
Crosstalk of Multi-Omics Platforms with Plants of Therapeutic Importance

 ,
,  ,
,
Abstract
:1. Introduction
1.1. Medicinal Plants
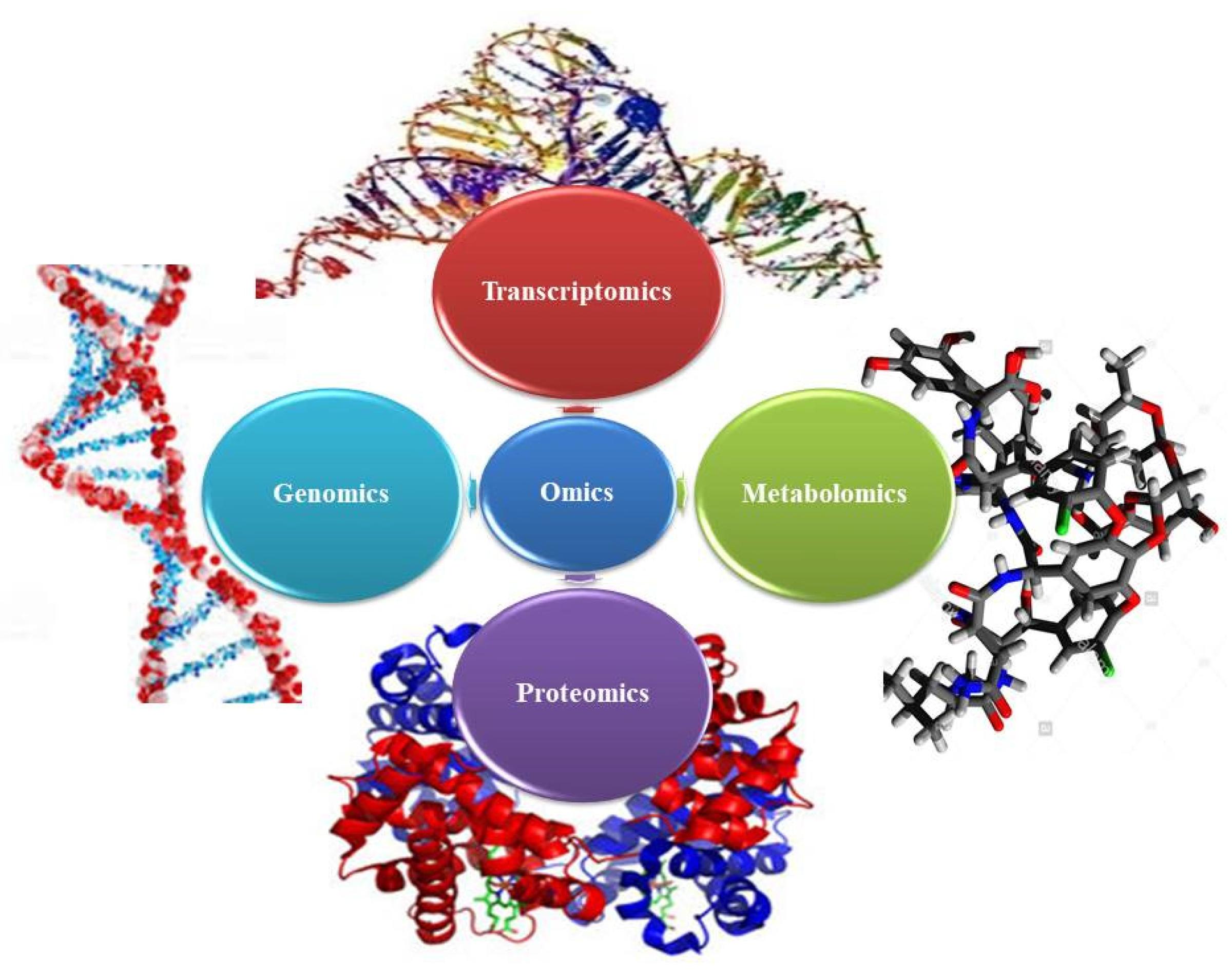
1.2. Omics Technologies
2. Omics in Medicinal Plants
3. Genomics in Medicinal Plants
4. Transcriptomics in Medicinal Plants
5. Proteomics and Functional Analysis of Proteins in Medicinal Plants

{kind=link}
| Medicinal Plants | Reference/s |
|---|---|
| Genomics | |
| Grapes | [56,57] |
| Phalaenopsis equestris (Schauer) Rchb.f | [58] |
| Brassica napus L. | [59] |
| Capsicum annuum L. | [60,61] |
| Momordica charantia L. | [62] |
| Coffea canephora Pierre ex A. Froehner | [63,74,75] |
| Salvia miltiorrhiza Bunge | [64] |
| Ziziphus jujuba Mill. | [65,66] |
| Glycyrrhiza uralensis Fisch. ex DC. | [67] |
| Dendrobium officinale Kimura et Migo | [68] |
| Azadirachta indica A. Juss., 1830 | [69] |
| Catharanthus roseus (L.) G.Don | [45,70,72] |
| Pogostemon cablin (Blanco) Benth. | [71] |
| Solanum tuberosum L. | [76] |
| Panax ginseng | [77] |
| Dendrobium officinale Kimura et Migo | [78] |
| Cypripedium macranthos Sw. (1800) | [78] |
| Ocimum sanctum L. | [79] |
| Ocimum basilicum L. | [80] |
| Papaver somniferum L. | [42,43] |
| Viscum coloratum (Kom.) Nakai | [83] |
| Sinopodophyllum hexandrum (Royle) Ying | [84] |
| Transcriptomics | |
| Podophyllum hexandrum Royle | [95] |
| Cannabis sativa L. | [96,97] |
| Chlorophytum borivilianum Santapau & R.R.Fern. | [98,121] |
| Catharanthus roseus (L.) G. Don | [100,101,113] |
| Curcuma longa L. | [104] |
| Ophiorrhiza pumila Champ. ex Benth. | [94] |
| Withania somnifera (L.) Dunal | [106] |
| Papaver somniferum L. | [44] |
| Salvia miltiorrhiza Burge (Danshen) | [109] |
| Lupinus angustifolius L. | [110] |
| Camptotheca acuminata Decne. | [112] |
| Rauvolfia serpentina (L.) Benth. ex Kurz | [111] |
| Vaccinium macrocarpon Aiton 1789 | [101] |
| Blueberry (Vaccinium sp.) | [115] |
| Rubus coreanus Miq. 1867 | [116] |
| Magnolia sprengeri Pampanini, Nuovo Giorn | [120] |
| Grapefruit | [122] |
| Proteomics | |
| Tripterygium wilfordii Hook.f. | [127] |
| Andrographis paniculata (Burm. F) | [129] |
| Periploca graeca L. | [131] |
| Curcuma longa L. | [132] |
| Coptis chinensis Franch. | [133] |
| Garcinia hanburyi Hook. f. | [134,135,136] |
| Garcinia oblongifolia Champ. ex Benth | [137,138] |
| Salvia miltiorrhiza Burge (Danshen) | [123] |
| Catharanthus roseus (L.) G. Don | [140] |
| Cannabis sativa L. | [141,142] |
| Artemisia annua L. | [143,144] |
| Withania sominifera (L.) Dunal | [146] |
| Metabolomics | |
| Aloe vera (L.) Burm.f. | [151] |
| Panax ginseng C.A. Mey., | [152] |
| Panax notoginseng (Burkill) F.H.Chen | [152] |
| Panax japonicus (T.Nees) C.A. Mey | [152] |
| Persicaria minor (Huds.) Opiz | [153,154] |
| Artemisia | [155] |
| Pulcaria crispa (Forssk.) Benth. ex Oliv. | [155] |
| Rubus coreanus Miq. 1867 | [116] |
6. Metabolomics in Medicinal Plants
7. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Costion, C.; Ford, A.; Cross, H.; Crayn, D.; Harrington, M.; Lowe, A. Plant DNA barcodes can accurately estimate species richness in poorly known floras. PLoS ONE 2011, 6, e26841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, Y.; Wickett, N.J.; Ayyampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; et al. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Song, J.; Sun, C.; Xu, J.; Zhu, Y.; Verpoorte, R.; Fan, T.P. Herbal genomics: Examining the biology of traditional medicines. Science 2015, 347, 527–529. [Google Scholar]
- Pandita, D.; Pandita, A.; Pandita, S. Is “The Doctrine of Signatures” Signature Sequences Designed by God? Int. J. Sci. Res. 2016, 6, 568–572. [Google Scholar]
- Payne, G.F.; Bringi, V.; Prince, C.; Shuler, M. The quest for Commercial Production of Chemicals from Plant Cell Culture. In Plant Cell and Tissue Culture in Liquid Systems; Payne, G.F., Bringi, V., Prince, C., Shuler, M.L., Eds.; Hanser: Munich, Germmany, 1991; pp. 1–10. [Google Scholar]
- Cámara-Leret, R.; Faurby, S.; Macía, M.J.; Balslev, H.; Göldel, B.; Svenning, J.C.; Kissling, W.D.; Rønsted, N.; Saslis-Lagoudakis, C.H. Fundamental species traits explain provisioning services of tropical American palms. Nat. Plants. 2017, 3, 16220. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K.; Sangster, M.; Konijnendijk, C.C. Forests, Trees and Human Health and Well-being: Introduction. In Forests, Trees and Human Health; Nilsson, K., Ed.; Springer: Dordrecht, The Netherlands, 2011. [Google Scholar] [CrossRef]
- Brusotti, G.; Cesari, I.; Dentamaro, A.; Caccialanza, G.; Massolini, G. Isolation and characterization of bioactive compounds from plant resources: The role of analysis in the ethnopharmacological approach. J. Pharm. Biomed. 2014, 87, 218–228. [Google Scholar] [CrossRef]
- Briskin, D.P. Medicinal plants and phytomedicines. Linking plant biochemistry and physiology to human health. Plant Physiol. 2000, 124, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Suárez, A.I.; Chávez, K. Appraisal of medicinal plants with anticancer properties in South America. In Anticancer Plants: Properties and Application; Akhtar, M.S., Swamy, M.K., Eds.; Springer: Singapore, 2018; Volume 1, pp. 229–283. [Google Scholar]
- Busia, K.; Kasilo, O.M. Overview of traditional medicine in Ecowas Member States. Afr. Health Monit. 2010, 13, 16–24. [Google Scholar]
- Nikolovski, B. Arab pharmacy in Macedonia. Bulletin 1961, 1, 20–27. [Google Scholar]
- Katic, R. The Chilandar Medical Codex N. 517; Milincevic, V., Ed.; National Library from Serbia: Belgrade, Serbia, 1980; pp. 9–80. [Google Scholar]
- Katic, R. The Serbian Medicine from 9th to 19th Centuries; Scientific Work: Belgrade, Serbia, 1967; pp. 22–37. [Google Scholar]
- Schmitz, R. Friedrich Wilhelm Sertürner and the discovery of morphine. Pharm. Hist. 1985, 27, 61–74. [Google Scholar]
- Joshi, K.; Ghodke, Y.; Shintre, P. Traditional medicine and genomics. J. Ayurveda Integr. Med. 2010, 1, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modak, M.; Dixit, P.; Londhe, J.; Ghaskadbi, S.; Devasagayam, T.P.A. Indian herbs and herbal drugs used for the treatment of diabetes. J. Clin. Biochem. Nutr. 2007, 40, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Quality Control Methods for Medicinal Plants Materials; WHO: Geneva, Switzerland, 2005; Available online: http://apps.who.int/medicinedocs/pdf/s7918,e/s7918,e.pdf (accessed on 24 December 2015).
