
Direct Interaction of Polar Scaffolding Protein Wag31 with Nucleoid-Associated Protein Rv3852 Regulates Its Polar Localization

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Media
2.2. Co-Immunoprecipitation of Rv3852 Interacting Proteins from Mtb and M. smegmatis
2.3. Generation of M. smegmatis and Mtb Strains
2.4. Purification of Proteins
2.5. In Vitro Protein–Protein Interaction
2.6. Growth Curves
2.7. Sub-Cellular Fractionation
2.8. Immunofluorescence
2.9. BODIPY FL Vancomycin Staining
3. Results
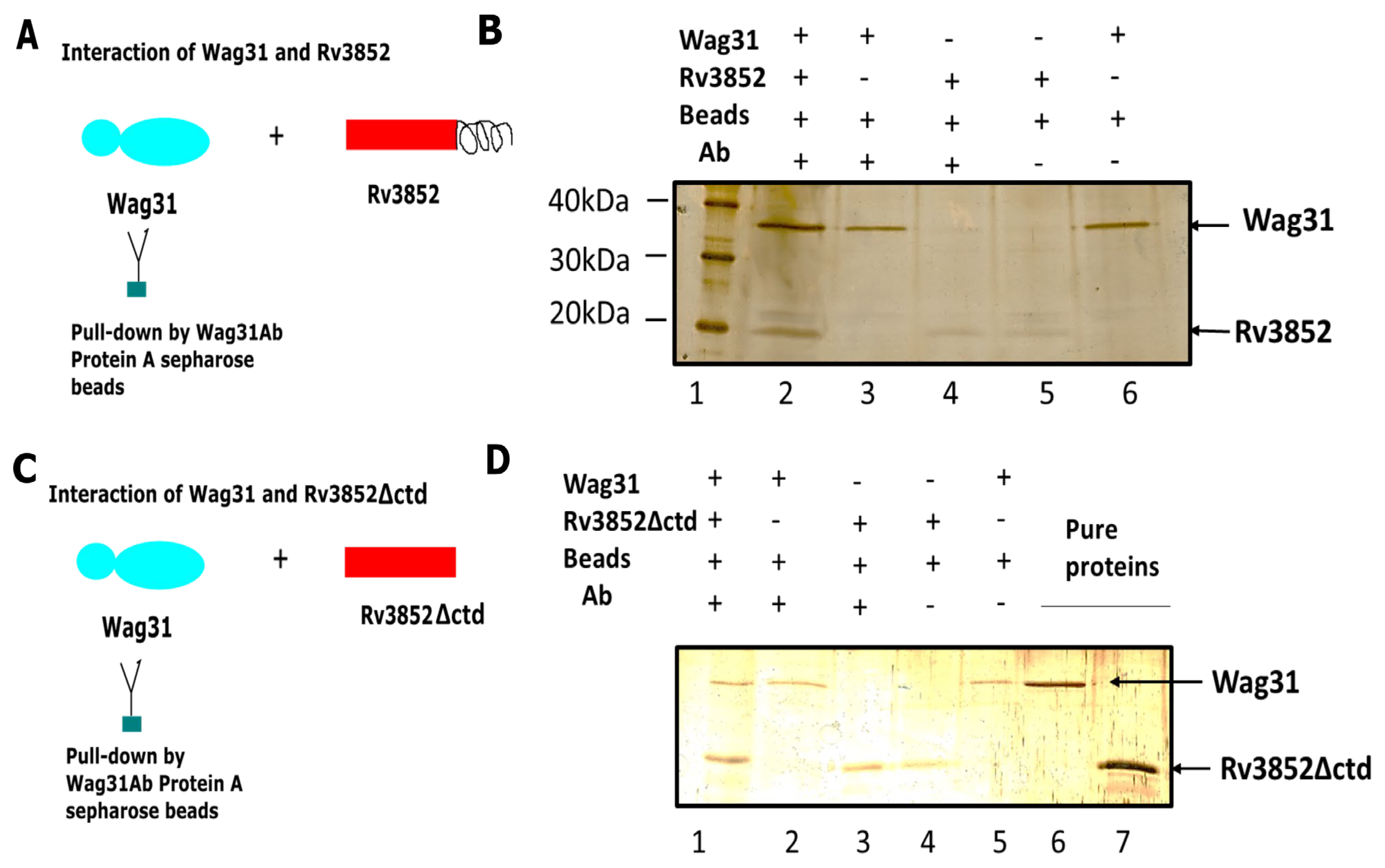
3.1. Rv3852 Interacts with Wag31 in Mtb
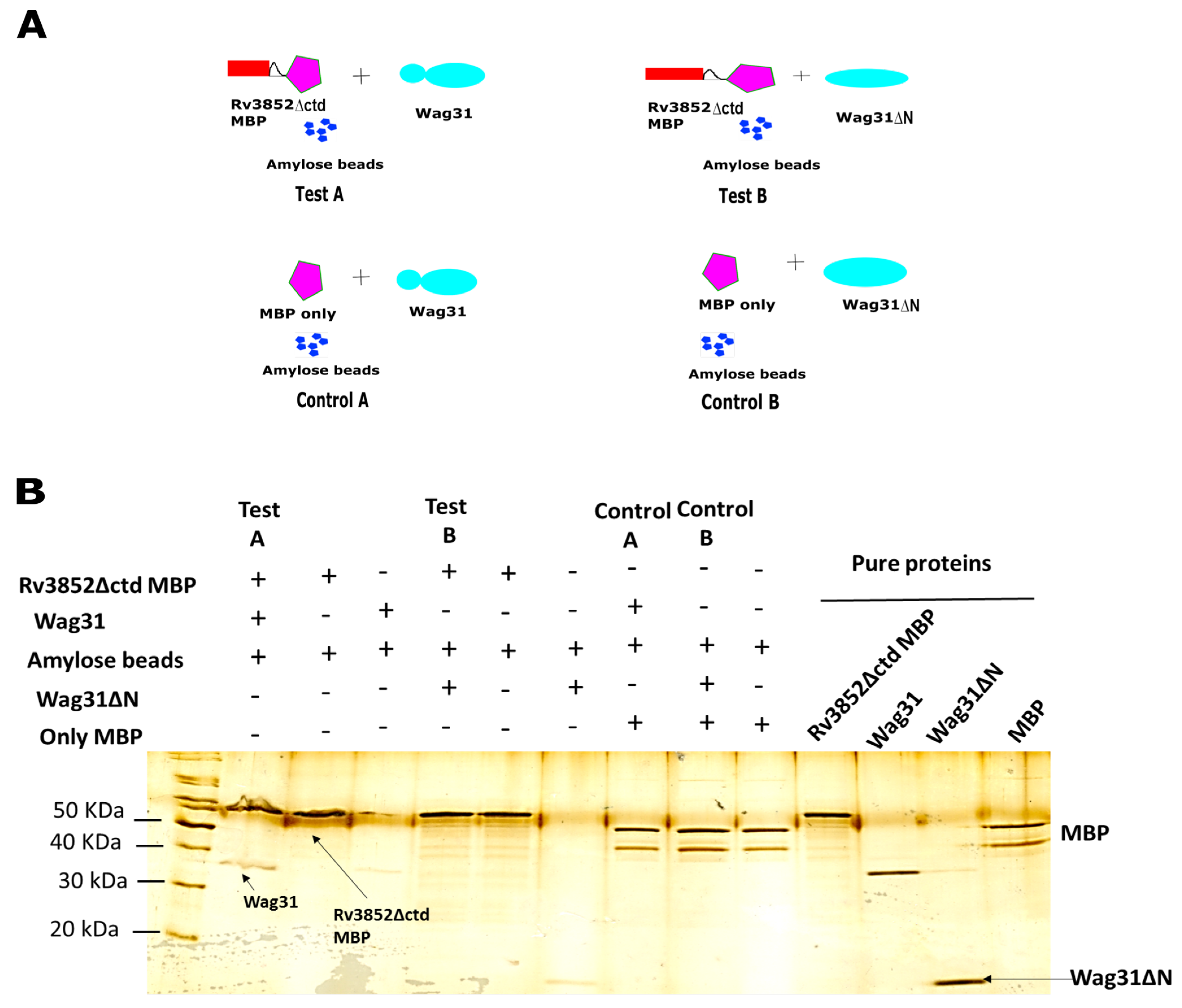
3.2. Direct Physical Interaction between Rv3852 and Wag31
3.3. Rv3852 Interacts with N-terminal Region of Wag31
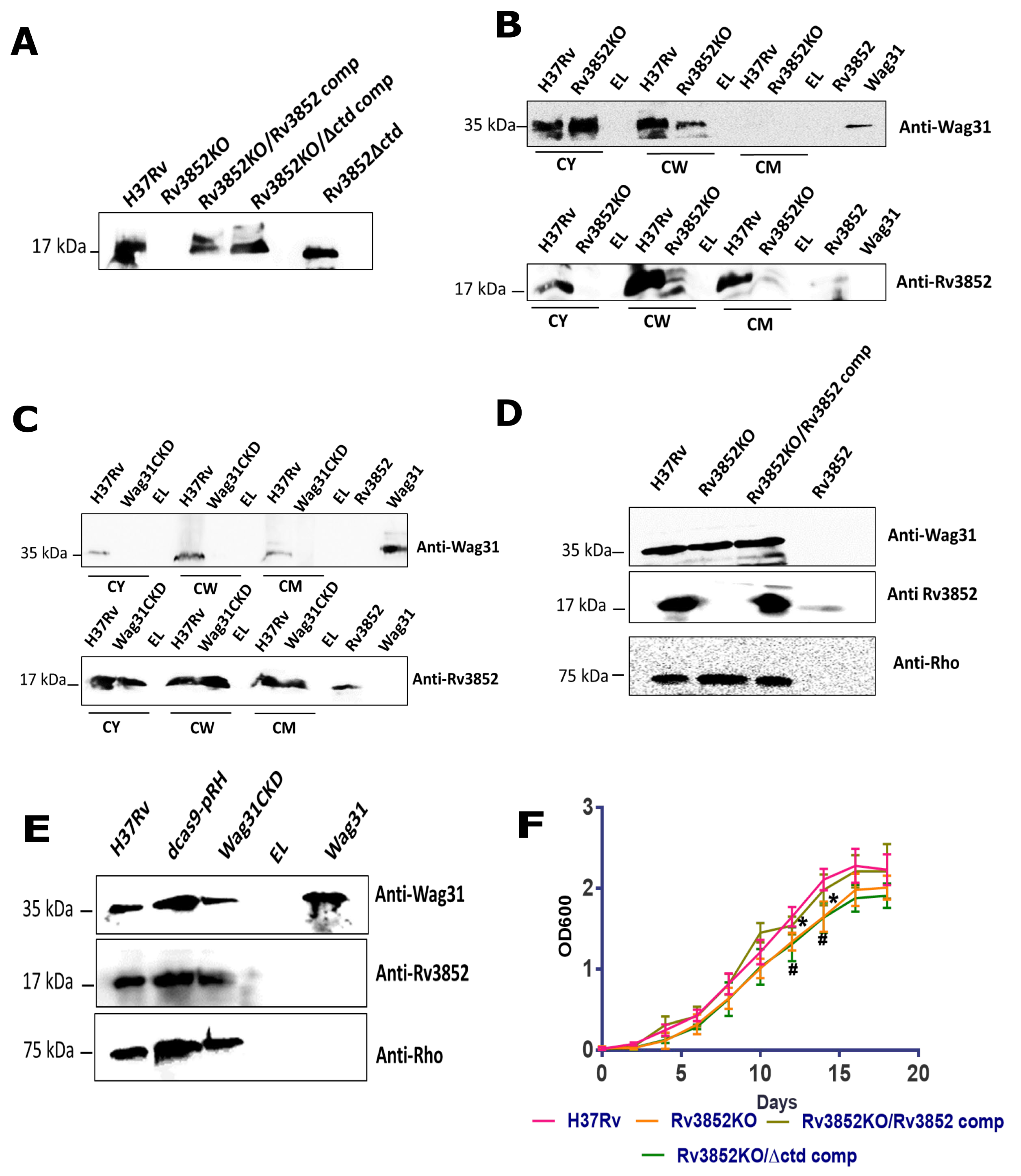
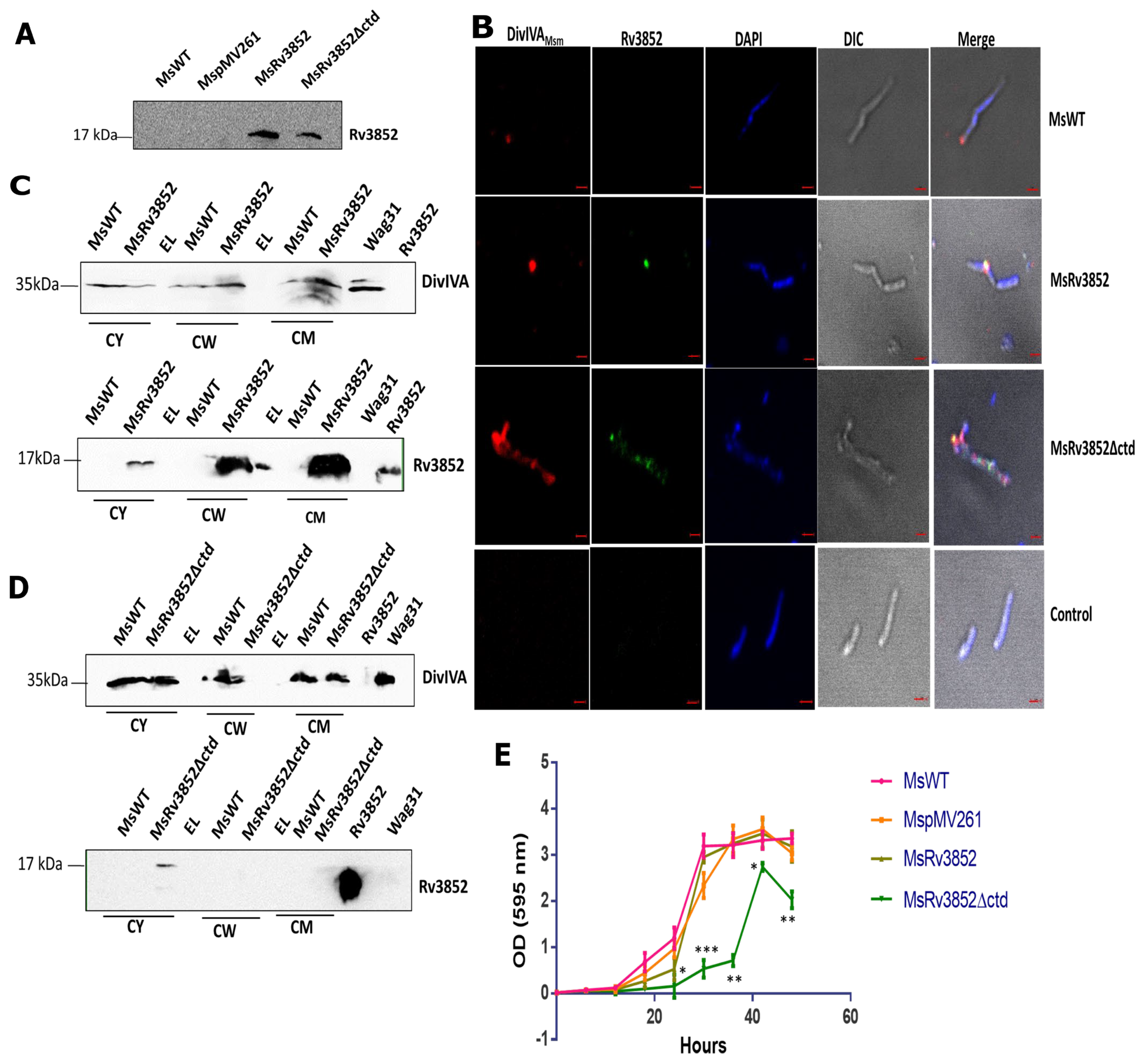
3.4. Rv3852 Influences Localization of Wag31 in the Mycobacterial Cell Wall
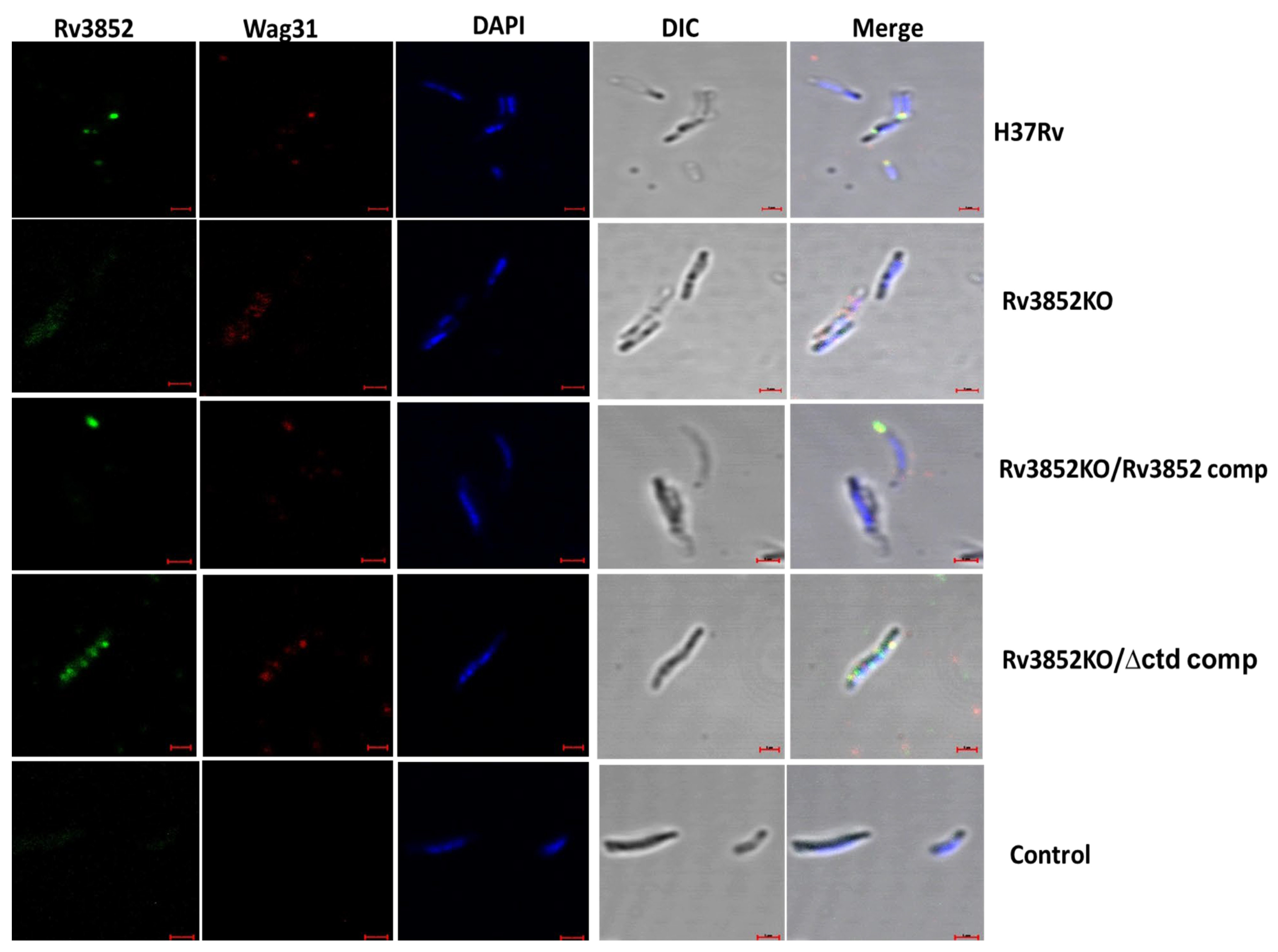
3.5. Absence of Rv3852 Disturbs Polar Localization of Wag31
3.6. Ectopic Expression of Rv3852 in M. smegmatis Leads to More DivIVAMsm Deposition in Cell Wall
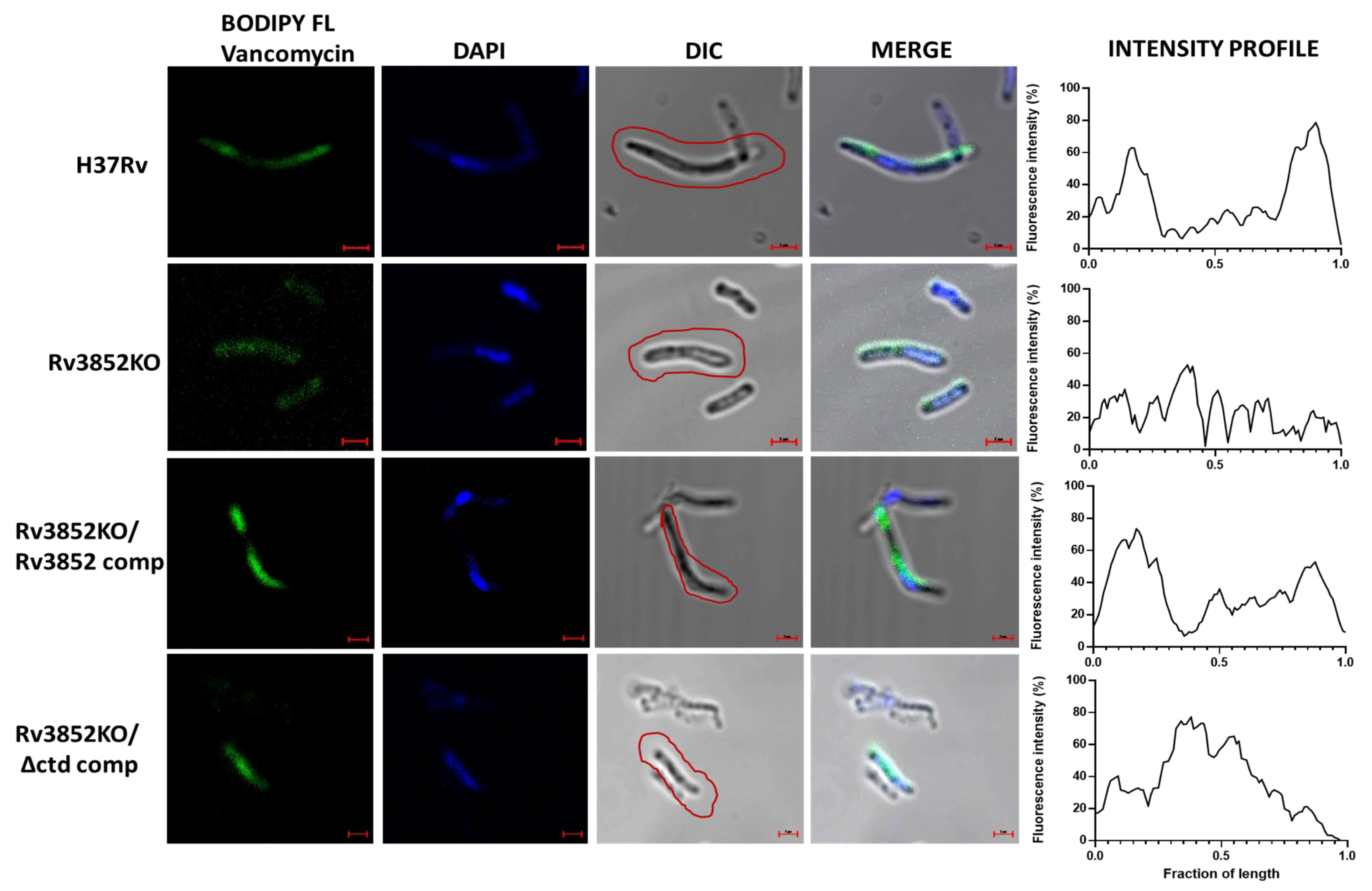
3.7. Absence of Rv3852 Affects Nascent Peptidoglycan Synthesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dorman, C.J.; Deighan, P. Regulation of gene expression by histone-like proteins in bacteria. Curr. Opin. Genet. Dev. 2003, 13, 179–184. [Google Scholar] [CrossRef]
- Dillon, S.C.; Dorman, C.J. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Dorman, C.J. Genome architecture and global gene regulation in bacteria: Making progress towards a unified model? Nat. Rev. Microbiol. 2013, 11, 349–355. [Google Scholar] [CrossRef]
- Betts, J.C.; Lukey, P.T.; Robb, L.C.; McAdam, R.A.; Duncan, K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 2002, 43, 717–731. [Google Scholar] [CrossRef]
