
High-Fat Diet Alters the Retinal Transcriptome in the Absence of Gut Microbiota

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Sterility Monitoring
2.3. RNA Extraction
2.4. RNA Sequencing
2.5. Statistical Analysis
3. Results
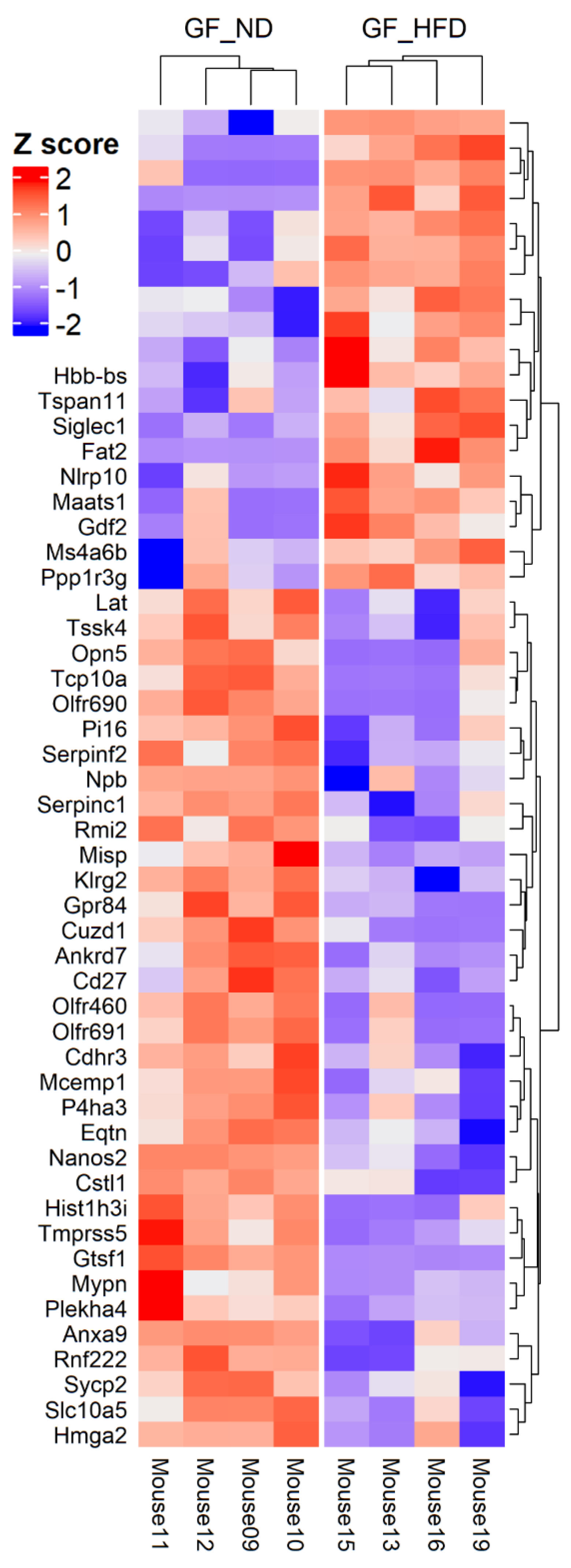
3.1. HFD Is Associated with Differential Retinal Gene Expression in the Absence of the Microbiome
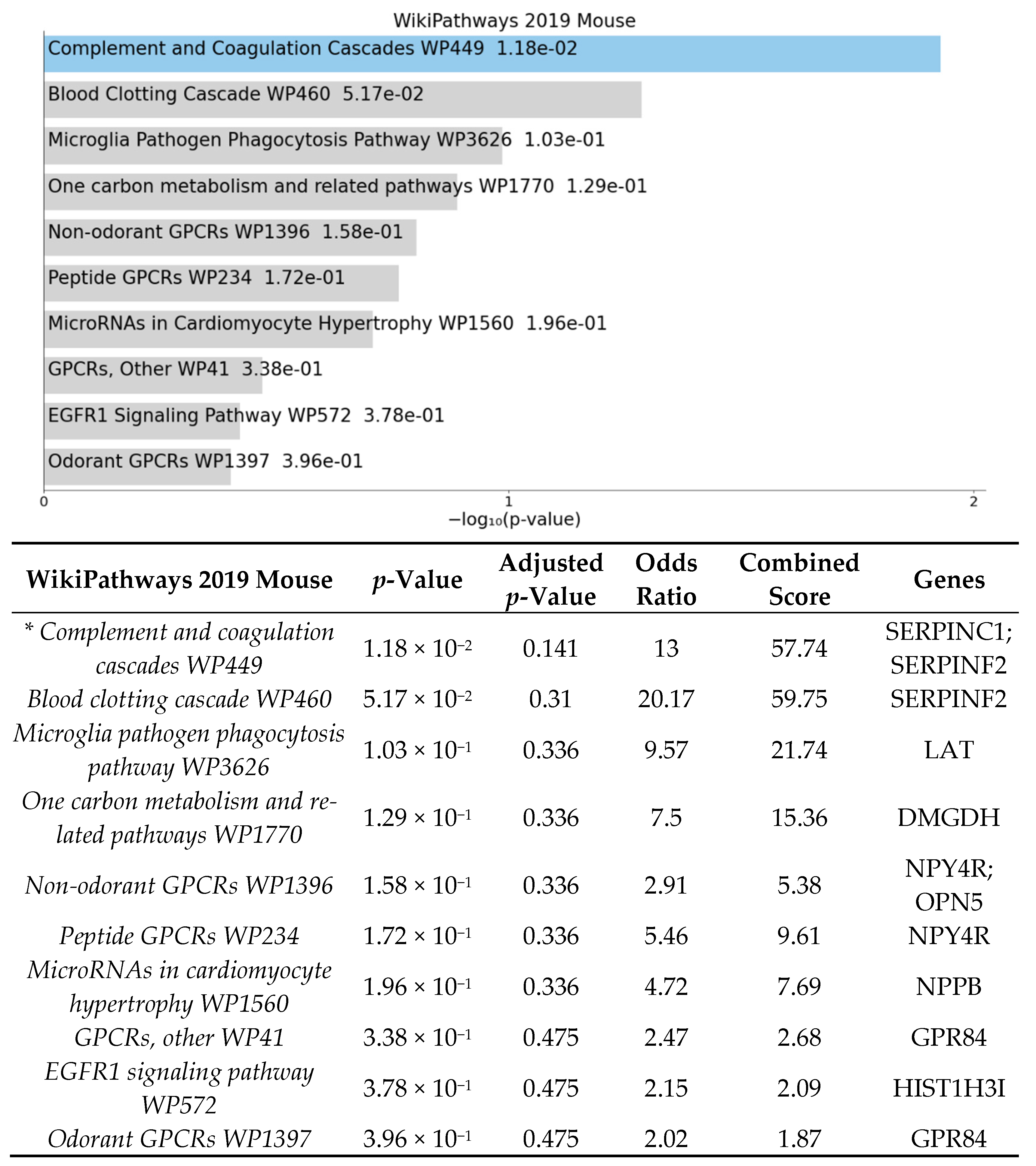
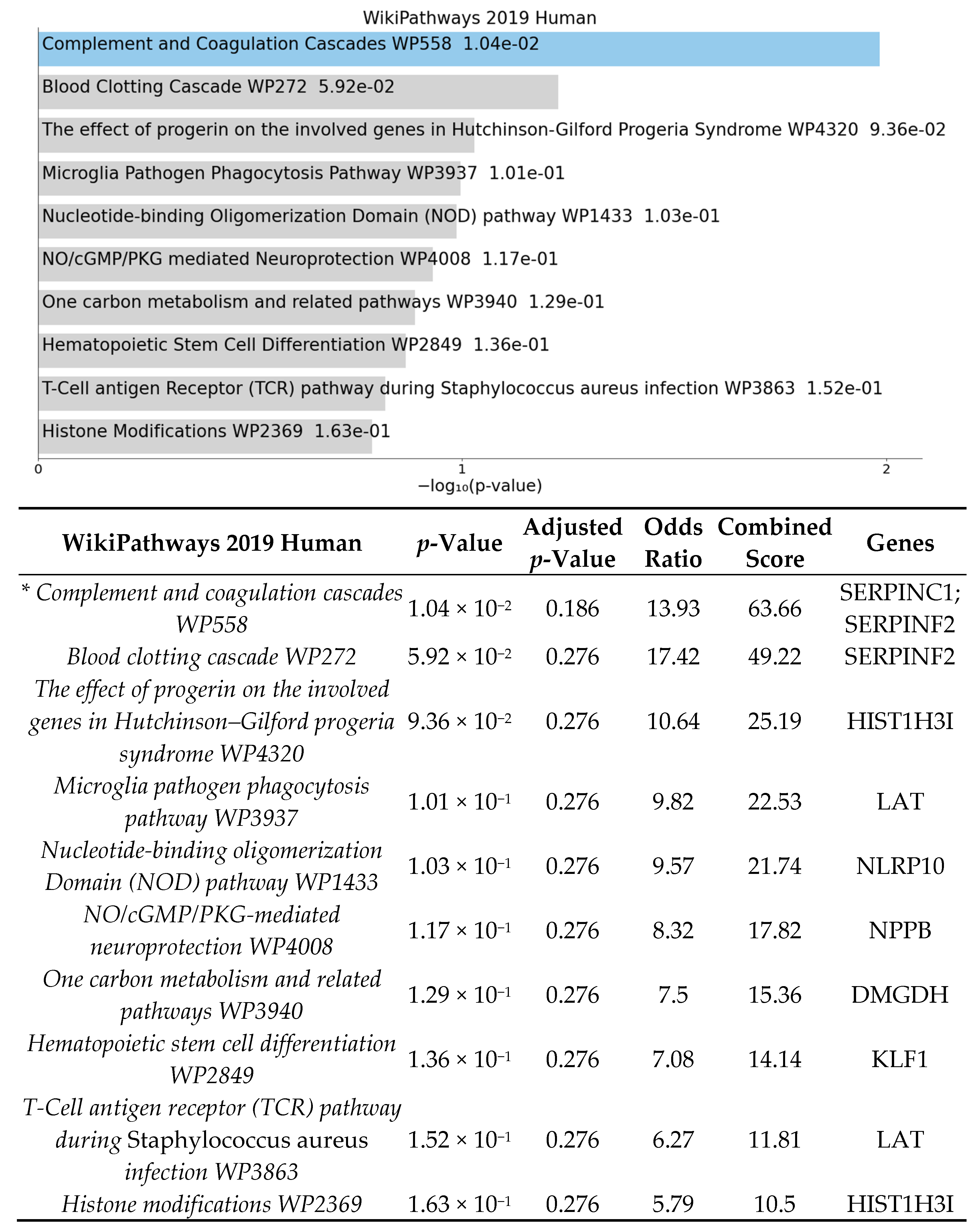
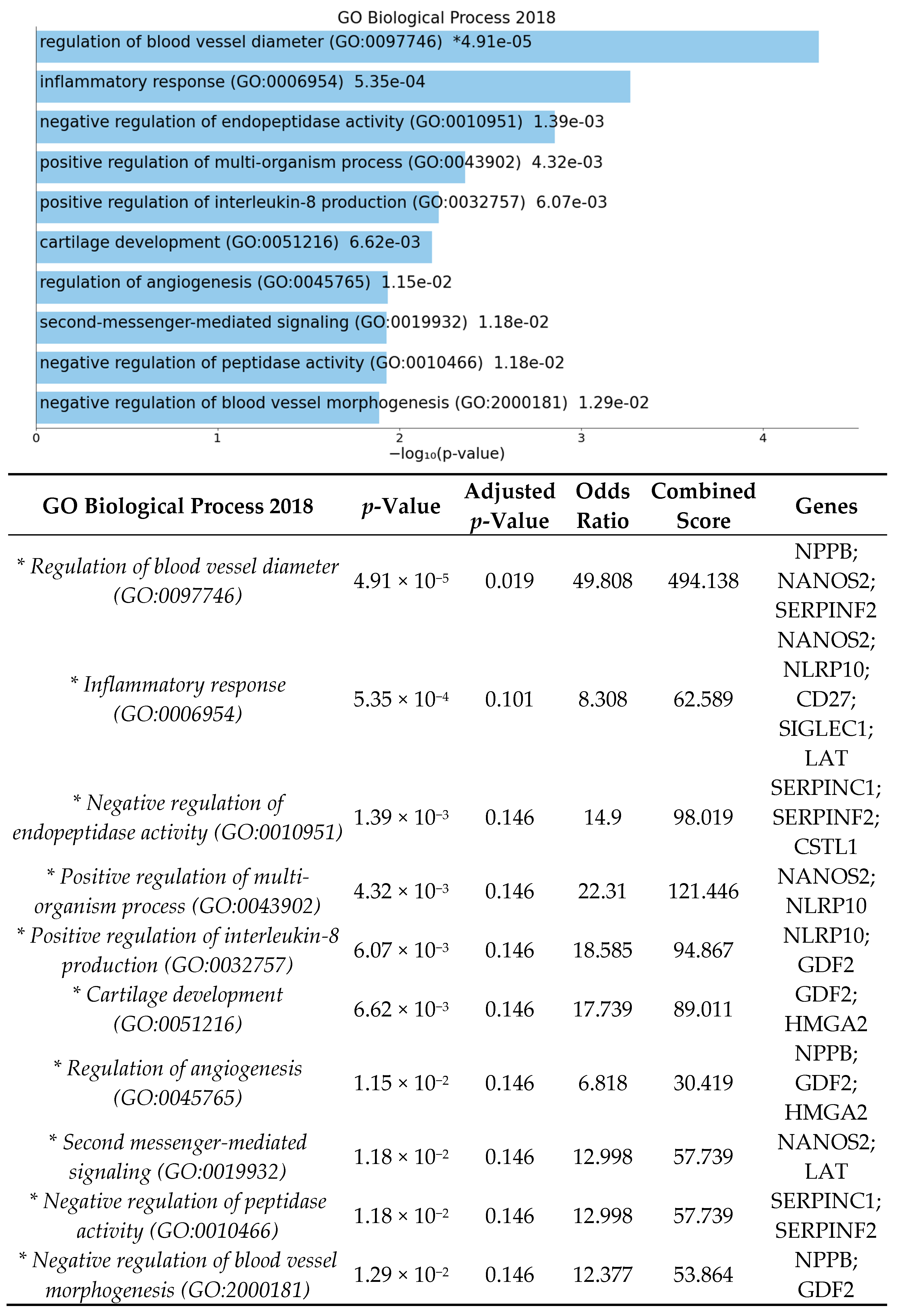
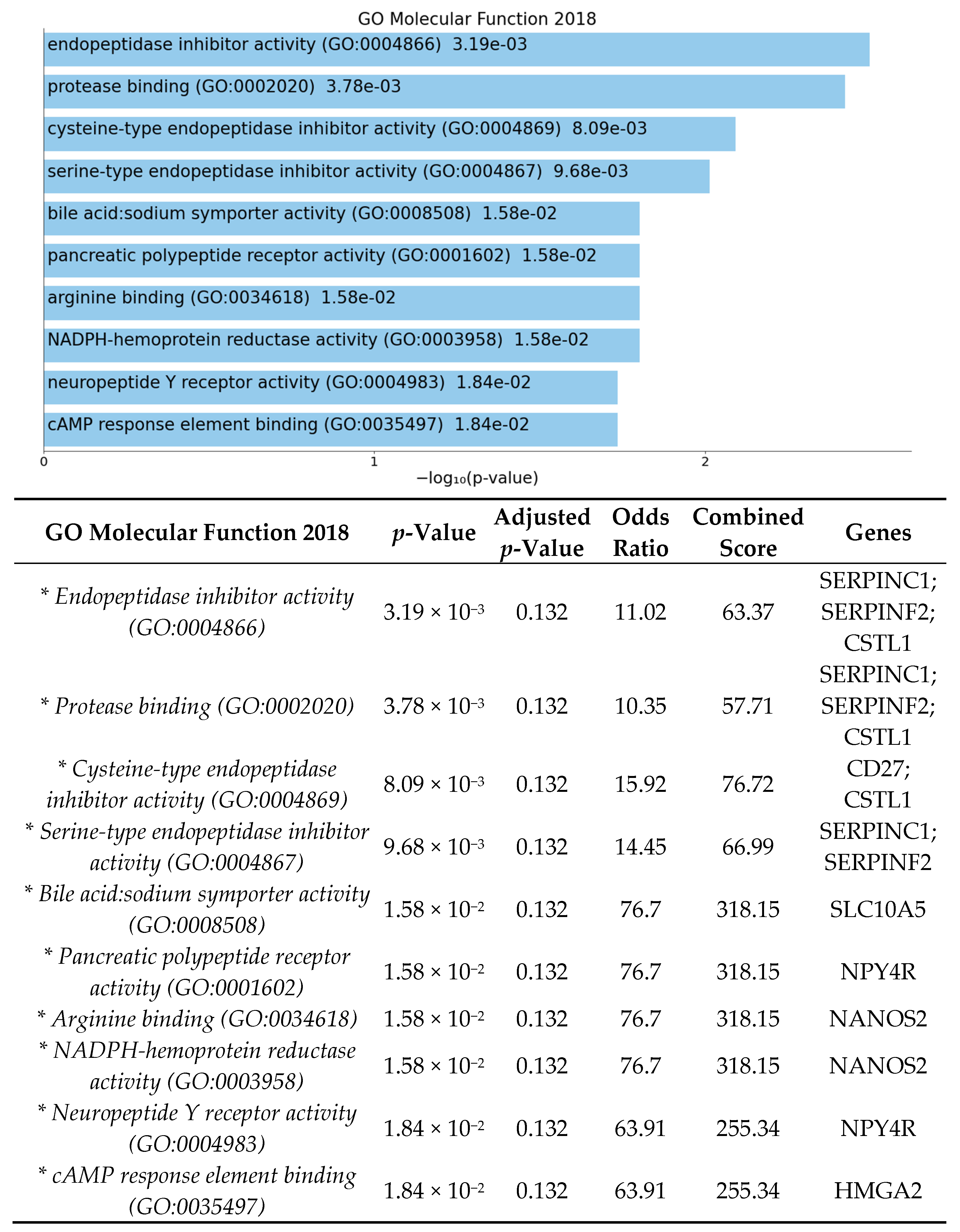
3.2. Significant Biologic Functions and Processes Are Overrepresented by Functional Enrichment Analysis
4. Discussion
4.1. High-Fat Diet May Affect Expression of Genes Involved in Inflammatory Pathways in Germ-Free Mice
4.2. High-Fat Diet May Influence Genes and Pathways Involved in Angiogenesis in Germ-Free Mice
4.3. High-Fat Diet May Affect Expression of Genes Involved in RPE Function and Ciliogenesis in Germ-Free Mice
4.4. Additional Genes and Pathways of Retinal Transcriptome Affected by High-Fat Diet in Germ-Free Mice
5. Conclusions and Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang, J.H.; Willett, W.C.; Rosner, B.A.; Buys, E.; Wiggs, J.L.; Pasquale, L.R. Association of Dietary Nitrate Intake With Primary Open-Angle Glaucoma: A Prospective Analysis From the Nurses’ Health Study and Health Professionals Follow-up Study. JAMA Ophthalmol. 2016, 134, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.H.; Pasquale, L.R.; Willett, W.C.; Rosner, B.A.; Egan, K.M.; Faberowski, N.; Hankinson, S.E. Dietary Fat Consumption and Primary Open-Angle Glaucoma. Am. J. Clin. Nutr. 2004, 79, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weikel, K.A.; Chiu, C.-J.; Taylor, A. Nutritional Modulation of Age-Related Macular Degeneration. Mol. Asp. Med. 2012, 33, 318–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Mele, M.C.; Merendino, N.; Cintoni, M.; Anselmi, G.; Caporossi, A.; Gasbarrini, A.; Minnella, A.M. The Role of Diet, Micronutrients and the Gut Microbiota in Age-Related Macular Degeneration: New Perspectives from the Gut–Retina Axis. Nutrients 2018, 10, 1677. [Google Scholar] [CrossRef] [Green Version]
- Zemski Berry, K.A.; Gordon, W.C.; Murphy, R.C.; Bazan, N.G. Spatial Organization of Lipids in the Human Retina and Optic Nerve by MALDI Imaging Mass Spectrometry. J. Lipid Res. 2014, 55, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Wong-Riley, M.T.T. Energy Metabolism of the Visual System. Eye Brain 2010, 2, 99–116. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.-J. The Relationship of Major American Dietary Patterns to Age-Related Macular Degeneration. Am. J. Ophthalmol. 2014, 158, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parekh, N.; Voland, R.P.; Moeller, S.M.; Blodi, B.A.; Ritenbaugh, C.; Chappell, R.J.; Wallace, R.B.; Mares, J.A.; CAREDS Research Study Group. Association between Dietary Fat Intake and Age-Related Macular Degeneration in the Carotenoids in Age-Related Eye Disease Study (CAREDS): An Ancillary Study of the Women’s Health Initiative. Arch. Ophthalmol. 2009, 127, 1483–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressler, N.M. Age-Related Macular Degeneration Is the Leading Cause of Blindness. JAMA 2004, 291, 1900–1901. [Google Scholar] [CrossRef] [PubMed]
- Chong, E.W.-T.; Robman, L.D.; Simpson, J.A.; Hodge, A.M.; Aung, K.Z.; Dolphin, T.K.; English, D.R.; Giles, G.G.; Guymer, R.H. Fat Consumption and Its Association with Age-Related Macular Degeneration. Arch. Ophthalmol. 2009, 127, 674–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson-Townsend, D.A.; Douglass, A.J.; Singh, A.; Allen, R.S.; Uwaifo, I.N.; Pardue, M.T. Impacts of High Fat Diet on Ocular Outcomes in Rodent Models of Visual Disease. Exp. Eye Res. 2021, 204, 108440. [Google Scholar] [CrossRef]
