
Sexually Dimorphic Transcriptomic Changes of Developing Fetal Brain Reveal Signaling Pathways and Marker Genes of Brain Cells in Domestic Pigs


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Fetal Brain Collection
2.2. Transcriptome Profiling by RNA-Seq
2.3. RNA-Seq Data Analysis
2.4. Functional Annotation of Differentially Expressed Genes
2.5. Analysis of Marker Genes of Brain Cells
2.6. Statistical Analyses
3. Results
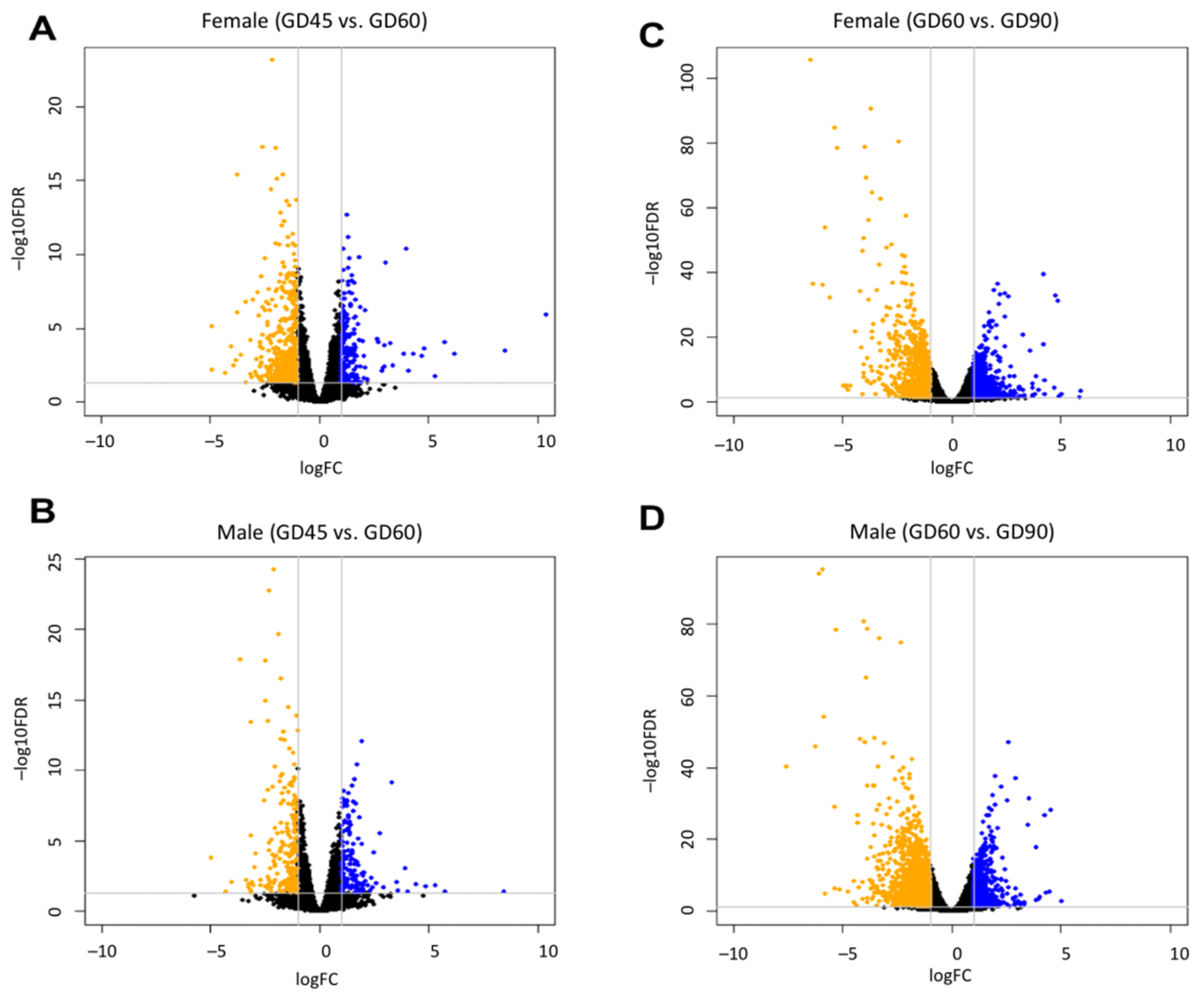
3.1. Developmental Changes in Gene Expression of Fetal Brain
3.2. Influence of Fetal Sex on Gene Expression Changes of Brain
3.3. Effect of Adjacent Fetus on Gene Expression of Brain
3.4. Bias in Exon-Level Expression of Fetal Brain
3.5. Functional Annotation of Differentially Expressed Genes
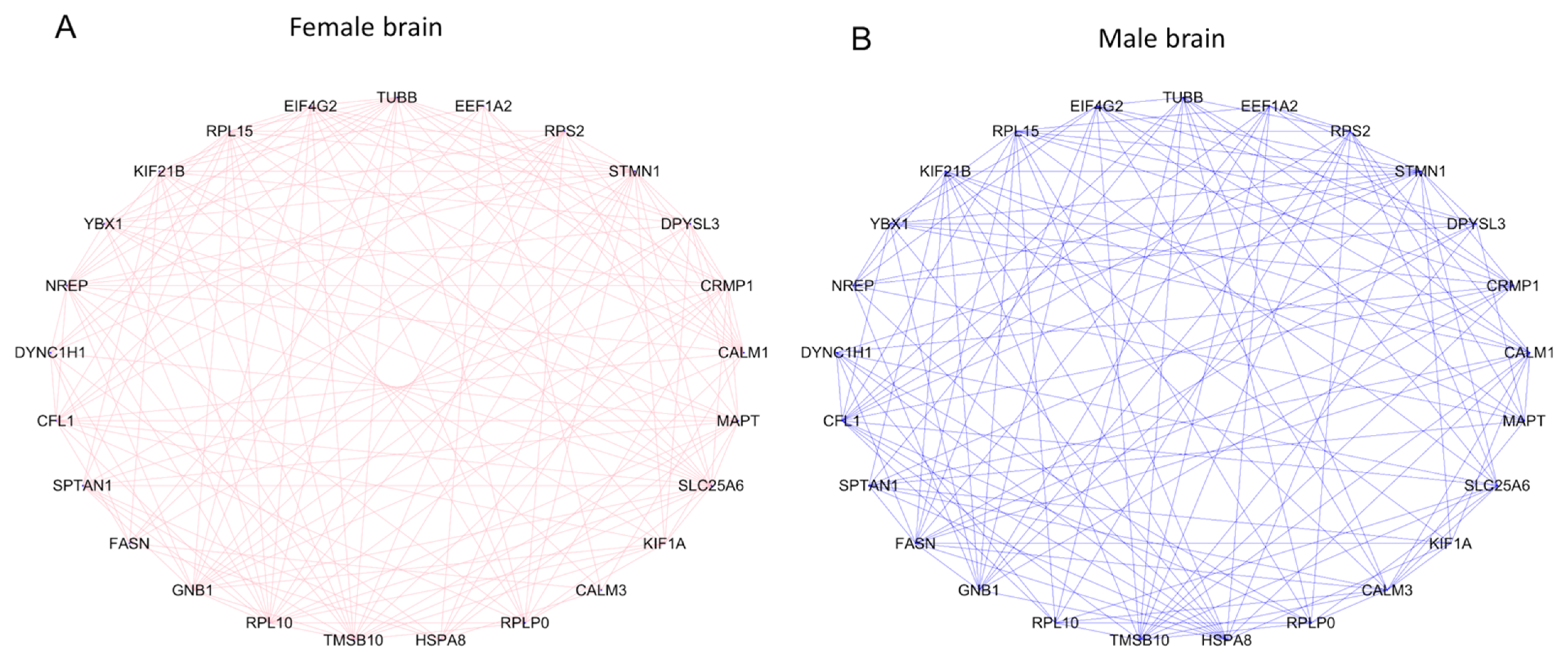
3.6. Identification DE Marker Genes of Brain Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monteagudo, A.; Timor-Tritsch, I.E. Normal sonographic development of the central nervous system from the second trimester onwards using 2D, 3D and transvaginal sonography. Prenat. Diagn. 2008, 29, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, R.S.; Hayes, N.L. CNS development: An overview. Dev. Psychopathol. 1999, 11, 395–417. [Google Scholar] [CrossRef] [PubMed]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106-107, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickerson, J.W.; Dobbing, J. Prenatal and postnatal growth and development of the central nervous system of the pig. Proc. R. Soc. London. Ser. B: Boil. Sci. 1967, 166, 384–395. [Google Scholar] [CrossRef]
- Conrad, M.S.; Dilger, R.N.; Johnson, R.W. Brain Growth of the Domestic Pig (Sus scrofa) from 2 to 24 Weeks of Age: A Longitudinal MRI Study. Dev. Neurosci. 2012, 34, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Groenen, M.A.M.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.-J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef]
- Rothkötter, H.J.; Sowa, E.; Pabst, R. The pig as a model of developmental immunology. Hum. Exp. Toxicol. 2002, 21, 533–536. [Google Scholar] [CrossRef]
- Vodička, P.; Smetana, K.; Dvořánková, B.; Emerick, T.; Xu, Y.Z.; Ourednik, J.; Ourednik, V.; Motlík, J. The Miniature Pig as an Animal Model in Biomedical Research. Ann. N. Y. Acad. Sci. 2005, 1049, 161–171. [Google Scholar] [CrossRef]