- Kaushal, N.; Rao, S.; Ghanghas, P.; Abraham, S.; George, T.; D’Souza, S.; Mathew, J.M.; Chavali, J.; Swamy, M.K.; Baliga, M.S. Usefulness of Ocimum sanctum Linn. in cancer prevention: An update. In Anticancer Plants: Properties and Application; Akhtar, M.S., Swamy, M.K., Eds.; Springer: Singapore, 2018; Volume 1, pp. 415–429. [Google Scholar]
- Farnsworth, N.R. Screening Plants for New Medicines. In Biodiversity; Wilson, E.O., Ed.; National Academy Press: Washington, DC, USA, 1988; pp. 83–97. [Google Scholar]
- Chen, S.L.; Yu, H.; Luo, H.M.; Wu, Q.; Li, C.F.; Steinmetz, A. Conservation and sustainable use of medicinal plants: Problems, progress, and prospects. Chin. Med. 2016, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.V.; Swamy, M.K.; Akhtar, M.S. Anticancer Plants and their Conservation Strategies: An Update. In Anticancer Plants: Properties and Application; Akhtar, M.S., Swamy, M.K., Eds.; Springer: Singapore, 2018; Volume 1, pp. 455–483. [Google Scholar]
- Bandaranayake, W.M. Quality control, screening, toxicity and regulation of herbal drugs. In Modern Phytomedicine: Turning Medicinal Plants Into Drugs; Iqbal, A., Farrukh, A., Eds.; Wiley: New Jersey, NJ, USA, 2006; pp. 25–57. [Google Scholar]
- DeLuca, V.; Salim, V.; Atsumi, S.M.; Yu, F. Mining the biodiversity of plants: A revolution in the making. Science 2012, 336, 1658–1661. [Google Scholar] [CrossRef] [PubMed]
- GIA: Global Industry Analysts (GIA), Inc. Report on the Global Herbal Supplements and Remedies Market Trends Drivers & Projections. Available online: http://www.strategyr.com/marketresearch/herbalsupplementsandremediesmarkettrends.asp (accessed on 15 February 2015).
- Aneesh, T.P.; Hisham, M.; Sekhar, M.S.; Manjusree, M.; Deepa, T.V. International market scenario of traditional Indian herbal drugs in India is declining. Int. J. Green Pharm. 2009, 3, 184–190. [Google Scholar] [CrossRef]
- Prasad, L.V. Indian System of Medicine and Homoeopathy Traditional Medicine in Asia; Chaudhury, R.R., Rafei, U.M., Eds.; WHO-Regional Office for South East Asia: New Delhi, India, 2002; pp. 283–286. [Google Scholar]
- Biswas, K.; Biswas, R. DNA molecular markers based authentication of herbal drugs–A review. IJPRS 2014, 3, 581–593. [Google Scholar]
- Mishra, P.; Kumar, A.; Nagireddy, A.; Mani, D.N.; Shukla, A.K.; Tiwari, R.; Sundaresan, V. DNA barcoding: An efficient tool to overcome authentication challenges in the herbal market. Plant. Biotechnol. 2015, 14, 8–21. [Google Scholar] [CrossRef]
- Pandita, D.; Pandita, A.; Pandita, S. The Revitalizing & Recuperative Higher Tracheophytes of Jammu Province, Jammu and Kashmir (India). Int. J. Indig. Med. Plants 2014, 47, 1603–1620. [Google Scholar]
- Pandita, D.; Pandita, A.; Pandita, S. Herbaceous Medicinal & Therapeutic Plants of District Samba of Jammu Province, Jammu & Kashmir (India). Int. J. Indig. Med. Plants 2013, 46, 1224–1238. [Google Scholar]
- Pandita, D.; Pandita, A.; Pandita, S. Pharmacological and Phytochemical Portrayal of Dicotyledonous Medicinal Plants of Jammu and Kashmir abound with Antidiabetic Potential. J. Microbiol. Biotech. Res. 2015, 5, 16–27. [Google Scholar]
- Pandita, D.; Pandita, A.; Pandita, S. Pharmacological and Phytochemical Portrayal of Monocotyledonous Medicinal Plants of Jammu and Kashmir abound with Antidiabetic Potential. J. Microbiol. Biotech. Res. 2015, 5, 28–33. [Google Scholar]
- Mehta, A.; Hasija, Y. Bioinformatics Approaches for Genomics and Post Genomics Applications of Anticancer Plants. In Anticancer Plants: Mechanisms and Molecular Interactions; Akhtar, M.S., Swamy, M.K., Eds.; Springer: Singapore, 2018; Volume 4, pp. 283–317. [Google Scholar]
- Chakraborty, P. Herbal genomics as tools for dissecting new metabolic pathways of unexplored medicinal plants and drug discovery. Biochim. Open 2018, 6, 9–16. [Google Scholar] [CrossRef]
- Swamy, M.K.; Paramashivaiah, S.; Hiremath, L.; Akhtar, M.S.; Sinniah, U.R. Micropropagation and Conservation of Selected Endangered Anticancer Medicinal Plants from the Western Ghats of India. In Anticancer Plants: Natural Products and Biotechnological Implements; Akhtar, M.S., Swamy, M.K., Eds.; Springer: Singapore, 2018; Volume 2, pp. 481–505. [Google Scholar]
- Swamy, M.K.; Sinniah, U.R.; Ghasemzadeh, A. Anticancer potential of rosmarinic acid and its improved production through biotechnological interventions and functional genomics. Appl. Microbiol. Biotechnol. 2018, 102, 7775–7793. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Fukushima, A.; Saito, K. Transcriptome data modeling for targeted plant metabolic engineering. Curr. Opin. Biotechnol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hale, V.; Keasling, J.D.; Renninger, N.; Diagana, T.T. Microbially derived artemisinin: A biotechnology solution to the global problem of access to affordable antimalarial drugs. Am. J. Trop. Med. Hyg. 2007, 77 (Suppl. 6), 198–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K. Modern plant metabolomics: Advanced natural product gene discoveries, improved technologies, and future prospects. Nat. Prod. Rep. 2015, 32, 212–229. [Google Scholar] [CrossRef]
- Suzuki, M.; Nakabayashi, R.; Ogata, Y.; Sakurai, N.; Tokimatsu, T.; Goto, S.; Suzuki, M.; Jasinski, M.; Martinoia, E.; Otagaki, S.; et al. Multiomics in grape berry skin revealed specific induction of the stilbene synthetic pathway by ultraviolet-C irradiation. Plant Physiol. 2015, 168, 47–59. [Google Scholar] [CrossRef]
- Hagel, J.M.; Facchini, P.J. Dioxygenases catalyze the Odemethylation steps of morphine biosynthesis in opium poppy. Nat. Chem. Biol. 2010, 6, 273–275. [Google Scholar] [CrossRef]
- Gesell, A.; Rolf, M.; Ziegler, J.; Diaz Chavez, M.L.; Huang, F.C.; Kutchan, T.M. CYP719B1 is salutaridine synthase, the C-C phenol-coupling enzyme of morphine biosynthesis in opium poppy. J. Biol. Chem. 2009, 284, 24432–24442. [Google Scholar] [CrossRef] [Green Version]
- Winzer, T.; Gazda, V.; He, Z.; Kaminski, F.; Kern, M.; Larson, T.R.; Li, Y.; Meade, F.; Teodor, R.; Vaistij, F.E.; et al. A Papaver somniferum 10- gene cluster for synthesis of the anticancer alkaloid noscapine. Science 2012, 336, 1704–1708. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.T. Phytochemical Omics in Medicinal Plants. Biomolecules 2020, 10, 936. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Meng, Y.; Wang, P.; Tang, Z.; Wang, H.; Xie, T. Bioinformatics-assisted, integrated omics studies on medicinal plants. Brief. Bioinform. 2020, 21, 1857–1874. [Google Scholar] [CrossRef] [PubMed]