- Colangeli, R.; Helb, D.; Vilchèze, C.; Hazbón, M.H.; Lee, C.G.; Safi, H.; Sayers, B.; Sardone, I.; Jones, M.B.; Fleischmann, R.D.; et al. Transcriptional regulation of multi-drug tolerance and antibiotic-induced responses by the histone-like protein Lsr2 in M. tuberculosis. PLoS Pathog. 2007, 3, 0780–0793. [Google Scholar] [CrossRef]
- Stewart, G.R.; Wernisch, L.; Stabler, R.; Mangan, J.A.; Hinds, J.; Laing, K.G.; Young, D.B.; Butcher, P.D. Dissection of the heat-shock response in Mycobacterium tuberculosis using mutants and microarrays. Microbiology 2002, 148, 3129–3138. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Rustad, T.; Mahajan, G.; Kumar, A.; Rao, K.V.S.; Banerjee, S.; Sherman, D.R.; Mande, S.C. Towards understanding the biological function of the unusual chaperonin Cpn60.1 (GroEL1) of Mycobacterium tuberculosis. Tuberculosis 2016, 97, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Dosanjh, N.S.; Rawat, M.; Chung, J.H.; Av-Gay, Y. Thiol specific oxidative stress response in Mycobacteria. FEMS Microbiol. Lett. 2005, 249, 87–94. [Google Scholar] [CrossRef]
- Monahan, I.M.; Betts, J.; Banerjee, D.K.; Butcher, P.D. Differential expression of mycobacterial proteins following phagocytosis by macrophages. Microbiology 2001, 147, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Indi, S.S.; Nagaraja, V. Regulation of lipid biosynthesis, sliding motility, and biofilm formation by a membrane-anchored nucleoid-associated protein of Mycobacterium tuberculosis. J. Bacteriol. 2013, 195, 1769–1778. [Google Scholar] [CrossRef] [Green Version]
- Kriel, N.L.; Gallant, J.; van Wyk, N.; van Helden, P.; Sampson, S.L.; Warren, R.M.; Williams, M.J. Mycobacterial nucleoid associated proteins: An added dimension in gene regulation. Tuberculosis 2018, 108, 169–177. [Google Scholar] [CrossRef]