- Hoffe, B.; Holahan, M.R. The Use of Pigs as a Translational Model for Studying Neurodegenerative Diseases. Front. Physiol. 2019, 10, 838. [Google Scholar] [CrossRef]
- Kinder, H.; Baker, E.W.; West, F.D. The pig as a preclinical traumatic brain injury model: Current models, functional outcome measures, and translational detection strategies. Neural Regen. Res. 2019, 14, 413. [Google Scholar] [CrossRef]
- Lind, N.M.; Moustgaard, A.; Jelsing, J.; Vajta, G.; Cumming, P.; Hansen, A.K. The use of pigs in neuroscience: Modeling brain disorders. Neurosci. Biobehav. Rev. 2007, 31, 728–751. [Google Scholar] [CrossRef]
- Mudd, A.T.; Dilger, R.N. Early-Life Nutrition and Neurodevelopment: Use of the Piglet as a Translational Model. Adv. Nutr. 2017, 8, 92–104. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.M. Multifaceted origins of sex differences in the brain. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150106. [Google Scholar] [CrossRef]
- McCarthy, M.M. Sex Differences in the Developing Brain as a Source of Inherent Risk. Dialogues Clin. Neurosci. 2016, 18, 361–372. [Google Scholar]
- Crawford, M.A.; Doyle, W.; Meadows, N. Gender differences at birth and differences in fetal growth. Hum. Reprod. 1987, 2, 517–520. [Google Scholar] [CrossRef]
- Raichle, M.E.; Gusnard, D.A. Appraising the brain’s energy budget. Proc. Natl. Acad. Sci. USA 2002, 99, 10237–10239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tekola-Ayele, F.; Workalemahu, T.; Gorfu, G.; Shrestha, D.; Tycko, B.; Wapner, R.; Zhang, C.; Louis, G.M.B. Sex differences in the associations of placental epigenetic aging with fetal growth. Aging 2019, 11, 5412–5432. [Google Scholar] [CrossRef]
- De Lacoste, M.C.; Horvath, D.S.; Woodward, D.J. Possible Sex Differences in the Developing Human Fetal Brain. J. Clin. Exp. Neuropsychol. 1991, 13, 831–846. [Google Scholar] [CrossRef] [PubMed]
- Resko, J.A.; Roselli, C.E. Prenatal Hormones Organize Sex Differences of the Neuroendocrine Reproductive System: Observations on Guinea Pigs and Nonhuman Primates. Cell. Mol. Neurobiol. 1997, 17, 627–648. [Google Scholar] [CrossRef]
- Weinstock, M. Gender Differences in the Effects of Prenatal Stress on Brain Development and Behaviour. Neurochem. Res. 2007, 32, 1730–1740. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L. The Placenta and Neurodevelopment: Sex Differences in Prenatal Vulnerability. Dialogues Clin. Neurosci. 2016, 18, 459–464. [Google Scholar]