- Khoomrung, S.; Wanichthanarak, K.; Nookaew, I.; Thamsermsang, O.; Seubnooch, P.; Laohapand, T.; Akarasereenont, P. Metabolomics and Integrative Omics for the Development of Thai Traditional Medicine. Front. Pharmacol. 2017, 8, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, J.; Sen, P.; Choudhury, M.D.; Dehury, B.; Barooah, M.; Modi, M.K.; Talukdar, A.D. Rediscovering medicinal plants’ potential with OMICS: Microsatellite survey in expressed sequence tags of eleven traditional plants with potent antidiabetic properties. Omics J. Integr. Biol. 2014, 18, 298–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellner, A. Genome Sequence of Catharanthus Roseus. 2015. Available online: http://medicinalplantgenomics.msu.edu/ (accessed on 25 December 2019).
- Kellner, F.; Kim, J.; Clavijo, B.J.; Hamilton, J.P.; Childs, K.L.; Vaillancourt, B.; Cepela, J.; Habermann, M.; Steuernagel, B.; Clissold, L.; et al. Genome-guided investigation of plant natural product biosynthesis. Plant J. 2015, 82, 680–692. [Google Scholar] [CrossRef]
- Transcriptome Characterization, Sequencing, and Assembly of Medicinal Plants Relevant to Human Health. 2011. Available online: https://apps.pharmacy.uic.edu/depts/pcrps/MedTranscriptomePlants/database.html (accessed on 20 December 2019).
- One Thousand Plant Transcriptomes Initiative. One thousand plant transcriptomes and the phylogenomics of green plants. Nature 2019, 574, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.Y.; Wegel, E.; Osbourn, A. From hormones to secondary metabolism: The emergence of metabolic gene clusters in plants. Plant J. 2011, 66, 66–79. [Google Scholar] [CrossRef]
- Wurtele, E.; Chappell, J.; Jones, A.; Celiz, M.; Ransom, N.; Hur, M.; Rizshsky, L.; Crispin, M.; Dixon, P.; Liu, J.; et al. Medicinal plants: A public resource for metabolomics and hypothesis development. Metabolites 2012, 2, 1031–1059. [Google Scholar] [CrossRef]
- Gahlaut, A.; Vikas, D.M.; Gothwal, A.; Kulharia, M.; Chhillar, A.K.; Hooda, V.; Dabur, R. Proteomics and metabolomics: Mapping biochemical regulations. Drug Invent. Today 2013, 5, 321–326. [Google Scholar] [CrossRef]
- Seki, H.; Sawai, S.; Ohyama, K.; Mizutani, M.; Ohnishi, T.; Sudo, H.; Fukushima, E.O.; Akashi, T.; Aoki, T.; Saito, K.; et al. Triterpene functional genomics in licorice for identification of CYP72A154 involved in the biosynthesis of glycyrrhizin. Plant Cell 2011, 23, 4112–4123. [Google Scholar] [CrossRef] [Green Version]
- Hao, D.C.; Xiao, P.G. Genomics and evolution in traditional medicinal plants: Road to a healthier life. EBO 2015, 11, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Boutanaev, A.M.; Moses, T.; Zi, J.; Nelson, D.R.; Mugford, S.T.; Peters, R.J.; Osbourn, A. Investigation of terpene diversification across multiple sequenced plant genomes. Proc. Natl. Acad. Sci. USA 2015, 112, E81–E88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Sun, Y.; Xu, J.; Luo, H.; Sun, C.; He, L.; Cheng, X.; Zhang, B.; Xiao, P. Strategies of the study on herb genome program. Acta Pharm. Sin. (Yao Xue Xue Bao) 2010, 45, 807–812. [Google Scholar]
- Velasco, R.; Zharkikh, A.; Troggio, M.; Cartwright, D.A.; Cestaro, A.; Pruss, D.; Pindo, M.; Fitzgerald, L.M.; Vezzulli, S.; Reid, J.; et al. A high quality draft consensus sequence of the genome of a heterozygous grapevine variety. PLoS ONE 2007, 2, e132. [Google Scholar] [CrossRef] [Green Version]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. French-Italian Public Consortium for Grapevine Genome C: The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar]
- Cai, J.; Liu, X.; Vanneste, K.; Proost, S.; Tsai, W.C.; Liu, K.W.; Chen, L.J.; He, Y.; Xu, Q.; Bian, C.; et al. The genome sequence of the orchid Phalaenopsis equestris. Nat. Genet. 2015, 47, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Park, M.; Yeom, S.I.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef]
- Urasaki, N.; Takagi, H.; Natsume, S.; Uemura, A.; Taniai, N.; Miyagi, N.; Fukushima, M.; Suzuki, S.; Tarora, K.; Tamaki, M.; et al. Draft genome sequence of bitter gourd (Momordica charantia), a vegetable and me-dicinal plant in tropical and subtropical regions. DNA Res. 2017, 24, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Denoeud, F.; Carretero-Paulet, L.; Dereeper, A.; Droc, G.; Guyot, R.; Pietrella, M.; Zheng, C.; Alberti, A.; Anthony, F.; Aprea, G.; et al. The coffee genome provides insight into the convergent evolution of caffeine biosynthesis. Science 2014, 345, 1181–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, J.; Song, J.; Gao, H.; Zhu, Y.; Xu, J.; Pang, X.; Yao, H.; Sun, C.; Li, X.; Li, C. The complete chloroplast genome sequences of the medicinal plant Salvia miltiorrhiza. PLoS ONE 2013, 8, e57607. [Google Scholar] [CrossRef]
- Mahajan, R.T.C.M. Phyto-pharmacology of Ziziphus jujuba Mill-a plant review. Pharmacol. Rev. 2009, 3, 320–329. [Google Scholar]
- Li, Y.; Xu, C.; Lin, X. De novo assembly and characterization of the fruit transcriptome of Chinese Jujuba (Zizipus jujuba Mill) using 454 pyrosequencing and the development of novel trinucleotide SSR markers. PLoS ONE 2014, 9, e106438. [Google Scholar] [CrossRef]
- Mochida, K.; Sakurai, T.; Seki, H.; Yoshida, T.; Takahagi, K.; Sawai, S.; Uchiyama, H.; Muranaka, T.; Saito, K. Draft genome assembly and annotation of Glycyrrhiza uralensis, a medicinal legume. Plant J. 2017, 89, 181–194. [Google Scholar] [CrossRef]
- Yan, L.; Wang, X.; Liu, H.; Tian, Y.; Lian, J.; Yang, R.; Hao, S.; Wang, X.; Yang, S. The genome of Dendrobium officinale illu-minates the biology of the important traditional Chinese orchid herb. Mol. Plant 2015, 8, 922–934. [Google Scholar] [CrossRef] [Green Version]
- Shivaraj, Y.; Govind, S.; Jogaiah, S.; Sannaningaiah, D. Functional analysis of medicinal plants using system biology approaches. Int. J. Pharm. Pharmaceut. Sci. 2015, 7, 41–43. [Google Scholar]
- Ku, C.; Chung, W.C.; Chen, L.L.; Kuo, C.H. The Complete Plastid Genome Sequence of Madagascar Periwinkle Catharanthus roseus (L.) G. Don: Plastid Genome Evolution, Molecular Marker Identification, and Phylogenetic Implications in Asterids. PLoS ONE 2013, 8, e68518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Xiao, H.; Deng, C.; Xiong, L.; Yang, J.; Peng, C. The complete chloroplast genome sequences of the medicinal plant Pogostemon cablin. Int. J. Mol. Sci. 2016, 17, 820–830. [Google Scholar] [CrossRef] [Green Version]