- Cole, S.T.; Brosch, R.; Parkhill, J.; Garnier, T.; Churcher, C.; Harris, D.; Gordon, S.V.; Eiglmeier, K.; Gas, S.; Barry, C.E.; et al. Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 1998, 393, 537–544. [Google Scholar] [CrossRef]
- Kang, C.-M.; Nyayapathy, S.; Lee, J.-Y.; Suh, J.-W.; Husson, R.N. Wag31, a homologue of the cell division protein DivIVA, regulates growth, morphology and polar cell wall synthesis in mycobacteria. Microbiology 2008, 154, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.-M.; Abbott, D.W.; Park, S.T.; Dascher, C.C.; Cantley, L.C.; Husson, R.N. The Mycobacterium tuberculosis serine/threonine kinases PknA and PknB: Substrate identification and regulation of cell shape. Genes Dev. 2005, 19, 1692–1704. [Google Scholar] [CrossRef] [Green Version]
- Flärdh, K. Essential role of DivlVA in polar growth and morphogenesis in Streptomyces coelicolor A3(2). Mol. Microbiol. 2003, 49, 1523–1536. [Google Scholar] [CrossRef]
- Kaval, K.G.; Rismondo, J.; Halbedel, S. A function of DivIVA in Listeria monocytogenes division site selection. Mol. Microbiol. 2014, 94, 637–654. [Google Scholar] [CrossRef]
- Halbedel, S.; Hahn, B.; Daniel, R.A.; Flieger, A. DivIVA affects secretion of virulence-related autolysins in Listeria monocytogenes. Mol. Microbiol. 2012, 83, 821–839. [Google Scholar] [CrossRef]
- Cha, J.H.; Stewart, G.C. The divIVA minicell locus of Bacillus subtilis. J. Bacteriol. 1997, 179, 1671–1683. [Google Scholar] [CrossRef] [Green Version]
- Marston, A.L.; Thomaides, H.B.; Edwards, D.H.; Sharpe, M.E.; Errington, J. Polar localization of the MinD protein of Bacillus subtilis and its role in selection of the mid-cell division site. Genes Dev. 1998, 12, 3419–3430. [Google Scholar] [CrossRef] [Green Version]