- Albert, F.W.; Somel, M.; Carneiro, M.; Aximu-Petri, A.; Halbwax, M.; Thalmann, O.; Blanco-Aguiar, J.A.; Plyusnina, I.Z.; Trut, L.; Villafuerte, R.; et al. A Comparison of Brain Gene Expression Levels in Domesticated and Wild Animals. PLoS Genet. 2012, 8, e1002962. [Google Scholar] [CrossRef] [Green Version]
- Reichert, H. Conserved genetic mechanisms for embryonic brain patterning. Int. J. Dev. Biol. 2002, 46, 81–87. [Google Scholar] [PubMed]
- Saetre, P.; Lindberg, J.; Leonard, J.; Olsson, K.; Pettersson, U.; Ellegren, H.; Bergström, T.; Vilà, C.; Jazin, E. From wild wolf to domestic dog: Gene expression changes in the brain. Mol. Brain Res. 2004, 126, 198–206. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-Sequencing Transcriptome and Splicing Database of Glia, Neurons, and Vascular Cells of the Cerebral Cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-H.; D, D.; Tarn, W.-Y. Alternative Splicing in Neurogenesis and Brain Development. Front. Mol. Biosci. 2018, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Zeltser, L.M.; Leibel, R.L. Roles of the placenta in fetal brain development. Proc. Natl. Acad. Sci. USA 2011, 108, 15667–15668. [Google Scholar] [CrossRef] [Green Version]
- Kratimenos, P.; Penn, A.A. Placental programming of neuropsychiatric disease. Pediatr. Res. 2019, 86, 157–164. [Google Scholar] [CrossRef]
- Behura, S.K.; Kelleher, A.M.; Spencer, T.E. Evidence for functional interactions between the placenta and brain in pregnant mice. FASEB J. 2019, 33, 4261–4272. [Google Scholar] [CrossRef]
- Behura, S.K.; Dhakal, P.; Kelleher, A.M.; Balboula, A.; Patterson, A.; Spencer, T.E. The brain-placental axis: Therapeutic and pharmacological relevancy to pregnancy. Pharmacol. Res. 2019, 149, 104468. [Google Scholar] [CrossRef]
- Blackburn, D.G. Evolution of vertebrate viviparity and specializations for fetal nutrition: A quantitative and qualitative analysis. J. Morphol. 2015, 276, 961–990. [Google Scholar] [CrossRef]
- Ostrovsky, A.N.; Lidgard, S.; Gordon, D.P.; Schwaha, T.; Genikhovich, G.; Ereskovsky, A. Matrotrophy and placentation in invertebrates: A new paradigm. Biol. Rev. 2015, 91, 673–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, R.M.; Green, J.A.; Schulz, L.C. The evolution of the placenta. Reproduction 2016, 152, R179–R189. [Google Scholar] [CrossRef] [Green Version]
