord transection in rats. Exp. Neurol. 2006, 198, 483–499. [Google Scholar] [CrossRef]
- Toft, A.; Scott, D.T.; Barnett, S.C.; Riddell, J.S. Electrophysiological evidence that olfactory cell transplants improve function after spinal cord injury. Brain 2007, 130, 970–984. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, M.D.; Hsu, D.; Takeoka, A.; Zhong, H.; Ramon-Cueto, A.; Phelps, P.E.; Roy, R.R.; Edgerton, V.R. Further evidence of olfactory ensheathing glia facilitating axonal regeneration after a complete spinal cord transection. Exp. Neurol. 2011, 229, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, A.G.; Kirkwood, P.A.; Raisman, G.; Li, Y. Restoration of hand function in a rat model of repair of brachial plexus injury. Brain 2009, 132, 1268–1276. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Decherchi, P.; Raisman, G. Transplantation of olfactory ensheathing cells into spinal cord lesions restores breathing and climbing. JNSCI 2003, 23, 727–731. [Google Scholar] [CrossRef]
- Doucette, R. Olfactory ensheathing cells: Potential for glial cell transplantation into areas of CNS injury. Histol. Histopathol. 1995, 10, 503–507. [Google Scholar]
- Au, E.; Roskams, A.J. Olfactory ensheathing cells of the lamina propria in vivo and in vitro. Glia 2003, 41, 224–236. [Google Scholar] [CrossRef]
- Barnett, S.C.; Hutchins, A.M.; Noble, M. Purification of olfactory nerve ensheathing cells from the olfactory bulb. Dev. Biol. 1993, 155, 337–350. [Google Scholar] [CrossRef]
- Mackay-Sim, A.; Feron, F.; Cochrane, J.; Bassingthwaighte, L.; Bayliss, C.; Davies, W.; Fronek, P.; Gray, C.; Kerr, G.; Licina, P.; et al. Autologous olfactory ensheathing cell transplantation in human paraplegia: A 3-year clinical trial. Brain 2008, 131, 2376–2386. [Google Scholar] [CrossRef] [Green Version]
- Kawaja, M.D.; Boyd, J.G.; Smithson, L.J.; Jahed, A.; Doucette, R. Technical Strategies to Isolate Olfactory Ensheathing Cells for Intraspinal Implantation. J. Neurotrauma 2009, 26, 155–177. [Google Scholar] [CrossRef]
- Ramer, L.M.; Au, E.; Richter, M.W.; Liu, J.; Tetzlaff, W.; Roskams, A.J. Peripheral olfactory ensheathing cells reduce scar and cavity formation and promote regeneration after spinal cord injury. J. Comp. Neurol. 2004, 473, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Feron, F.; Perry, C.; Cochrane, J.; Licina, P.; Nowitzke, A.; Urquhart, S.; Geraghty, T.; Mackay-Sim, A. Autologous olfactory ensheathing cell transplantation in human spinal cord injury. Brain 2005, 128, 2951–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Feron, F.; Mackay-Sim, A.; Waite, P.M. Olfactory ensheathing cells promote locomotor recovery after delayed transplantation into transected spinal cord. Brain 2002, 125, 14–21. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.; Li, D.Q.; Law, S.; Powell, M.; Raisman, G. A prospective observational study of the yield of olfactory ensheathing cells cultured from biopsies of septal nasal mucosa. Neurosurgery 2008, 62, 1140–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miedzybrodzki, R.; Tabakow, P.; Fortuna, W.; Czapiga, B.; Jarmundowicz, W. The olfactory bulb and olfactory mucosa obtained from human cadaver donors as a source of olfactory ensheathing cells. Glia 2006, 54, 557–565. [Google Scholar] [CrossRef]
- Lima, C.; Escada, P.; Pratas-Vital, J.; Branco, C.; Arcangeli, C.A.; Lazzeri, G.; Maia, C.A.; Capucho, C.; Hasse-Ferreira, A.; Peduzzi, J.D. Olfactory mucosal autografts and rehabilitation for chronic traumatic spinal cord injury. Neurorehabilit. Neural Repair 2010, 24, 10–22. [Google Scholar] [CrossRef]
- Lima, C.; Pratas-Vital, J.; Escada, P.; Hasse-Ferreira, A.; Capucho, C.; Peduzzi, J.D. Olfactory mucosa autografts in human spinal cord injury: A pilot clinical study. J. Spinal Cord Med. 2006, 29, 191–203, discussion 204–196. [Google Scholar] [CrossRef] [Green Version]