- Giddings, L.A.; Liscombe, D.K.; Hamilton, J.P.; Childs, K.L.; DellaPenna, D.; Buell, C.R.; O’Connor, S.E. A stereo selective hydroxylation step of alkaloid biosynthesis by a unique cytochrome P450 in Catharanthus roseus. J. Biol. Chem. 2011, 286, 16751–16757. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, N.M.; Pattnaik, S.; Jain, P.; Gaur, P.; Choudhary, R.; Vaidyanathan, S.; Deepak, S.; Hariharan, A.K.; Krishna, P.G.B.; Nair, J. A draft of the genome and four transcriptomes of a medicinal and pesticidal angiosperm Azadirachta indica. BMC Genom. 2012, 13, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Yuan, L.; Wu, B.; Li, X.; Chen, S.; Lu, S. Genome-wide identification and characterization of novel genes involved in terpenoid biosynthesis in Salvia miltiorrhiza. J. Exp. Bot. 2012, 63, 2809–2823. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Song, J.; Luo, H.; Zhang, Y.; Li, Q.; Zhu, Y.; Xu, J.; Li, Y.; Song, C.; Wang, B.; et al. Analysis 26 of the Genome Sequence of the Medicinal Plant Salvia miltiorrhiza. Mol. Plant 2016, 9, 949–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridlender, M.; Kapulnik, Y.; Koltai, H. Plant derived substances with anticancer activity: From folklore to practice. Front. Plant Sci. 2015, 6, 799. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yin, J.; Guo, H.; Zhang, Y.; Xiao, W.; Sun, C.; Wu, J.; Qu, X.; Yu, J.; Wang, X.; et al. The complete chloroplast genome provides insight into the evolution and polymorphism of Panax ginseng. Front. Plant Sci. 2015, 5, 696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Hou, B.W.; Niu, Z.T.; Liu, W.; Xue, Q.Y.; Ding, X.Y. Comparative chloroplast genomes of photosynthetic orchids: Insights into evolution of the Orchidaceae and development of molecular markers for phylogenetic applications. PLoS ONE 2014, 9, e99016. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Kalra, A.; Gupta, V. Unravelling the genome of Holy basil an incomparable“elixir of life” of traditional Indian medicine. BMC Genom. 2015, 16, 413–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, S.; Meena, S.; Bhattacharya, A. De novo sequencing and comparative analysis of holy and sweet basil transcrip-tomes. BMC Genom. 2014, 15, 588–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, A.K.; Chacko, A.R.; Gandhimathi, A.; Ghosh, P.; Harini, K.; Joseph, A.P.; Joshi, A.G.; Karpe, S.D.; Kaushik, S.; Ku-ravadi, N.; et al. Genome sequencing of herb Tulsi (Ocimum tenuiflorum) unravels key genes behind its strong me-dicinal properties. BMC Plant Biol. 2015, 15, 212. [Google Scholar] [CrossRef] [Green Version]
- Rubin, B.E.; Ree, R.H.; Moreau, C.S. Inferring phylogenies from RAD sequence data. PLoS ONE 2012, 7, e33394. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.Y.; Park, H.S.; Kim, S.; Kim, Y.D. Development of microsatellite markers for Viscum coloratum (Santalaceae) and their application to wild populations. Appl. Plant Sci. 2017, 5, 1600102. [Google Scholar] [CrossRef]
- Liu, W.; Yin, D.; Liu, J.; Li, N. Genetic diversity and structure of Sinopodophyl¬lum hexandrum (Royle) Ying in the Qinling Mountains, China. PLoS ONE 2014, 9, e110500. [Google Scholar]
- Jiao, Y.; Paterson, A.H. Polyploidy-associated genome modifications during land plant evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollister, J.D.; Greiner, S.; Wang, W.; Wang, J.; Zhang, Y.; Wong, G.K.; Wright, S.I.; Johnson, M.T. Recurrent loss of sex is associated with accumulation of deleterious mutations in Oenothera. Mol. Biol. Evol. 2015, 32, 896–905. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; McKain, M.R.; Harkess, A.; Nelson, M.N.; Dash, S.; Deyholos, M.K.; Peng, Y.; Joyce, B.; Stewart, C.N., Jr.; Rolf, M. Multiple polyploidy events in the early radiation of nodulating and nonnodulating legumes. Mol. Biol. Evol. 2015, 32, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Moore, M.J.; Brockington, S.F.; Soltis, D.E.; Wong, G.K.; Carpenter, E.J.; Zhang, Y.; Chen, L.; Yan, Z.; Xie, Y. Dissecting molecular evolution in the highly diverse plant clade Caryo-phyllales using transcriptome sequencing. Mol. Biol. Evol. 2015, 32, 2001–2014. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.C.; Xiao, P.G.; Peng, Y.; Dong, J.; Liu, W. Evaluation of the chloroplast barcoding markers by mean and smallest interspecific distances. Pak. J. Bot. 2012, 44, 1271–1274. [Google Scholar]
- Zhang, F.; Gao, Q.; Khan, G.; Luo, K.; Chen, S. Comparative transcriptome analysis of aboveground and underground tissues of Rhodiola algida, an important ethno-medicinal herb endemic to the Qinghai-Tibetan Plateau. Gene 2014, 553, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.C.; Ge, G.; Xiao, P.; Zhang, Y.; Yang, L. The first insight into the tissue specific taxus transcriptome via Illumina second generation sequencing. PLoS ONE 2011, 6, e21220. [Google Scholar]
- Hao, D.C.; Chen, S.L.; Osbourn, A.; Kontogianni, V.G.; Liu, L.W.; Jordán, M.J. Temporal transcriptome changes induced by methyl jasmonate in Salvia sclarea. Gene 2015, 558, 41–53. [Google Scholar] [CrossRef]
- Hao, D.C.; Ma, P.; Mu, J.; Chen, S.; Xiao, P.; Peng, Y.; Huo, L.; Xu, L.; Sun, C. De novo characterization of the root transcriptome of a traditional Chinese medicinal plant Polygonum cuspidatum. Sci. China Life Sci. 2012, 55, 452–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamazaki, M.; Mochida, K.; Asano, T.; Nakabayashi, R.; Chiba, M.; Udomson, N.; Yamazaki, Y.; Goodenowe, D.B.; Sankawa, U.; Yoshida, T.; et al. Coupling deep transcriptome analysis with untargeted metabolic profiling in Ophiorrhiza pumila to further the understanding of the biosynthesis of the anti-cancer alkaloid camptothecin and anthraquinones. Plant. Cell Physiol. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, W.; Sattely, E.S. Six enzymes from mayapple that complete the biosynthetic pathway to the etoposide aglycone. Science 2015, 349, 1224–1228. [Google Scholar] [CrossRef] [Green Version]
- Gagne, S.J.; Stout, J.M.; Liu, E.; Boubakir, Z.; Clark, S.M.; Page, J.E. Identification of olivetolic acid cyclase from Cannabis sativa reveals a unique catalytic route to plant polyketides. Proc. Natl. Acad. Sci. USA 2012, 109, 1281–1286. [Google Scholar] [CrossRef] [Green Version]
- Stout, J.M.; Boubakir, Z.; Ambrose, S.J.; Purves, R.W.; Page, J.E. The hexanoyl-CoA precursor for cannabinoid biosynthesis is formed by an acyl-activating enzyme in Cannabis sativa trichomes. Plant J. 2012, 71, 353–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Meena, P.; Verma, S.; Kumar, M.; Kumar, A. Anti-tumour, anti-mutagenic and chemomodulatory potential of Chlorophytum borivilianum. A Pac. J. Cancer Prev. 2010, 11, 327–334. [Google Scholar]