- Chang, R.C.-A.; Shi, L.; Huang, C.C.-Y.; Kim, A.J.; Ko, M.L.; Zhou, B.; Ko, G.Y.-P. High-Fat Diet-Induced Retinal Dysfunction. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2367–2380. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-J.; Wang, P.-W.; Yang, I.-H.; Huang, H.-M.; Chang, C.-S.; Wu, C.-L.; Chuang, J.-H. High-Fat Diet Induces Toll-like Receptor 4-Dependent Macrophage/Microglial Cell Activation and Retinal Impairment. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3041–3050. [Google Scholar] [CrossRef] [Green Version]
- Albouery, M.; Buteau, B.; Grégoire, S.; Martine, L.; Gambert, S.; Bron, A.M.; Acar, N.; Chassaing, B.; Bringer, M.-A. Impact of a High-Fat Diet on the Fatty Acid Composition of the Retina. Exp. Eye Res. 2020, 196, 108059. [Google Scholar] [CrossRef] [PubMed]
- Skondra, D.; She, H.; Zambarakji, H.J.; Connolly, E.; Michaud, N.; Chan, P.; Kim, I.K.; Gragoudas, E.S.; Miller, J.W.; Hafezi-Moghadam, A. Effects of ApoE Deficiency, Aging and High Fat Diet on Laser-Induced Choroidal Neovascularization and Bruch’s Membrane-RPE Interface Morphology. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1768. [Google Scholar]
- Asare-Bediako, B.; Noothi, S.K.; Li Calzi, S.; Athmanathan, B.; Vieira, C.P.; Adu-Agyeiwaah, Y.; Dupont, M.; Jones, B.A.; Wang, X.X.; Chakraborty, D.; et al. Characterizing the Retinal Phenotype in the High-Fat Diet and Western Diet Mouse Models of Prediabetes. Cells 2020, 9, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, I.N.; Hafez, S.S.; Fairaq, A.; Ergul, A.; Imig, J.D.; El-Remessy, A.B. Thioredoxin-Interacting Protein Is Required for Endothelial NLRP3 Inflammasome Activation and Cell Death in a Rat Model of High-Fat Diet. Diabetologia 2014, 57, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Andriessen, E.M.; Wilson, A.M.; Mawambo, G.; Dejda, A.; Miloudi, K.; Sennlaub, F.; Sapieha, P. Gut Microbiota Influences Pathological Angiogenesis in Obesity-driven Choroidal Neovascularization. EMBO Mol. Med. 2016, 8, 1366–1379. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the Normal Gut Microbiota. World J. Gastroenterol. WJG 2015, 21, 8787–8803. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, K.E.; Lynch, S.V. Microbiota in Allergy and Asthma and the Emerging Relationship with the Gut Microbiome. Cell Host Microbe 2015, 17, 592–602. [Google Scholar] [CrossRef] [Green Version]
- Mohajeri, M.H.; Fata, G.L.; Steinert, R.E.; Weber, P. Relationship between the Gut Microbiome and Brain Function. Nutr. Rev. 2018, 76, 481–496. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Kitai, T.; Hazen, S.L. Gut Microbiota in Cardiovascular Health and Disease. Circ. Res. 2017, 120, 1183–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Zhou, Q.; Li, H. Gut Microbiota and Nonalcoholic Fatty Liver Disease: Insights on Mechanisms and Therapy. Nutrients 2017, 9, 1124. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Taylor, A. The Role of Microbiota in Retinal Disease. Adv. Exp. Med. Biol. 2018, 1074, 429–435. [Google Scholar] [PubMed]
- Rowan, S.; Jiang, S.; Korem, T.; Szymanski, J.; Chang, M.-L.; Szelog, J.; Cassalman, C.; Dasuri, K.; McGuire, C.; Nagai, R.; et al. Involvement of a Gut-Retina Axis in Protection against Dietary Glycemia-Induced Age-Related Macular Degeneration. Proc. Natl. Acad. Sci. USA 2017, 114, E4472–E4481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.; Zhang, S.; Li, Q.; Zuo, C.; Gao, X.; Zheng, B.; Lin, M. Gut Microbiota Compositional Profile and Serum Metabolic Phenotype in Patients with Primary Open-Angle Glaucoma. Exp. Eye Res. 2020, 191, 107921. [Google Scholar] [CrossRef] [PubMed]
- Zinkernagel, M.S.; Zysset-Burri, D.C.; Keller, I.; Berger, L.E.; Leichtle, A.B.; Largiadèr, C.R.; Fiedler, G.M.; Wolf, S. Association of the Intestinal Microbiome with the Development of Neovascular Age-Related Macular Degeneration. Sci. Rep. 2017, 7, 40826. [Google Scholar] [CrossRef] [Green Version]