- Tabakow, P.; Jarmundowicz, W.; Czapiga, B.; Fortuna, W.; Miedzybrodzki, R.; Czyz, M.; Huber, J.; Szarek, D.; Okurowski, S.; Szewczyk, P.; et al. Transplantation of autologous olfactory ensheathing cells in complete human spinal cord injury. Cell Transplant. 2013, 22, 1591–1612. [Google Scholar] [CrossRef]
- Ibrahim, A.; Li, D.Q.; Collins, A.; Tabakow, P.; Raisman, G.; Li, Y. Comparison of Olfactory Bulbar and Mucosal Cultures in a Rat Rhizotomy Model. Cell Transplant. 2014, 23, 1465–1470. [Google Scholar] [CrossRef] [Green Version]
- Collins, A.; Li, D.; Liadi, M.; Tabakow, P.; Fortuna, W.; Raisman, G.; Li, Y. Partial Recovery of Proprioception in Rats with Dorsal Root Injury after Human Olfactory Bulb Cell Transplantation. J. Neurotrauma 2018, 35, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, M.; Trimaille, T.; Brezun, J.M.; Feron, F.; Gigmes, D.; Marqueste, T.; Decherchi, P. Motor and sensitive recovery after injection of a physically cross-linked PNIPAAm-g-PEG hydrogel in rat hemisectioned spinal cord. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 107, 110354. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, M.; Alluin, O.; Trimaille, T.; Gigmes, D.; Marqueste, T.; Decherchi, P. Delayed Injection of a Physically Cross-Linked PNIPAAm-g-PEG Hydrogel in Rat Contused Spinal Cord Improves Functional Recovery. ACS Omega 2020, 5, 10247–10259. [Google Scholar] [CrossRef]
- Tam, R.Y.; Fuehrmann, T.; Mitrousis, N.; Shoichet, M.S. Regenerative therapies for central nervous system diseases: A biomaterials approach. Neuropsychopharmacology 2014, 39, 169–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.; Ibrahim, A.; Li, D.; Liadi, M.; Li, Y. Reconstruction of the Damaged Dorsal Root Entry Zone by Transplantation of Olfactory Ensheathing Cells. Cell Transplant. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, A.; Li, D.; McMahon, S.B.; Raisman, G.; Li, Y. Transplantation of Cultured Olfactory Bulb Cells Prevents Abnormal Sensory Responses During Recovery From Dorsal Root Avulsion in the Rat. Cell Transplant. 2017, 26, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Barnett, S.C.; Roskams, A.J. Olfactory ensheathing cells. Isolation and culture from the rat olfactory bulb. Methods Mol. Biol. 2002, 198, 41–48. [Google Scholar]
- Ramon-Cueto, A.; Nieto-Sampedro, M. Glial cells from adult rat olfactory bulb: Immunocytochemical properties of pure cultures of ensheathing cells. Neuroscience 1992, 47, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Sauve, Y.; Li, D.Q.; Lund, R.D.; Raisman, G. Transplanted olfactory ensheathing cells promote regeneration of cut adult rat optic nerve axons. JNSCI 2003, 23, 7783–7788. [Google Scholar] [CrossRef] [Green Version]
- Reshamwala, R.; Shah, M.; St John, J.; Ekberg, J. Survival and Integration of Transplanted Olfactory Ensheathing Cells are Crucial for Spinal Cord Injury Repair: Insights from the Last 10 Years of Animal Model Studies. Cell Transplant. 2019, 28, 132S–159S. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, H.; Xi, H.; Zhang, F.; Liu, Y.; Chen, D.; Xiao, J. A prospective randomized double-blind clinical trial using a combination of olfactory ensheathing cells and Schwann cells for the treatment of chronic complete spinal cord injuries. Cell Transplant. 2014, 23 (Suppl. S1), S35–S44. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Xi, H.; Chen, L.; Zhang, F.; Liu, Y. Long-term outcome of olfactory ensheathing cell therapy for patients with complete chronic spinal cord injury. Cell Transplant. 2012, 21 (Suppl. S1), S23–S31. [Google Scholar] [CrossRef]