- Hao, D.C.; He, C.N.; Shen, J.; Xiao, P.G. Anticancer chemodiversity of Ranunculaceae medicinal plants: Molecular mecha-nisms and functions. Curr. Genom. 2017, 18, 39–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinda, B.; Chowdhury, R.; Mohanta, D. Naturally occurring iridoids, secoiridoids and their bioactivity. Chem. Pharm. Bull. 2009, 57, 765–796. [Google Scholar] [CrossRef] [Green Version]
- Geu-Flores, F.; Sherden, N.H.; Courdavault, V.; Burlat, V.; Glenn, W.S.; Wu, C.; Nims, E.; Cui, Y.; O’Connor, S.E. An alternative route to cyclic terpenes by reductive cyclization in iridoid biosynthesis. Nature 2012, 492, 138–142. [Google Scholar] [CrossRef]
- Dinda, B.; Debnath, S.; Banik, R. Naturally occurring iridoids and secoiridoids. Chem. Pharm. Bull. 2011, 59, 803–833. [Google Scholar] [CrossRef] [Green Version]
- Tundis, R.; Loizzo, M.R.; Menichini, F. Biological and pharmacological activities of iridoids: Recent developments. Med. Chem. 2008, 8, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Annadurai, R.S.; Neethiraj, R.; Jayakumar, V.; Damodaran, A.C.; Rao, S.N.; Katta, M.A.; Gopinathan, S.; Sarma, S.P.; Senthilkumar, V.; Niranjan, V.; et al. De novo transcriptome assembly (NGS) of Curcuma longa L. rhizome reveals novel transcripts related to anticancer and antimalarial terpenoids. PLoS ONE 2013, 8, e56217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, D.C.; Xiao, P.G.; Liu, L.W.; Peng, Y.; He, C.N. Essentials of pharmacophylogeny: Knowledge pedigree, episte-mology and paradigm shift. China J. Chin. Mat. Med. 2015, 40, 1–8. [Google Scholar]
- Gupta, P.; Goel, R.; Agarwal, A.K.; Asif, M.H.; Sangwan, N.S.; Sangwan, R.S.; Trivedi, P.K. Comparative tran-scriptome analysis of different chemotypes elucidates withanolide biosynthesis pathway from medicinal plant Withania somnifera. Sci. Rep. 2015, 5, 1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, T.; Kobayashi, K.; Kashihara, E.; Sudo, H.; Sasaki, R.; Iijima, Y.; Aoki, K.; Shibata, D.; Saito, K.; Yamazaki, M. Suppression of camptothecin biosynthetic genes results in metabolic modification of secondary products in hairy roots of Ophiorrhiza pumila. Phytochemistry 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, G.; Huang, L.; Tang, X.; Zhao, J. Candidate genes involved in tanshinone biosynthesis in hairy roots of Salvia miltiorrhiza revealed by cDNA microarray. Mol. Biol. Rep. 2011, 38, 2471–2478. [Google Scholar] [CrossRef]
- Wenping, H.; Yuan, Z.; Jie, S.; Lijun, Z.; Zhezhi, W. De novo transcriptome sequencing in Salvia miltiorrhiza to identify genes involved in the biosynthesis of active ingredients. Genomics 2011, 98, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Bunsupa, S.; Katayama, K.; Ikeura, E.; Oikawa, A.; Toyooka, K.; Saito, K.; Yamazaki, M. Lysine decarboxylase catalyzes the first step of quinolizidine alkaloid biosynthesis and coevolved with alkaloid production in Leguminosae. Plant Cell 2012, 24, 1202–1216. [Google Scholar] [CrossRef] [Green Version]
- Facchini, P.J.; Bohlmann, J.; Covello, P.S.; De Luca, V.; Mahadevan, R.; Page, J.E.; Ro, D.K.; Sensen, C.W.; Storms, R.; Martin, V.J. Synthetic biosystems for the production of high-value plant metabolites. Trends Biotechnol. 2012, 30, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Salim, V.; Yu, F.; Altarejos, J.; De Luca, V. Virus-induced gene silencing identifies Catharanthus roseus 7-deoxyloganic acid-7-hydroxylase, a step in iridoid and monoterpene indole alkaloid biosynthesis. Plant J. 2013, 76, 754–765. [Google Scholar] [CrossRef]
- Gongora-Castillo, E.; McKnight, T.D.; O’Connor, S.; Childs, K.L.; Buell, C.; Fedewa, G.; Hamilton, J.P.; Liscombe, D.K.; Magallanes-Lundback, M.; Mandadi, K.K.; et al. Development of transcriptomic resources for interrogating the biosynthesis of monoterpene indole alkaloids in medicinal plant species. PLoS ONE 2012, 7, e52506. [Google Scholar] [CrossRef]
- Stavrinides, A.; Tatsis, E.C.; Foureau, E.; Caputi, L.; Kellner, F.; Courdavault, V.; O’Connor, S.E. Unlocking the diversity of alkaloids in Catharanthus roseus: Nuclear localization suggests metabolic channeling in secondary metabolism. Chem. Biol. 2015, 22, 336–341. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Liu, Y.; Gai, Y.; Geng, J.; Chen, L.; Liu, H.; Kang, L.; Tian, Y.; Li, Y. De novo sequencing and analysis of the cranberry fruit transcriptome to identify putative genes involved in flavonoid biosynthesis, transport and regulation. BMC Genom. 2015, 16, 652–669. [Google Scholar] [CrossRef] [Green Version]
- Hyun, T.K.; Lee, S.; Rim, Y.; Kumar, R.; Han, X.; Lee, S.Y.; Lee, C.H.; Kim, J.Y. De-novo RNA sequencing and metabolite profiling to identify genes involved in anthocyanin biosynthesis in Korean black raspberry (Rubus coreanus Miquel). PLoS ONE 2014, 9, e88292. [Google Scholar] [CrossRef] [Green Version]
- Bovy, A.; Schijlen, E.; Hall, R.D. Metabolic engineering of flavonoids in tomato: The potential for metabolomics. Metabolomics 2007, 3, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Hong, L.; Qian, Q.; Tang, D.; Wang, K.; Li, M.; Cheng, Z. A mutation in the rice chalcone isomerase gene causes the golden hull and internode 1 phenotype. Planta 2012, 236, 141–151. [Google Scholar] [CrossRef]
- Muir, S.R.; Collins, G.J.; Robinson, S.; Hughes, S.; Bovy, A.; Ric De Vos, C.H.; van Tunen, A.J.; Verhoeyen, M.E. Overexpression of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001, 19, 470–474. [Google Scholar] [CrossRef]
- Shi, S.G.; Yang, M.; Zhang, M.; Wang, P.; Kang, Y.X.; Liu, J.J. Genome-wide transcriptome analysis of genes involved in flavonoid biosynthesis between red and white strains of Magnolia sprengeri pamp. BMC Genom. 2014, 15, 706–776. [Google Scholar] [CrossRef] [Green Version]
- Kaushik, N. Saponins of Chlorophytum species. Phytochem. Rev. 2005, 4, 191–196. [Google Scholar] [CrossRef]
- Grimplet, J.; Cramer, G.R.; Dickerson, J.A.; Mathiason, K.; Van Hemert, J.; Fennell, A.Y. VitisNet. “Omics” integration through grapevine molecular networks. PLoS ONE 2009, 4, e8365. [Google Scholar] [CrossRef] [Green Version]