- Zysset-Burri, D.C.; Keller, I.; Berger, L.E.; Neyer, P.J.; Steuer, C.; Wolf, S.; Zinkernagel, M.S. Retinal Artery Occlusion Is Associated with Compositional and Functional Shifts in the Gut Microbiome and Altered Trimethylamine-N-Oxide Levels. Sci. Rep. 2019, 9, 15303. [Google Scholar] [CrossRef]
- Skondra, D.; Rodriguez, S.H.; Sharma, A.; Gilbert, J.; Andrews, B.; Claud, E.C. The Early Gut Microbiome Could Protect against Severe Retinopathy of Prematurity. J. AAPOS Off. Publ. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2020, 24, 236–238. [Google Scholar] [CrossRef]
- Nadeem, U.; Xie, B.; Movahedan, A.; D’Souza, M.; Barba, H.; Deng, N.; Leone, V.A.; Chang, E.; Sulakhe, D.; Skondra, D. High Throughput RNA Sequencing of Mice Retina Reveals Metabolic Pathways Involved in the Gut-Retina Axis. bioRxiv 2020. [Google Scholar] [CrossRef]
- Orešič, M.; Seppänen-Laakso, T.; Yetukuri, L.; Bäckhed, F.; Hänninen, V. Gut Microbiota Affects Lens and Retinal Lipid Com-Position. Exp. Eye Res. 2009, 89, 604–607. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Do, M.H.; Lee, E.; Oh, M.-J.; Kim, Y.; Park, H.-Y. High-Glucose or -Fructose Diet Cause Changes of the Gut Microbiota and Metabolic Disorders in Mice without Body Weight Change. Nutrients 2018, 10, 761. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of High-Fat Diet on Gut Microbiota: A Driving Force for Chronic Disease Risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515–520. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theriault, B.; Wang, Y.; Chen, L.; Vest, A.; Bartman, C.; Alegre, M.-L. Long-Term Maintenance of Sterility After Skin Transplantation in Germ-Free Mice. Transplant. Direct 2015, 1, e28. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. From Reads to Genes to Pathways: Differential Expression Analysis of RNA-Seq Experiments Using Rsubread and the EdgeR Quasi-Likelihood Pipeline. F1000Research 2016, 5, 1438. [Google Scholar] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Kuleshov, M.V. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, 90–97. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keenan, T.D.; Agrón, E.; Mares, J.; Clemons, T.E.; van Asten, F.; Swaroop, A.; Chew, E.Y.; Age-Related Eye Disease Studies (AREDS) 1 and 2 Research Groups. Adherence to the Mediterranean Diet and Progression to Late Age-Related Macular Degeneration in the Age-Related Eye Disease Studies 1 and 2. Ophthalmology 2020, 127, 1515–1528. [Google Scholar] [CrossRef]
- Dow, C.; Mancini, F.; Rajaobelina, K.; Boutron-Ruault, M.-C.; Balkau, B.; Bonnet, F.; Fagherazzi, G. Diet and Risk of Diabetic Retinopathy: A Systematic Review. Eur. J. Epidemiol. 2018, 33, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.E.; Tseng, V.L.; Yu, F.; Caprioli, J.; Coleman, A.L. Association of Dietary Fatty Acid Intake With Glaucoma in the United States. JAMA Ophthalmol. 2018, 136, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrón, E.; Mares, J.; Clemons, T.E.; Swaroop, A.; Chew, E.Y.; Keenan, T.D.L.; AREDS and AREDS2 Research Groups. Dietary Nutrient Intake and Progression to Late Age-Related Macular Degeneration in the Age-Related Eye Disease Studies 1 and 2. Ophthalmology 2021, 128, 425–442. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.A.; King, K.Y.; Baldridge, M.T. Mouse Microbiota Models: Comparing Germ-Free Mice and Antibiotics Treatment as Tools for Modifying Gut Bacteria. Front. Physiol. 2018, 9, 1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, D.S.; Schmidt-Erfurth, U.; Lookeren Campagne, M.; Henry, E.C.; Brittain, C. The Pathophysiology of Geographic Atrophy Secondary to Age-Related Macular Degeneration and the Complement Pathway as a Therapeutic Target. Retina 2017, 37, 819–835. [Google Scholar] [CrossRef] [PubMed]