- Tabakow, P.; Raisman, G.; Fortuna, W.; Czyz, M.; Huber, J.; Li, D.; Szewczyk, P.; Okurowski, S.; Miedzybrodzki, R.; Czapiga, B.; et al. Functional regeneration of supraspinal connections in a patient with transected spinal cord following transplantation of bulbar olfactory ensheathing cells with peripheral nerve bridging. Cell Transplant. 2014, 23, 1631–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Field, P.M.; Raisman, G. Regeneration of adult rat corticospinal axons induced by transplanted olfactory ensheathing cells. J. Neurosci. 1998, 18, 10514–10524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Raisman, G.; Li, D.Q.; Li, Y. Transplanted olfactory mucosal cells restore paw reaching function without regeneration of severed corticospinal tract fibres across the lesion. Brain Res. 2009, 1303, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Franssen, E.H.; Roet, K.C.; De Bree, F.M.; Verhaagen, J. Olfactory ensheathing glia and Schwann cells exhibit a distinct interaction behavior with meningeal cells. J. Neurosci. Res. 2009, 87, 1556–1564. [Google Scholar] [CrossRef]
- Franssen, E.H.; De Bree, F.M.; Essing, A.H.; Ramon-Cueto, A.; Verhaagen, J. Comparative gene expression profiling of olfactory ensheathing glia and Schwann cells indicates distinct tissue repair characteristics of olfactory ensheathing glia. Glia 2008, 56, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.W.; Fletcher, P.A.; Liu, J.; Tetzlaff, W.; Roskams, A.J. Lamina propria and olfactory bulb ensheathing cells exhibit differential integration and migration and promote differential axon sprouting in the lesioned spinal cord 4. JNSCI 2005, 25, 10700–10711. [Google Scholar] [CrossRef]
- Kueh, J.L.L.; Raisman, G.; Li, Y.; Stevens, R.; Li, D.Q. Comparison of Bulbar and Mucosal Olfactory Ensheathing Cells Using FACS and Simultaneous Antigenic Bivariate Cell Cycle Analysis. Glia 2011, 59, 1658–1671. [Google Scholar] [CrossRef]
- Tome, M.; Lindsay, S.L.; Riddell, J.S.; Barnett, S.C. Identification of nonepithelial multipotent cells in the embryonic olfactory mucosa. Stem Cells 2009, 27, 2196–2208. [Google Scholar] [CrossRef] [PubMed]
- Czyz, M.; Tabakow, P.; Gheek, D.; Mis, M.; Jarmundowicz, W.; Raisman, G. The supraorbital keyhole approach via an eyebrow incision applied to obtain the olfactory bulb as a source of olfactory ensheathing cells radiological feasibility study. Br. J. Neurosurg. 2014, 28, 234–240. [Google Scholar] [CrossRef]
- Chhabra, H.S.; Lima, C.; Sachdeva, S.; Mittal, A.; Nigam, V.; Chaturvedi, D.; Arora, M.; Aggarwal, A.; Kapur, R.; Khan, T.A. Autologous olfactory [corrected] mucosal transplant in chronic spinal cord injury: An Indian Pilot Study. Spinal Cord 2009, 47, 887–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granger, N.; Blamires, H.; Franklin, R.J.; Jeffery, N.D. Autologous olfactory mucosal cell transplants in clinical spinal cord injury: A randomized double-blinded trial in a canine translational model. Brain 2012, 135, 3227–3237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddell, J.S.; Enriquez-Denton, M.; Toft, A.; Fairless, R.; Barnett, S.C. Olfactory ensheathing cell grafts have minimal influence on regeneration at the dorsal root entry zone following rhizotomy. Glia 2004, 47, 150–167. [Google Scholar] [CrossRef]
- Ramer, L.M.; Richter, M.W.; Roskams, A.J.; Tetzlaff, W.; Ramer, M.S. Peripherally-derived olfactory ensheathing cells do not promote primary afferent regeneration following dorsal root injury. Glia 2004, 47, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Bianco, J.I.; Perry, C.; Harkin, D.G.; Mackay-Sim, A.; Féron, F. Neurotrophin 3 promotes purification and proliferation of olfactory ensheathing cells from human nose. Glia 2004, 45, 111–123. [Google Scholar] [CrossRef]
- Nash, H.H.; Borke, R.C.; Anders, J.J. New method of purification for establishing primary cultures of ensheathing cells from the adult olfactory bulb. Glia 2001, 34, 81–87. [Google Scholar] [CrossRef]
- Kempski, O.; Wroblewska, B.; Spatz, M. Effects of forskolin on growth and morphology of cultured glial and cerebrovascular endothelial and smooth muscle cells. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 1987, 5, 435–445. [Google Scholar] [CrossRef]
- Monje, P.V. The properties of human Schwann cells: Lessons from in vitro culture and transplantation studies. Glia 2020, 68, 797–810. [Google Scholar] [CrossRef]