- Lao, Y.; Wang, X.; Xu, N.; Zhang, H.; Xu, H. Application of proteomics to determine the mechanism of action of traditional Chinese medicine remedies. J. Ethnopharmacol. 2014, 155, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Sun, H.; Yuan, Y.; Sun, W.; Jiao, G.; Wang, X. An in vivo analysis of the therapeutic and synergistic properties of Chinese medicinal formula Yin-Chen-Hao-Tang based on its active constituents. Fitoterapia 2011, 82, 1160–1168. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, R.Y.; He, Q.Y. Proteomic analysis of anticancer TCMs targeted at mitochondria. Evid. Based Complement. Altern. Med. 2015, 2015, 539260. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.S.; Kuo, Y.H.; Chin, C.C.; Wang, J.Y.; Yu, H.R.; Sheen, J.M.; Tung, S.Y.; Shen, C.H.; Chen, T.C.; Sung, M.L. Proteomic analysis of the effects of baicalein on colorectal cancer cells. Proteomics 2012, 12, 810–819. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, L.; Zhou, G.B. The main anticancer bullets of the Chinese medicinal herb, thunder God vine. Molecules 2011, 16, 5283–5297. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Song, F.; Wu, W.K.; He, M.; Zhao, L.; Sun, X.; Li, H.; Jiang, Y.; Yang, Y.; Peng, K. Triptolide inhibits colon cancer cell proliferation and induces cleavage and translocation of 14–3-3 epsilon. Cell Biochem. Funct. 2012, 30, 271–278. [Google Scholar] [CrossRef]
- Valdiani, A.; Kadir, M.A.; Tan, S.G.; Talei, D.; Abdullah, M.P.; Nikzad, S. Nain-e Havandi (Andrographis paniculata) present yesterday, absent today: A plenary review on underutilized herb of Iran’s pharmaceutical plants. Mol. Biol. Rep. 2012, 39, 5409–5424. [Google Scholar] [CrossRef]
- Talei, D.; Valdiani, A.; Rafii, M.Y.; Maziah, M. Proteomic analysis of the salt-responsive leaf and root proteins in the anticancer plant Andrographis paniculata Nees. PLoS ONE 2014, 91, e112907. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Song, Q.; Yang, J.; Zhao, X.; Zhang, X.; Yang, P.; Kang, J. Comparative proteomic analysis of anti-cancer mechanism by periplocin treatment in lung cancer cells. Cell Physiol. Biochem. 2014, 33, 859–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, H.Y.; Chen, S.B.; Guo, D.J.; Pan, S.Y.; Yu, Z.L. Proteomic identification of differentially expressed proteins in curcumin-treated MCF-7 cells. Phytomedicine 2011, 18, 697–703. [Google Scholar] [CrossRef]
- Wang, N.; Wang, X.; Tan, H.Y.; Li, S.; Tsang, C.M.; Tsao, S.W.; Feng, Y. Berberine suppresses cyclin D1 expression through proteasomal degradation in human hepatoma cells. Int. J. Mol. Sci. 2016, 17, 1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chantarasriwong, O.; Batova, A.; Chavasiri, W.; Theodorakis, E.A. Chemistry and biology of the caged Garcinia xanthones. Chem. Eur. J. 2010, 16, 9944–9962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Zhang, R.H.; He, S.C.; Xu, Q.Y.; Ma, L.; Wang, G.C.; Qiu, N.; Peng, F.; Chen, J.Y.; Qiu, J.X.; et al. Synthesis and antiangiogenic activity of novel gambogic acid derivatives. Molecules 2012, 17, 6249–6268. [Google Scholar] [CrossRef]
- Anantachoke, N.; Tuchinda, P.; Kuhakarn, C.; Pohmakotr, M.; Reutrakul, V. Prenylated caged xanthones: Chemistry and biology. Pharm. Biol. 2012, 50, 78–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, W.M.; Zhang, J.F.; Wang, H.; Tan, H.S.; Wang, W.M.; Chen, S.C.; Zhu, X.; Chan, T.M.; Tse, C.M.; Leung, K.S.; et al. Apoptosis induced by 1,3,6,7-tetrahydroxy xanthone in Hepatocellular carcinoma and proteomic analysis. Apoptosis 2012, 12, 842–851. [Google Scholar] [CrossRef]
- Fu, W.M.; Zhang, J.F.; Wang, H.; Xi, Z.C.; Wang, W.M.; Zhuang, P.; Zhu, X.; Chen, S.C.; Chan, T.M.; Leung, K.S.; et al. Heat shock protein 27 mediates the effect of 1,3,5-trihydroxy-13,13-dimethyl-2H- pyran [7,6-b] xanthone on mitochondrial apoptosis in hepatocellular carcinoma. J. Proteome. 2012, 75, 4833–4843. [Google Scholar] [CrossRef]
- Alsemari, A.; Alkhodairy, F.; Aldakan, A.; Al-Mohanna, M.; Bahoush, E.; Shinwari, Z.; Alaiya, A. The selective cytotoxic anticancer properties and proteomic analysis of Trigonella foenum-graecum. BMC Complement. Altern. Med. 2014, 14, 114. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, D.I.; Gaspari, M.; van der Greef, J.; van der Heijden, R.; Verpoorte, R. Proteome analysis of the medicinal plant Catharanthus roseus. Planta 2005, 221, 690–704. [Google Scholar] [CrossRef]
- Raharjo, T.J.; Widjaja, I.; Roytrakul, S.; Verpoorte, R. Comparative proteomics of cannabis sativa plant tissues. J. Biomol. Tech. 2004, 15, 97–106. [Google Scholar]
- Jenkins, C.; Orsburn, B. The Cannabis Proteome Draft Map Project. Int. J. Mol. Sci. 2020, 21, 965. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Yan, T.; Ji, L.; Dong, Y.; Sidoli, S.; Yuan, Z.; Cai, C.; Chen, J.; Tang, Y.; Shen, Q.; et al. Comprehensive Map of the Artemisia annua Proteome and Quantification of Differential Protein Expression in Chemotypes Producing High versus Low Content of Artemisinin. Proteomics 2020, 20, e1900310. [Google Scholar] [CrossRef] [PubMed]
- Bryant, L.; Flatley, B.; Patole, C.; Brown, G.D.; Cramer, R. Proteomic analysis of Artemisia annua–Towards elucidating the biosynthetic pathways of the antimalarial pro-drug artemisinin. BMC Plant Biol. 2015, 15, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.W.; Lee, S.H.; Min, C.W.; Jo, I.H.; Bang, K.H.; Hyun, D.Y.; Agrawal, G.K.; Rakwal, R.; Zargar, S.M.; Gupta, R. Ginseng (Panax sp.) proteomics: An update. Appl. Biol. Chem. 2017, 60, 311–320. [Google Scholar] [CrossRef]
- Nagappan, A.; Karunanithi, N.; Sentrayaperumal, S.; Park, K.I.; Park, H.S.; Lee, D.H.; Kang, S.R.; Kim, J.A.; Senthil, K.; Natesan, S.; et al. Comparative root protein profiles of Korean ginseng (Panax ginseng) and Indian ginseng (Withania somnifera). Am. J. Chin. Med. 2012, 40, 203–218. [Google Scholar] [CrossRef]
- Hartmann, T. From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef]
- Hao, D.C.; Xiao, P.G.; Liu, M.; Peng, Y.; He, C.N. Pharmaphylogeny vs. pharmacophylogenomics: Molecular phylogeny, evolution and drug discovery. Yao Xue Xue Bao 2014, 49, 1387–1394. [Google Scholar]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural Products (Secondary Metabolites). Biochem. Mol. Biol. Plants 2000, 24, 1250–1319. [Google Scholar]
- Taiz, L.; Zeiger, E. Secondary Metabolites and Plant Defense. In Plant Physiology, 5th ed.; Sinauer Associates Inc., Publishers: Sunderland, MA, USA, 2006; pp. 369–400. [Google Scholar]
- Baharum, S.N.; Bunawan, H.; Ghani, M.A.; Wan Aida Wan, M.; Noor, N.M. Analysis of the chemical composition of the essential oil of Polygonum minus Huds. Using two-dimensional gas chromatography-time-of-flight mass spectrometry (GC-TOF MS). Molecules 2010, 15, 7006–7015. [Google Scholar] [CrossRef] [Green Version]