- Lorés-Motta, L.; Paun, C.C.; Corominas, J.; Pauper, M.; Geerlings, M.J.; Altay, L.; Schick, T.; Daha, M.R.; Fauser, S.; Hoyng, C.B.; et al. Genome-Wide Association Study Reveals Variants in CFH and CFHR4 Associated with Systemic Complement Activation: Implications in Age-Related Macular Degeneration. Ophthalmology 2018, 125, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- Kassa, E.; Ciulla, T.A.; Hussain, R.M.; Dugel, P.U. Complement Inhibition as a Therapeutic Strategy in Retinal Disorders. Expert Opin. Biol. Ther. 2019, 19, 335–342. [Google Scholar] [CrossRef]
- Lee, J.; Yoo, J.H.; Kim, H.S.; Cho, Y.K.; Lee, Y.L.; Lee, W.J.; Park, J.-Y.; Jung, C.H. C1q/TNF-Related Protein-9 Attenuates Palmitic Acid-Induced Endothelial Cell Senescence via Increasing Autophagy. Mol. Cell. Endocrinol. 2021, 521, 111114. [Google Scholar] [CrossRef]
- Chavali, V.R.M.; Sommer, J.R.; Petters, R.M.; Ayyagari, R. Identification of a Promoter for the Human C1q-Tumor Necrosis Factor–Related Protein-5 Gene Associated with Late-Onset Retinal Degeneration. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5499–5507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinomiya, K.; Mukai, A.; Yoneda, K.; Ueno, M.; Sotozono, C.; Kinoshita, S.; Hamuro, J. The High Expression of C1q and Tumor Necrosis Factor- Related Protein (CTRP) 6, a New Complement Regulatory Factor, in the Drusen of Age-Related Macular Degeneration Eyes. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5013. [Google Scholar]
- Cheng, Y.; Qi, Y.; Liu, S.; Di, R.; Shi, Q.; Li, J.; Pei, C. C1q/TNF-Related Protein 9 Inhibits High Glucose-Induced Oxidative Stress and Apoptosis in Retinal Pigment Epithelial Cells Through the Activation of AMPK/Nrf2 Signaling Pathway. Cell Transplant. 2020, 29, 963689720962052. [Google Scholar] [CrossRef] [PubMed]
- Unterholzner, L.; Keating, S.E.; Baran, M.; Horan, K.A.; Jensen, S.B.; Sharma, S.; Sirois, C.M.; Jin, T.; Latz, E.; Xiao, T.S.; et al. IFI16 Is an Innate Immune Sensor for Intracellular DNA. Nat. Immunol. 2010, 11, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yu, S.-X.; Zhou, F.-H.; Zhang, X.-J.; Gao, W.-Y.; Li, K.-Y.; Liu, Z.-Z.; Han, W.-Y.; Yang, Y.-J. DNA Sensor IFI204 Contributes to Host Defense Against Staphylococcus Aureus Infection in Mice. Front. Immunol. 2019, 10, 474. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Muckersie, E.; Forrester, J.V.; Xu, H. Immune Activation in Retinal Aging: A Gene Expression Study. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5888–5896. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Luo, C.; Zhao, J.; Devarajan, G.; Xu, H. Immune Regulation in the Aging Retina. Prog. Retin. Eye Res. 2019, 69, 159–172. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Q.; Kowluru, R.A. Role of Histone Acetylation in the Development of Diabetic Retinopathy and the Metabolic Memory Phenomenon. J Cell Biochem. 2010, 110, 1306–1313. [Google Scholar] [CrossRef] [Green Version]
- Villeneuve, L.M.; Reddy, M.A.; Lanting, L.L.; Wang, M.; Meng, L.; Natarajan, R. Epigenetic Histone H3 Lysine 9 Methylation in Metabolic Memory and Inflammatory Phenotype of Vascular Smooth Muscle Cells in Diabetes. Proc. Natl. Acad. Sci. USA 2008, 105, 9047–9052. [Google Scholar] [CrossRef] [Green Version]