- Marston, A.L.; Errington, J. Selection of the midcell division site in Bacillus subtilis through MinD-dependent polar localization and activation of MinC. Mol. Microbiol. 1999, 33, 84–96. [Google Scholar] [CrossRef]
- Ben-Yehuda, S.; Rudner, D.Z.; Losick, R. RacA, a bacterial protein that anchors chromosomes to the cell poles. Science (80-. ). 2003, 299, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.J.; Errington, J. RacA and the Soj-Spo0J system combine to effect polar chromosome segregation in sporulating Bacillus subtilis. Mol. Microbiol. 2003, 49, 1463–1475. [Google Scholar] [CrossRef] [Green Version]
- Perry, S.E.; Edwards, D.H. The Bacillus subtilis DivIVA protein has a sporulation-specific proximity to Spo0J. J. Bacteriol. 2006, 188, 6039–6043. [Google Scholar] [CrossRef] [Green Version]
- Holmes, N.A.; Walshaw, J.; Leggett, R.M.; Thibessard, A.; Dalton, K.A.; Gillespie, M.D.; Hemmings, A.M.; Gust, B.; Kelemen, G.H. Coiled-coil protein Scy is a key component of a multiprotein assembly controlling polarized growth in Streptomyces. Proc. Natl. Acad. Sci. USA 2013, 110, E397–E406. [Google Scholar] [CrossRef] [Green Version]
- Donovan, C.; Sieger, B.; Krämer, R.; Bramkamp, M. A synthetic Escherichia coli system identifies a conserved origin tethering factor in Actinobacteria. Mol. Microbiol. 2012, 84, 105–116. [Google Scholar] [CrossRef]
- Mukherjee, P.; Sureka, K.; Datta, P.; Hossain, T.; Barik, S.; Das, K.P.; Kundu, M.; Basu, J. Novel role of Wag31 in protection of mycobacteria under oxidative stress. Mol. Microbiol. 2009, 73, 103–119. [Google Scholar] [CrossRef]
- Meniche, X.; Otten, R.; Siegrist, M.S.; Baer, C.E.; Murphy, K.C.; Bertozzi, C.R.; Sassetti, C.M. Subpolar addition of new cell wall is directed by DivIVA in mycobacteria. Proc. Natl. Acad. Sci. USA 2014, 111, E3243–E3251. [Google Scholar] [CrossRef] [Green Version]