- Monje, P.V.; Rendon, S.; Athauda, G.; Bates, M.; Wood, P.M.; Bunge, M.B. Non-antagonistic relationship between mitogenic factors and cAMP in adult Schwann cell re-differentiation. Glia 2009, 57, 947–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, S.; Magu, B.; Rasmussen, C.; Lancaster, S.; Kerksick, C.; Smith, P.; Melton, C.; Cowan, P.; Greenwood, M.; Earnest, C.; et al. Effects of Coleus Forskohlii Supplementation on Body Composition and Hematological Profiles in Mildly Overweight Women. J. Int. Soc. Sports Nutr. 2005, 2, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, H.L.; Astell, K.J.; Mathai, M.L.; Su, X.Q. Coleus forskohlii Extract Supplementation in Conjunction with a Hypocaloric Diet Reduces the Risk Factors of Metabolic Syndrome in Overweight and Obese Subjects: A Randomized Controlled Trial. Nutrients 2015, 7, 9508–9522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massey, J.M.; Amps, J.; Viapiano, M.S.; Matthews, R.T.; Wagoner, M.R.; Whitaker, C.M.; Alilain, W.; Yonkof, A.L.; Khalyfa, A.; Cooper, N.G.; et al. Increased chondroitin sulfate proteoglycan expression in denervated brainstem targets following spinal cord injury creates a barrier to axonal regeneration overcome by chondroitinase ABC and neurotrophin-3. Exp. Neurol. 2008, 209, 426–445. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Carlstedt, T.; Berthold, C.H.; Raisman, G. Interaction of transplanted olfactory-ensheathing cells and host astrocytic processes provides a bridge for axons to regenerate across the dorsal root entry zone. Exp. Neurol. 2004, 188, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, A.; Franklin, R.J.M.; Barnett, S.C. Olfactory ensheathing cells and Schwann cells differ in their in vitro interactions with astrocytes. Glia 2000, 32, 214–225. [Google Scholar] [CrossRef]
- Ahuja, C.S.; Fehlings, M. Concise Review: Bridging the Gap: Novel Neuroregenerative and Neuroprotective Strategies in Spinal Cord Injury. Stem Cells Transl. Med. 2016, 5, 914–924. [Google Scholar] [CrossRef] [Green Version]
- Taras, J.S.; Jacoby, S.M.; Lincoski, C.J. Reconstruction of digital nerves with collagen conduits. J. Hand. Surg. Am. 2011, 36, 1441–1446. [Google Scholar] [CrossRef]
- Khaing, Z.Z.; Schmidt, C.E. Advances in natural biomaterials for nerve tissue repair. Neurosci. Lett. 2012, 519, 103–114. [Google Scholar] [CrossRef]
- Zhong, Y.; Bellamkonda, R.V. Biomaterials for the central nervous system. J. R. Soc. Interface 2008, 5, 957–975. [Google Scholar] [CrossRef]
- Joosten, E.A.; Bar, P.R.; Gispen, W.H. Collagen implants and cortico-spinal axonal growth after mid-thoracic spinal cord lesion in the adult rat. J. Neurosci. Res. 1995, 41, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Orive, G.; Anitua, E.; Pedraz, J.L.; Emerich, D.F. Biomaterials for promoting brain protection, repair and regeneration. Nat. Rev. Neurosci. 2009, 10, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Joosten, E.A.J.; Bär, P.R.; Gispen, W.H. Directional regrowth of lesioned corticospinal tract axons in adult rat spinal cord. Neuroscience 1995, 69, 619–626. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minkelyte, K.; Collins, A.; Liadi, M.; Ibrahim, A.; Li, D.; Li, Y. High-Yield Mucosal Olfactory Ensheathing Cells Restore Loss of Function in Rat Dorsal Root Injury. Cells 2021, 10, 1186. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051186
Minkelyte K, Collins A, Liadi M, Ibrahim A, Li D, Li Y. High-Yield Mucosal Olfactory Ensheathing Cells Restore Loss of Function in Rat Dorsal Root Injury. Cells. 2021; 10(5):1186. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051186
Chicago/Turabian StyleMinkelyte, Kamile, Andrew Collins, Modinat Liadi, Ahmed Ibrahim, Daqing Li, and Ying Li. 2021. "High-Yield Mucosal Olfactory Ensheathing Cells Restore Loss of Function in Rat Dorsal Root Injury" Cells 10, no. 5: 1186. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10051186