- Banday, A.R.; Baumgartner, M.; Al Seesi, S.; Karunakaran, D.K.P.; Venkatesh, A.; Congdon, S.; Lemoine, C.; Kilcollins, A.M.; Mandoiu, I.; Punzo, C.; et al. Replication-Dependent Histone Genes Are Actively Transcribed in Differentiating and Aging Retinal Neurons. Cell Cycle 2014, 13, 2526–2541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, F.R.; Chader, G.J.; Johnson, L.V.; Tombran-Tink, J. Pigment Epithelium-Derived Factor: Neurotrophic Activity and Identification as a Member of the Serine Protease Inhibitor Gene Family. Proc. Natl. Acad. Sci. USA 1993, 90, 1526–1530. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Cheng, R.; Benyajati, S.; Ma, J.-X. PEDF and Its Roles in Physiological and Pathological Conditions: Implication in Diabetic and Hypoxia-Induced Angiogenic Diseases. Clin. Sci. 2015, 128, 805–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youngblood, H.; Robinson, R.; Sharma, A.; Sharma, S. Proteomic Biomarkers of Retinal Inflammation in Diabetic Retinopathy. Int. J. Mol. Sci. 2019, 20, 4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, B.; Ni, J.; Vatish, M.; Singer, D.R.J. Xanthine Oxidase Interaction with Vascular Endothelial Growth Factor in Human Endothelial Cell Angiogenesis. Microcirculation 2008, 15, 251–267. [Google Scholar] [CrossRef] [PubMed]
- Elmasry, K.; Ibrahim, A.S.; Abdulmoneim, S.; Al-Shabrawey, M. Bioactive Lipids and Pathological Retinal Angiogenesis. Br. J. Pharmacol. 2019, 176, 93–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadeqzadeh, E.; Bock, C.E.; Thorne, R.F. Sleeping Giants: Emerging Roles for the Fat Cadherins in Health and Disease. Med. Res. Rev. 2014, 34, 190–221. [Google Scholar] [CrossRef] [PubMed]
- Horne-Badovinac, S. Fat-like Cadherins in Cell Migration—Leading from Both the Front and the Back. Curr. Opin. Cell Biol. 2017, 48, 26–32. [Google Scholar] [CrossRef]
- Cao, J.; Ehling, M.; März, S.; Seebach, J.; Tarbashevich, K.; Sixta, T.; Pitulescu, M.E.; Werner, A.-C.; Flach, B.; Montanez, E.; et al. Polarized Actin and VE-Cadherin Dynamics Regulate Junctional Remodelling and Cell Migration during Sprouting Angiogenesis. Nat. Commun. 2017, 8, 2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.-T.T. An Opsin 5–Dopamine Pathway Mediates Light-Dependent Vascular Development in the Eye. Nat. Cell Biol. 2019, 21, 420–429. [Google Scholar] [CrossRef]
- Buhr, E.D.; Yue, W.W.S.; Ren, X.; Jiang, Z.; Liao, H.-W.R.; Mei, X.; Vemaraju, S.; Nguyen, M.-T.; Reed, R.R.; Lang, R.A.; et al. Neuropsin (OPN5)-Mediated Photoentrainment of Local Circadian Oscillators in Mammalian Retina and Cornea. Proc. Natl. Acad. Sci. USA 2015, 112, 13093–13098. [Google Scholar] [CrossRef] [Green Version]
- Aaltonen, V.; Kinnunen, K.; Jouhilahti, E.-M.; Peltonen, J.; Nikinmaa, M.; Kaarniranta, K.; Arjamaa, O. Hypoxic Conditions Stimulate the Release of B-Type Natriuretic Peptide from Human Retinal Pigment Epithelium Cell Culture. Acta Ophthalmol. (Copenh.) 2014, 92, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Rollín, R.; Mediero, A.; Roldán-Pallarés, M.; Fernández-Cruz, A.; Fernández-Durango, R. Natriuretic Peptide System in the Human Retina. Mol. Vis. 2004, 10, 15–22. [Google Scholar]
- Jovancevic, N.; Wunderlich, K.A.; Haering, C.; Flegel, C.; Maßberg, D.; Weinrich, M.; Weber, L.; Tebbe, L.; Kampik, A.; Gisselmann, G.; et al. Deep Sequencing of the Human Retinae Reveals the Expression of Odorant Receptors. Front. Cell. Neurosci. 2017, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Pavan, B.; Capuzzo, A.; Dalpiaz, A. Potential Therapeutic Effects of Odorants through Their Ectopic Receptors in Pigmented Cells. Drug Discov. Today 2017, 22, 1123–1130. [Google Scholar] [CrossRef]
- Pavan, B.; Dalpiaz, A. Retinal Pigment Epithelial Cells as a Therapeutic Tool and Target against Retinopathies. Drug Discov. Today 2018, 23, 1672–1679. [Google Scholar] [CrossRef]
- Jovancevic, N.; Khalfaoui, S.; Weinrich, M.; Weidinger, D.; Simon, A.; Kalbe, B.; Kernt, M.; Kampik, A.; Gisselmann, G.; Gelis, L.; et al. Odorant Receptor 51E2 Agonist β-Ionone Regulates RPE Cell Migration and Proliferation. Front. Physiol. 2017, 8, 888. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, T.; Watanabe, T.; Kuramochi-Miyagawa, S.; Takemoto, N.; Shiromoto, Y.; Kudo, A.; Kanai-Azuma, M.; Tashiro, F.; Miyazaki, S.; Katanaya, A.; et al. Mouse GTSF1 Is an Essential Factor for Secondary PiRNA Biogenesis. EMBO Rep. 2018, 19, e42054. [Google Scholar] [CrossRef]