- Carel, C.; Nukdee, K.; Cantaloube, S.; Bonne, M.; Diagne, C.T.; Laval, F.; Daffé, M.; Zerbib, D. Mycobacterium tuberculosis proteins involved in mycolic acid synthesis and transport localize dynamically to the old growing pole and septum. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Kaval, K.G.; Hauf, S.; Rismondo, J.; Hahn, B.; Halbedel, S. Genetic dissection of DivIVA functions in Listeria monocytogenes. J. Bacteriol. 2017, 199, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ginda, K.; Bezulska, M.; Ziólkiewicz, M.; Dziadek, J.; Zakrzewska-Czerwińska, J.; Jakimowicz, D. ParA of Mycobacterium smegmatis co-ordinates chromosome segregation with the cell cycle and interacts with the polar growth determinant DivIVA. Mol. Microbiol. 2013, 87, 998–1012. [Google Scholar] [CrossRef]
- Pióro, M.; Małecki, T.; Portas, M.; Magierowska, I.; Trojanowski, D.; Sherratt, D.; Zakrzewska-Czerwińska, J.; Ginda, K.; Jakimowicz, D. Competition between DivIVA and the nucleoid for ParA binding promotes segrosome separation and modulates mycobacterial cell elongation. Mol. Microbiol. 2019, 111, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Free, R.B.; Hazelwood, L.A.; Sibley, D.R. Identifying novel protein-protein interactions using co-immunoprecipitation and mass spectroscopy. Curr. Protoc. Neurosci. 2009. [CrossRef] [PubMed] [Green Version]
- Datta, C.; Jha, R.K.; Ahmed, W.; Ganguly, S.; Ghosh, S.; Nagaraja, V. Physical and functional interaction between nucleoid-associated proteins HU and Lsr2 of Mycobacterium tuberculosis: Altered DNA binding and gene regulation. Mol. Microbiol. 2019, 111, 981–994. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Carette, X.; Potluri, L.-P.; Sharp, J.D.; Xu, R.; Prisic, S.; Husson, R.N. Investigating essential gene function in Mycobacterium tuberculosis using an efficient CRISPR interference system. Nucleic Acids Res. 2016, 44, e143. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Dhar, N.; Pató, J.; Kolly, G.S.; Korduláková, J.; Forbak, M.; Evans, J.C.; Székely, R.; Rybniker, J.; Palčeková, Z.; et al. Identification of aminopyrimidine-sulfonamides as potent modulators of Wag31-mediated cell elongation in mycobacteria. Mol. Microbiol. 2017, 103, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Rezwan, M.; Lanéelle, M.-A.; Sander, P.; Daffé, M. Breaking down the wall: Fractionation of mycobacteria. J. Microbiol. Methods 2007, 68, 32–39. [Google Scholar] [CrossRef]
- Thanky, N.R.; Young, D.B.; Robertson, B.D. Unusual features of the cell cycle in mycobacteria: Polar-restricted growth and the snapping-model of cell division. Tuberculosis 2007, 87, 231–236. [Google Scholar] [CrossRef]
- Sassetti, C.M.; Boyd, D.H.; Rubin, E.J. Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 2003, 48, 77–84. [Google Scholar] [CrossRef] [PubMed]
- de Souza, G.A.; Leversen, N.A.; Målen, H.; Wiker, H.G. Bacterial proteins with cleaved or uncleaved signal peptides of the general secretory pathway. J. Proteomics 2011, 75, 502–510. [Google Scholar] [CrossRef] [Green Version]
- García-Heredia, A.; Pohane, A.A.; Melzer, E.S.; Carr, C.R.; Fiolek, T.J.; Rundell, S.R.; Lim, H.C.; Wagner, J.C.; Morita, Y.S.; Swarts, B.M.; et al. Peptidoglycan precursor synthesis along the sidewall of pole-growing mycobacteria. Elife 2018, 7. [Google Scholar] [CrossRef]
- Jani, C.; Eoh, H.; Lee, J.J.; Hamasha, K.; Sahana, M.B.; Han, J.S.; Nyayapathy, S.; Lee, J.Y.; Suh, J.W.; Lee, S.H.; et al. Regulation of polar peptidoglycan biosynthesis by Wag31 phosphorylation in mycobacteria. BMC Microbiol. 2010, 10, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, T.A.; Zupan, J.R.; Zambryski, P.C. The essential features and modes of bacterial polar growth. Trends Microbiol. 2015, 23, 347–353. [Google Scholar] [CrossRef]