- Saito, K.; Matsuda, F. Metabolomics for functional genomics, systems biology, and biotechnology. Annu. Rev. Plant Biol. 2010, 61, 463–489. [Google Scholar] [CrossRef]
- Ulrich-Merzenich, G.; Zeitler, H.; Jobst, D.; Panek, D.; Vetter, H.; Wagner, H. Application of the ‘omic’ technologies in phytomedicine. Phytomedicine 2007, 14, 70–82. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Harwansh, R.K.; Bahadur, S.; Biswas, S.; Kuchibhatla, L.N.; Tetali, S.D.; Raghavendra, A.S. Metabo-lomics of medicinal plants- a versatile tool for standardization of herbal products and quality evaluation of Ayurvedic formulations. Curr. Sci. 2016, 111, 1624–1630. [Google Scholar] [CrossRef]
- Kim, H.K.; Wilson, E.G.; Choi, Y.H.; Verpoorte, R. Metabolomics: A tool for anticancer lead-finding from natural products. Planta Med. 2010, 76, 1094–1102. [Google Scholar] [CrossRef] [Green Version]
- Okada, T.; Mochamad Afendi, F.; Altaf-Ul-Amin, M.; Takahashi, H.; Nakamura, K.; Kanaya, S. Metabolomics of me-dicinal plants: The importance of multivariate analysis of analytical chemistry data. Curr. Comput. Aided Drug Des. 2010, 6, 179–196. [Google Scholar] [CrossRef]
- Field, B.; Fiston-Lavier, A.S.; Kemen, A.; Geisler, K.; Quesneville, H.; Osbourn, A.E. Formation of plant metabolic gene clusters within dynamic chromosomal regions. Proc. Natl. Acad. Sci. USA 2011, 108, 16116–16121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, R.A.; Strack, D. Phytochemistry meets genome analysis, and beyond. Phytochemistry 2003, 62, 815–816. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Matsuda, F.; Kasai, K.; Fukuoka, S.; Kitamura, K.; Tozawa, Y.; Miyagawa, H.; Wakasa, K. Mutation of a rice gene encoding a phenylalanine biosynthetic enzyme results in accumulation of phenylalanine and tryptophan. Plant Cell 2008, 20, 1316–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noorolahi, S.M.; Sadeghi, S.; Mohammadi, M.; Azadi, M.; Rahimi, N.A.; Vahabi, F.; Arjmand, M.; Hosseini, H.; Mosallatpur, S.; Zamani, Z. Metabolomic profiling of cancer cells to Aloe vera extract by 1HNMR spectroscopy. J. Metabol. 2016, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Xie, G.; Plumb, R.; Su, M.; Xu, Z.; Zhao, A.; Qiu, M.; Long, X.; Liu, Z.; Jia, W. Ultra-performance LC/TOF MS analysis of medicinal Panax herbs for metabolomic research. J. Sep. Sci. 2008, 31, 1015–1026. [Google Scholar] [CrossRef]
- Goh, H.H.; Khairudin, K.; Sukiran, N.A.; Normah, M.N.; Baharum, S.N. Metabolite profiling reveals temperature effects on the VOCs and flavonoids of different plant populations. Plant Biol. 2016, 18, 130–139. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta. 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981, to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- El-Naggar, S.A.; Abdel-Farid, I.B.; Elgebaly, H.A.; Germoush, M.O. Metabolomic profiling, antioxidant capacity and in vitro anticancer activity of some compositae plants growing in Saudi Arabia. Afr. J. Pharm. Pharmacol. 2015, 9, 764–774. [Google Scholar]
- Marraffa, J.M. Amiodarone. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 197–199. [Google Scholar] [CrossRef]
- Meyer, U. From khellin to sodium cromoglycate—A tribute to the work of Dr. R. E. C. Altounyan (1922–1987). Die Pharm. 2002, 57, 62–69. [Google Scholar]
- Gregson, K.S.; Bennett, J.D. Drugs Acting on the Respiratory System. In Pharmacology and Therapeutics for Dentistry, 7th ed.; Dowd, F.J., Johnson, B., Mariotti, A., Eds.; Mosby: Maryland Heights, MO, USA, 2017; pp. 392–403. [Google Scholar] [CrossRef]
- Jarukamjorn, K.; Nemoto, N. Pharmacological aspects of Andrographis paniculata on health and its major diterpenoid constituent andrographolide. J. Health Sci. 2008, 54, 370–381. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, K.; Zhang, T.; Sun, Y.; Chen, W.; Li, Q.; Chen, R.; Sun, X. The Neuroprotective Effect and Probable Mechanism of DL3-n-Butylphthalide in Brain Diseases. Integr. Med. Int. 2014, 1, 51–55. [Google Scholar] [CrossRef]
- Aderibigbe, B.A. Design of Drug Delivery Systems Containing Artemisinin and Its Derivatives. Molecules 2017, 22, 323. [Google Scholar] [CrossRef] [Green Version]
- Tallarida, R.J. Donnatal® (Robins). In TOP 200; Springer: New York, NY, USA, 1982. [Google Scholar] [CrossRef]
- Sekhar, V.C.; Viswanathan, G.; Baby, S. Insights Into the Molecular Aspects of Neuroprotective Bacoside A and Bacopaside, I. Curr. Neuropharmacol. 2019, 17, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Jiang, T.; Li, Q.; Ling, X. Camptothecin (CPT) and its derivatives are known to target topoisomerase I (Top1) as their mechanism of action: Did we miss something in CPT analogue molecular targets for treating human disease such as cancer? Am. J. Cancer Res. 2017, 7, 2350–2394. [Google Scholar]
- De Petrocellis, L.; Ligresti, A.; Moriello, A.S.; Allarà, M.; Bisogno, T.; Petrosino, S. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br. J. Pharmacol. 2011, 163, 1479–1494. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Bley, K. Topical capsaicin for pain management: Therapeutic potential and mechanisms of action of the new high-concentration capsaicin 8% patch. Br. J. Anaesth. 2011, 107, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azra, A.A.; Babak, D.H.; Hassan, E.; Ahmad, M. High in vitro production of anti-canceric indole alkaloids from periwinkle (Catharanthus roseus) tissue culture. Afr. J. Biotechnol. 2008, 7, 2834–2839. [Google Scholar]
- Günter, G.; Desiree, A.; Brigitte, W.; Hanns, H. Effects of Quinine, Quinidine and Chloroquine on Human Muscle Nicotinic Acetylcholine Receptors. Front. Pharm. 2018, 9. [Google Scholar] [CrossRef]
- Dubey, K.K.; Ray, A.R.; Behera, B.K. Production of demethylated colchicine through microbial transformation and scale-up process development. Process Biochem. 2008, 43, 251–257. [Google Scholar] [CrossRef]
- Leung, Y.Y.; Yao Hui, L.L.; Kraus, V.B. Colchicine—Update on mechanisms of action and therapeutic uses. Semin. Arthritis Rheum. 2015, 45, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Luthra, P.M.; Singh, R.; Chandra, R. Therapeutic uses of Curcuma longa (turmeric). Indian J. Clin. Biochem. 2001, 16, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollman, A. Drugs for atrial fibrillation. Digoxin comes from Digitalis lanata. BMJ 1996, 312, 912. [Google Scholar] [CrossRef]
- The American Society of Health-System Pharmacists. Digoxin. Available online: https://www.drugs.com/monograph/digoxin.html (accessed on 8 December 2016).