- Fukuda, S.; Varshney, A.; Fowler, B.J.; Wang, S.-B.; Narendran, S.; Ambati, K.; Yasuma, T.; Magagnoli, J.; Leung, H.; Hirahara, S.; et al. Cytoplasmic Synthesis of Endogenous Alu Complementary DNA via Reverse Transcription and Implications in Age-Related Macular Degeneration. Proc. Natl. Acad. Sci. USA 2021, 118, e2022751118. [Google Scholar] [CrossRef] [PubMed]
- Zurdel, J.; Finckh, U.; Menzer, G.; Nitsch, R.M.; Richard, G. CST3 Genotype Associated with Exudative Age Related Macular Degeneration. Br. J. Ophthalmol. 2002, 86, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Crawford, F.C.; Freeman, M.J.; Schinka, J.A.; Abdullah, L.I.; Gold, M.; Hartman, R.; Krivian, K.; Morris, M.D.; Richards, D.; Duara, R.; et al. A Polymorphism in the Cystatin C Gene Is a Novel Risk Factor for Late-Onset Alzheimer’s Disease. Neurology 2000, 55, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.M.; Sharif, U.; Ali, M.; McKibbin, M.; Thompson, J.P.; Gale, R.; Yang, Y.C.; Inglehearn, C.; Paraoan, L. A Missense Variant in CST3 Exerts a Recessive Effect on Susceptibility to Age-Related Macular Degeneration Resembling Its Association with Alzheimer’s Disease. Hum. Genet. 2015, 134, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Carlsson, E.; Supharattanasitthi, W.; Jackson, M.; Paraoan, L. Increased Rate of Retinal Pigment Epithelial Cell Migration and Pro-Angiogenic Potential Ensuing From Reduced Cystatin C Expression. Investig. Ophthalmol. Vis. Sci. 2020, 61, 9. [Google Scholar] [CrossRef]
- Yamamoto, S.; Yabuki, R.; Kitagawa, D. Biophysical and Biochemical Properties of Deup1 Self-Assemblies: A Potential Driver for Deuterosome Formation during Multiciliogenesis. Biol. Open 2021, 10, bio056432. [Google Scholar] [CrossRef]
- Bujakowska, K.M.; Liu, Q.; Pierce, E.A. Photoreceptor Cilia and Retinal Ciliopathies. Cold Spring Harb. Perspect. Biol. 2017, 9, a028274. [Google Scholar] [CrossRef]
- Martinez, G.; Beurois, J.; Dacheux, D.; Cazin, C.; Bidart, M.; Kherraf, Z.-E.; Robinson, D.R.; Satre, V.; Le Gac, G.; Ka, C.; et al. Biallelic Variants in MAATS1 Encoding CFAP91, a Calmodulin-Associated and Spoke-Associated Complex Protein, Cause Severe Astheno-Teratozoospermia and Male Infertility. J. Med. Genet. 2020, 57, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Kurashige, T.; Morino, H.; Matsuda, Y.; Mukai, T.; Murao, T.; Toko, M.; Kume, K.; Ohsawa, R.; Torii, T.; Tokinobu, H.; et al. Retinitis Pigmentosa Prior to Familial ALS Caused by a Homozygous Cilia and Flagella-Associated Protein 410 Mutation. J. Neurol. Neurosurg. Psychiatry 2020, 91, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Santos-Carvalho, A.; Ambrósio, A.F.; Cavadas, C. Neuropeptide Y System in the Retina: From Localization to Function. Prog. Retin. Eye Res. 2015, 47, 19–37. [Google Scholar] [CrossRef]
- Jaakkola, U.; Pesonen, U.; Vainio-Jylhä, E.; Koulu, M.; Pöllönen, M.; Kallio, J. The Leu7Pro Polymorphism of Neuropeptide Y Is Associated with Younger Age of Onset of Type 2 Diabetes Mellitus and Increased Risk for Nephropathy in Subjects with Diabetic Retinopathy. Exp. Clin. Endocrinol. Diabetes Off. J. Ger. Soc. Endocrinol. Ger. Diabetes Assoc. 2006, 114, 147–152. [Google Scholar] [CrossRef]
- Koulu, M.; Movafagh, S.; Tuohimaa, J.; Jaakkola, U.; Kallio, J.; Pesonen, U.; Geng, Y.; Karvonen, M.K.; Vainio-Jylhä, E.; Pöllönen, M.; et al. Neuropeptide Y and Y2-Receptor Are Involved in Development of Diabetic Retinopathy and Retinal Neovascularization. Ann. Med. 2004, 36, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, M.C. Distribution and Function of Neuropeptides W/B Signaling System. Front. Physiol. 2018, 9, 981. [Google Scholar] [CrossRef]