- Nguyen, L.; Scherr, N.; Gatfield, J.; Walburger, A.; Pieters, J.; Thompson, C.J. Antigen 84, an effector of pleiomorphism in Mycobactenum smegmatis. J. Bacteriol. 2007, 189, 7896–7910. [Google Scholar] [CrossRef] [Green Version]
- Melzer, E.S.; Sein, C.E.; Chambers, J.J.; Sloan Siegrist, M. DivIVA concentrates mycobacterial cell envelope assembly for initiation and stabilization of polar growth. Cytoskeleton 2018, 75, 498–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Baarle, S.; Bramkamp, M. The MinCDJ system in Bacillus subtilis prevents minicell formation by promoting divisome disassembly. PLoS One 2010, 5. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.X.; Zhang, L.; Mai, J.T.; Peng, R.C.; Yang, E.Z.; Peng, C.; Wang, H.H. The Wag31 protein interacts with AccA3 and coordinates cell wall lipid permeability and lipophilic drug resistance in Mycobacterium smegmatis. Biochem. Biophys. Res. Commun. 2014, 448, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Plocinski, P.; Martinez, L.; Sarva, K.; Plocinska, R.; Madiraju, M.; Rajagopalan, M. Mycobacterium tuberculosis CwsA overproduction modulates cell division and cell wall synthesis. Tuberculosis 2013, 93, S21–S27. [Google Scholar] [CrossRef]
- Plocinski, P.; Arora, N.; Sarva, K.; Blaszczyk, E.; Qin, H.; Das, N.; Plocinska, R.; Ziolkiewicz, M.; Dziadek, J.; Kiran, M.; et al. Mycobacterium tuberculosis CwsA interacts with CrgA and Wag31, and the CrgA-CwsA complex is involved in peptidoglycan synthesis and cell shape determination. J. Bacteriol. 2012, 194, 6398–6409. [Google Scholar] [CrossRef] [Green Version]