- Chandrasekara, A.; Kumar, T.J. Roots and tuber crops as functional foods: A review on phytochemical constituents and their potential health benefits. Int. J. Food Sci. 2016, 3631, 647. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, H.A.; Bhalsing, S.R. Plant derived Novel Biomedicinal: Diosgenin. Int. J. Pharmacogn. Phytochem. Res. 2014, 6, 780–784. [Google Scholar]
- Biscoping, J.; Bachmann-Mennenga, M.B. Lokalanästhetika: Vom Ester zum Isomer [Local anesthetics from ester to isomer]. Anasthesiol. Intensivmed. Notfallmed. Schmerzther. 2000, 35, 285–292. (In German) [Google Scholar] [CrossRef]
- Behloul, N.; Wu, G. Genistein: A promising therapeutic agent for obesity and diabetes treatment. Eur. J. Pharmacol. 2013, 698, 31–38. [Google Scholar] [CrossRef]
- Hartmann-Boyce, J.; Chepkin, S.C.; Ye, W.; Bullen, C.; Lancaster, T. Nicotine replacement therapy versus control for smoking cessation. Cochrane Database Syst. Rev. 2018, 5, CD000146. [Google Scholar] [CrossRef]
- Barut, G.A.; Tunç, M.; Şahin, Ş.; Ulus, F.; Sazak, H. Effects of epidural morphine and levobupivacaine combination before incision and after incision and in the postoperative period on thoracotomy pain and stress response. Turk. J. Med. Sci. 2018, 48, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Rida, P.C.; LiVecche, D.; Ogden, A.; Zhou, J.; Aneja, R. The Noscapine Chronicle: A Pharmaco-Historic Biography of the Opiate Alkaloid Family and its Clinical Applications. Med. Res. Rev. 2015, 35, 1072–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soni, D.; Grover, A. “Picrosides” from Picrorhiza kurroa as potential anti-carcinogenic agents. Biomed. Pharm. 2019, 109, 1680–1687. [Google Scholar] [CrossRef] [PubMed]
- Rajeshkumar, N.V.; Kuttan, R. Inhibition of N-nitrosodiethylamine-induced hepatocarcinogenesis by Picroliv. J. Exp. Clin. Cancer Res. 2000, 19, 459–465. [Google Scholar]
- Ansari, R.A.; Tripathi, S.C.; Patnaik, G.K.; Dhawan, B.N. Antihepatotoxic properties of picroliv: An active fraction from rhizomes of Picrorhiza kurrooa. J. Ethnopharmacol. 1991, 34, 61–68. [Google Scholar] [CrossRef]
- Ardalani, H.; Avan, A.; Ghayour-Mobarhan, M. Podophyllotoxin: A novel potential natural anticancer agent. Avicenna J. Phytomedicine 2017, 7, 285–294. [Google Scholar] [PubMed]
- Lobay, D. Rauwolfia in the Treatment of Hypertension. Integr. Med. 2015, 14, 40–46. [Google Scholar]
- Li, Z.M.; Xu, S.W.; Liu, P.Q. Salvia miltiorrhiza Burge (Danshen): A golden herbal medicine in cardiovascular therapeutics. Acta Pharmacol. Sin. 2018, 39, 802–824. [Google Scholar] [CrossRef] [Green Version]
- Weaver, B.A. How Taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef]

| Medicinal Plants | Medicinal Metabolites | Drugs/Synthetic Derivatives | Medicinal Properties | Structures | Reference/s |
|---|---|---|---|---|---|
| Ammi visnaga | Visnadin, visnagin, and khellin | Amiodarone for cardiac dysrhythmias; Cromolyn for treatment of asthma | Kidney stones, menstrual cramps to atherosclerosis. Cardiac arrhythmias, congestive heart failure, angina, and hypercholesterolemia |  Khellin Khellin
| [170,171,172] |
| Andrographis paniculata (Burm.f.) Nees | Andrographolide | Anti-inflammatory, antibacterial, antitumor, antidiabetic, antimalarial and hepatoprotective |  | [173] | |
| Apium graveolens L. | L-3-n-butylphthalide drug | Promising candidate for treatment of cerebral ischemia, Parkinson’s and Alzheimer’s disease |  | [174] | |
| Artemisia annua L. | Artemisinin | Artemether, artemether and artesunate | Antimalarial and antiviral activity and resist infections from protozoans |  | [175] |
| Atropa belladonna, Datura stramonium | Atropine/Hyoscyamine/Scopolamine | Donnatal | peripheral anticholinergic or antispasmodic action |  | [176] |
| Bacopa monniera L. | Bacoside A3, bacopaside 1, bacopaside 2, jujubogenin, bacosaponine C | Neuromedicine for various disorders such as anxiety, depression and memory loss |  | [177] | |
| Camptotheca acuminata Descne and Nothapodytes nimmoniana (J. Grah.) D.J. Mabberley | Camptothecin | Topotecan and irinotecan | Anticancer |  | [178] |
| Cannabis sativa L. | Cannabinoids | Anti-inflammatory, anticancer, analgesic, muscle relaxant, neuro-antioxidative and psychoactive drugs |  | [179] | |
| Capsicum sp. | Capsaicine | Apsaicin 8% patch (Qutenza™) | Topical analgesic, Neuropeptide-releasing agent selective for primary sensory peripheral neurons |  | [180] |
| Catharanthus roseus L. | Vinblastine and vincristine | Anticancer drugs |  Vincristine Vincristine | [181] | |
 Vinblastine Vinblastine | |||||
| Cinchona officinalis L. | Natural quinine, quinidine | Chloroquine | Antimalaria |  | [182] |
| Colchicum autumnale L. | Colchicine | Colchicine analogs namely, 3-demethyl colchicine, colchicoside, thiocolchicocide | Anticancer |  | [183] |
| Anti-gout and familial Mediterranean fever (FMF), pericarditis, coronary artery disease and other inflammatory and fibrotic conditions | [184] | ||||
| Curcuma longa L. | Curcuminoids mainly curcumin | Antioxidant, neuroprotective, antitumor, anti-inflammatory, anti-acidogenic, radioprotective and anti-arthritis |  | [185] | |
| Digitalislanata | Digoxin/digitoxin | Digitalis (digoxin) sold under brand name Lanoxin and digitoxin by brand name Crystodigin | Heart medicine |  Digoxin Digoxin | [186,187] |
 Digitoxin Digitoxin | |||||
| Dioscorea species | Diosgenin steroidal sapogenin | Sex hormones, corticosteroids, oral contraceptives and steroidal drugs |  | [188,189] | |
| Erytrhoxylum coca Lam. | Cocaine | Topical anesthesia of the eye |  | [190] | |
| Glycina max (L.) Merr | Genistein | Antiosteoporosis, anti-inflammatory, anticancer, antioxidant, antidiabetic, and antiobesity activities |  | [191] | |
| Nicotiana tabacum L. | Nicotine | Smoking cessation drug to relieve withdrawal symptoms |  | [192] | |
| Papaver somniferum L. | Morphine | Musculoskeletal pain, abdominal pain, chest pain, arthritis, and even headaches |  | [193] | |
| Papaver somniferum L. | Noscapine | Opiate analgesics, antitussive, stroke, anticancer |  Noscapine Noscapine | [194] | |
| Codeine | Opiate analgesics, antitussive |  Codeine Codeine | |||
| Picrorhiza kurroa Royle ex Benth. | Picroside | Picrosides as anticarcinogenic agents, hepatoprotective drug formulation, picroliv |  | [195,196,197] | |
| Podophyllum hexandrum Royle (syn. P. emodi Wall.) and Podophyllum peltatum L. | Podophyllotoxins | Etoposide and teniposide | Anticancer drugs |  | [198] |
| Rauwolfia serpentine L. | Reserpine | Hypertension |  | [199] | |
| Salvia miltiorrhiza Burge (Danshen) | Tanshinoate B, Danshensu, Isotanshinone ⅡA and Cryptotanshinone | Small-molecule cardiovascular drug discovery |  Danshensu Danshensu | [200] | |
| Taxum brevifolia Nutt. | Taxol | Paclitaxel |  | [201] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandita, D.; Pandita, A.; Wani, S.H.; Abdelmohsen, S.A.M.; Alyousef, H.A.; Abdelbacki, A.M.M.; Al-Yafrasi, M.A.; Al-Mana, F.A.; Elansary, H.O. Crosstalk of Multi-Omics Platforms with Plants of Therapeutic Importance. Cells 2021, 10, 1296. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061296
Pandita D, Pandita A, Wani SH, Abdelmohsen SAM, Alyousef HA, Abdelbacki AMM, Al-Yafrasi MA, Al-Mana FA, Elansary HO. Crosstalk of Multi-Omics Platforms with Plants of Therapeutic Importance. Cells. 2021; 10(6):1296. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061296
Chicago/Turabian StylePandita, Deepu, Anu Pandita, Shabir Hussain Wani, Shaimaa A. M. Abdelmohsen, Haifa A. Alyousef, Ashraf M. M. Abdelbacki, Mohamed A. Al-Yafrasi, Fahed A. Al-Mana, and Hosam O. Elansary. 2021. "Crosstalk of Multi-Omics Platforms with Plants of Therapeutic Importance" Cells 10, no. 6: 1296. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061296