- Singh, G.; Davenport, A.P. Neuropeptide B and W: Neurotransmitters in an Emerging G-Protein-Coupled Receptor System. Br. J. Pharmacol. 2006, 148, 1033–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, W.; Liu, Y.; Gao, Y.; Gong, R.; Wang, W.; Zhang, R.; Wu, Y.; Kang, T.; Wei, D. RMI2 Plays Crucial Roles in Growth and Metastasis of Lung Cancer. Signal Transduct. Target. Ther. 2020, 5, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Nie, S.; Gao, M.; Jiang, Y.; Wan, Y.; Ma, X.; Zhou, S.; Cheng, W. Identification of EPHX2 and RMI2 as Two Novel Key Genes in Cervical Squamous Cell Carcinoma by an Integrated Bioinformatic Analysis. J. Cell. Physiol. 2019, 234, 21260–21273. [Google Scholar] [CrossRef]
- Hudson, D.F.; Amor, D.J.; Boys, A.; Butler, K.; Williams, L.; Zhang, T.; Kalitsis, P. Loss of RMI2 Increases Genome Instability and Causes a Bloom-Like Syndrome. PLoS Genet. 2016, 12, e1006483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.; Lee, H.; Chung, J.; Seo, W.; Kim, G.; Ki, C.; Kim, Y.; Bang, O.Y. Ring Finger Protein 213 Variant and Plaque Characteristics, Vascular Remodeling, and Hemodynamics in Patients With Intracranial Atherosclerotic Stroke: A High-Resolution Magnetic Resonance Imaging and Hemodynamic Study. J. Am. Heart Assoc. 2019, 8, e011996. [Google Scholar] [CrossRef]
- Mapes, J.; Li, Q.; Kannan, A.; Anandan, L.; Laws, M.; Lydon, J.P.; Bagchi, I.C.; Bagchi, M.K. CUZD1 Is a Critical Mediator of the JAK/STAT5 Signaling Pathway That Controls Mammary Gland Development during Pregnancy. PLoS Genet. 2017, 13, e1006654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brink, T.C.; Sudheer, S.; Janke, D.; Jagodzinska, J.; Jung, M.; Adjaye, J. The Origins of Human Embryonic Stem Cells: A Biological Conundrum. Cells Tissues Organs 2008, 188, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Hauser, M.A.; Scott, W.K.; Postel, E.A.; Agarwal, A.; Gallins, P.; Wong, F.; Chen, Y.S.; Spencer, K.; Schnetz-Boutaud, N.; et al. Cigarette Smoking Strongly Modifies the Association of LOC387715 and Age-Related Macular Degeneration. Am. J. Hum. Genet. 2006, 78, 852–864. [Google Scholar] [CrossRef] [Green Version]
- Wortmann, S.B.; Mayr, J.A. Choline-related-inherited metabolic diseases—A mini review. J. Inherit. Metab. Dis. 2019, 42, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Govindarajan, G.; Mathews, S.; Srinivasan, K.; Ramasamy, K.; Periasamy, S. Establishment of Human Retinal Mitoscriptome Gene Expression Signature for Diabetic Retinopathy Using Cadaver Eyes. Mitochondrion 2017, 36, 150–181. [Google Scholar] [CrossRef]
- Kandpal, R.P.; Rajasimha, H.K.; Brooks, M.J.; Nellissery, J.; Wan, J.; Qian, J.; Kern, T.S.; Swaroop, A. Transcriptome Analysis Using next Generation Sequencing Reveals Molecular Signatures of Diabetic Retinopathy and Efficacy of Candidate Drugs. Mol. Vis. 2012, 18, 1123–1146. [Google Scholar]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, Transcriptome and Proteome: The Rise of Omics Data and Their Integration in Biomedical Sciences. Brief. Bioinform. 2018, 19, 286–302. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | LogFC | p-Value | Protein |
|---|---|---|---|
| Fat2 | 4.084 | 3.13 × 10−5 | FAT atypical cadherin 2 |
| Npy4r | 3.518 | 1.10 × 10−4 | Neuropeptide Y receptor Y4-2 |
| C1qtnf2 | 3.248 | 9.57 × 10−4 | C1q And TNF-related 2 |
| Deup1 | 3.133 | 1.81 × 10−4 | Deuterostome assembly protein 1 |
| Ifi204 | 2.881 | 5.62 × 10−3 | Interferon gamma inducible protein |
| Siglec1 | 2.845 | 3.57 × 10−4 | Sialic acid-binding Ig-like lectin 1 |
| Mrap | 2.724 | 1.02 × 10−3 | Melanocortin 2 receptor accessory protein |
| Dmgdh | 2.672 | 2.89 × 10−3 | Dimethylglycine dehydrogenase |
| Maats1 | 2.433 | 8.70 × 10−3 | Cilia and flagella-associated protein 91 |