- Ricci, D.P.; Melfi, M.D.; Lasker, K.; Dill, D.L.; McAdams, H.H.; Shapiro, L. Cell cycle progression in Caulobacter requires a nucleoid-associated protein with high AT sequence recognition. Proc. Natl. Acad. Sci. USA 2016, 113, E5952–E5961. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.A.; Panis, G.; Viollier, P.H.; Marczynski, G.T. A novel nucleoid-associated protein coordinates chromosome replication and chromosome partition. Nucleic Acids Res. 2017, 45, 8916–8929. [Google Scholar] [CrossRef] [Green Version]
- Finkel, S.E.; Johnson, R.C. The Fis protein: It’s not just for DNA inversion anymore. Mol. Microbiol. 1992, 6, 3257–3265. [Google Scholar] [CrossRef] [PubMed]
- Ben-Yehuda, S.; Fujita, M.; Liu, X.S.; Gorbatyuk, B.; Skoko, D.; Yan, J.; Marko, J.F.; Liu, J.S.; Eichenberger, P.; Rudner, D.Z.; et al. Defining a centromere-like element in Bacillus subtilis by identifying the binding sites for the chromosome-anchoring protein RacA. Mol. Cell 2005, 17, 773–782. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garg, R.; Anand, C.; Ganguly, S.; Rao, S.; Verma, R.; Nagaraja, V. Direct Interaction of Polar Scaffolding Protein Wag31 with Nucleoid-Associated Protein Rv3852 Regulates Its Polar Localization. Cells 2021, 10, 1558. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061558
Garg R, Anand C, Ganguly S, Rao S, Verma R, Nagaraja V. Direct Interaction of Polar Scaffolding Protein Wag31 with Nucleoid-Associated Protein Rv3852 Regulates Its Polar Localization. Cells. 2021; 10(6):1558. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061558
Chicago/Turabian StyleGarg, Rajni, Chinmay Anand, Sohini Ganguly, Sandhya Rao, Rinkee Verma, and Valakunja Nagaraja. 2021. "Direct Interaction of Polar Scaffolding Protein Wag31 with Nucleoid-Associated Protein Rv3852 Regulates Its Polar Localization" Cells 10, no. 6: 1558. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061558