| Nppb | 2.345 | 4.83 × 10−3 | Natriuretic-peptide B |
| Klf1 | 1.889 | 2.77 × 10−4 | Kruppel-like factor 1 |
| Hba-a2 | 1.867 | 7.19 × 10−3 | Hemoglobin subunit alpha 2 |
| Ppp1r3g | 1.735 | 8.87 × 10−3 | Protein phosphatase 1 regulatory subunit 3G |
| Hbb-bs | 1.378 | 4.73 × 10−3 | Hemoglobin subunit beta |
| Hba-a1 | 1.372 | 1.82 × 10−3 | Hemoglobin subunit alpha 1 |
| Ms4a6b | 1.336 | 4.78 × 10−3 | Membrane spanning 4-domains A6A |
| Gdf2 | 1.133 | 9.95 × 10−3 | Growth differentiation factor 2 |
| Tspan11 | 1.124 | 7.60 × 10−3 | Tetraspanin 11 |
| Nlrp10 | 1.103 | 2.63 × 10−3 | NLR family pyrin domain containing 10 |
| Gene | LogFC | p-Value | Protein |
|---|---|---|---|
| Olfr690 | −3.712 | 8.96 × 10−6 | Olfactory receptor family |
| Gtsf1 | −3.503 | 1.05 × 10−4 | Gametocyte specific factor 1 |
| Tcp10a | −3.121 | 1.77 × 10−3 | T-Complex 10-like 3, pseudogene |
| Olfr460 | −3.077 | 7.60 × 10−4 | Olfactory receptor family |
| Cuzd1 | −2.902 | 4.51 × 10−3 | CUB and zona pellucida-like domains 1 |
| Serpinc1 | −2.721 | 6.30 × 10−4 | Serpin family C member 1 |
| Olfr691 | −2.713 | 1.26 × 10−3 | Olfactory receptor family |
| Opn5 | −2.708 | 8.80 × 10−3 | Opsin 5 |
| Hist1h3i | −2.561 | 5.37 × 10−3 | H3 clustered histone 11 |
| Rmi2 | −2.546 | 6.18 × 10−3 | RecQ mediated genome instability 2 |
| Rnf222 | −2.370 | 6.91 × 10−3 | Ring finger protein 222 |
| Serpinf2 | −2.309 | 1.17 × 10−3 | Serpin family F member 2 |
| Cstl1 | −2.302 | 6.08 × 10−3 | Cystatin-like 1 |
| Npb | −2.000 | 6.35 × 10−3 | Neuropeptide B |
| Anxa9 | −1.746 | 3.87 × 10−4 | Annexin A9 |
| Nanos2 | −1.670 | 2.85 × 10−3 | Nanos C2HC-type zinc finger 2 |
| Tssk4 | −1.603 | 8.17 × 10−3 | Testis specific serine kinase 4 |
| Eqtn | −1.510 | 3.93 × 10−3 | Equatorin |
| Klrg2 | −1.436 | 6.80 × 10−5 | Killer-cell lectin-like receptor G2 |
| Mcemp1 | −1.419 | 3.67 × 10−3 | Mast cell expressed membrane protein 1 |
| Lat | −1.339 | 2.98 × 10−3 | Linker for activation of T-cells |
| Plekha4 | −1.334 | 7.90 × 10−3 | Pleckstrin homology domain containing A4 |
| Misp | −1.270 | 7.52 × 10−3 | Mitotic spindle positioning |
| P4ha3 | −1.176 | 4.91 × 10−3 | Prolyl 4-hydroxylase subunit alpha 3 |
| Gpr84 | −1.175 | 9.61 × 10−4 | G protein-coupled receptor 84 |
| Pi16 | −1.160 | 5.25 × 10−3 | Peptidase inhibitor 16 |
| Slc10a5 | −1.134 | 4.79 × 10−3 | Solute carrier family 10 member 5 |
| Ankrd7 | −1.118 | 7.66 × 10−3 | Ankyrin repeat domain 7 |
| Tmprss5 | −1.109 | 2.09 × 10−3 | Transmembrane serine protease 5 |
| Cdhr3 | −1.091 | 5.62 × 10−3 | Cadherin-related family member 3 |
| Hmga2 | −1.061 | 1.63 × 10−3 | High mobility group AT-hook 2 |
| Sycp2 | −1.061 | 4.34 × 10−3 | Synaptonemal complex protein 2 |
| Cd27 | −1.039 | 5.80 × 10−3 | T-cell activation antigen CD27 |
| Mypn | −1.036 | 6.94 × 10−3 | Myopalladin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dao, D.; Xie, B.; Nadeem, U.; Xiao, J.; Movahedan, A.; D’Souza, M.; Leone, V.; Hariprasad, S.M.; Chang, E.B.; Sulakhe, D.; et al. High-Fat Diet Alters the Retinal Transcriptome in the Absence of Gut Microbiota. Cells 2021, 10, 2119. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10082119
Dao D, Xie B, Nadeem U, Xiao J, Movahedan A, D’Souza M, Leone V, Hariprasad SM, Chang EB, Sulakhe D, et al. High-Fat Diet Alters the Retinal Transcriptome in the Absence of Gut Microbiota. Cells. 2021; 10(8):2119. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10082119
Chicago/Turabian StyleDao, David, Bingqing Xie, Urooba Nadeem, Jason Xiao, Asad Movahedan, Mark D’Souza, Vanessa Leone, Seenu M. Hariprasad, Eugene B. Chang, Dinanath Sulakhe, and et al. 2021. "High-Fat Diet Alters the Retinal Transcriptome in the Absence of Gut Microbiota" Cells 10, no. 8: 2119. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10082119