- Leiser, R.; Dantzer, V. Structural and functional aspects of porcine placental microvasculature. Brain Struct. Funct. 1988, 177, 409–419. [Google Scholar] [CrossRef]
- Soares, M.J.; Varberg, K.M.; Iqbal, K. Hemochorial placentation: Development, function, and adaptations†. Biol. Reprod. 2018, 99, 196–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- Franzén, O.; Gan, L.-M.; Björkegren, J.L.M. PanglaoDB: A web server for exploration of mouse and human single-cell RNA sequencing data. Database 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [Green Version]
- Steuer, R.; Kurths, J.; Daub, C.; Weise, J.; Selbig, J. The mutual information: Detecting and evaluating dependencies between variables. Bioinformatics 2002, 18, S231–S240. [Google Scholar] [CrossRef] [Green Version]
- Meyer, P.E.; Lafitte, F.; Bontempi, G. minet: A R/Bioconductor Package for Inferring Large Transcriptional Networks Using Mutual Information. BMC Bioinform. 2008, 9, 1–10. [Google Scholar] [CrossRef]
- Wilms, I.; Croux, C. Sparse canonical correlation analysis from a predictive point of view. Biom. J. 2015, 57, 834–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monzón-Sandoval, J.; Castillo-Morales, A.; Urrutia, A.O.; Gutierrez, H. Modular reorganization of the global network of gene regulatory interactions during perinatal human brain development. BMC Dev. Biol. 2016, 16, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goggolidou, P.; Soneji, S.; Powles-Glover, N.; Williams, D.; Sethi, S.; Baban, D.; Simon, M.M.; Ragoussis, I.; Norris, D.P. A chronological expression profile of gene activity during embryonic mouse brain development. Mamm. Genome 2013, 24, 459–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patapoutian, A.; Reichardt, L.F. Roles of Wnt proteins in neural development and maintenance. Curr. Opin. Neurobiol. 2000, 10, 392–399. [Google Scholar] [CrossRef] [Green Version]
- Ryan, B.C.; Vandenbergh, J.G. Intrauterine position effects. Neurosci. Biobehav. Rev. 2002, 26, 665–678. [Google Scholar] [CrossRef]
- Vom Saal, F.S. Se×ual Differentiation in Litter-Bearing Mammals: Influence of Se× of Adjacent Fetuses in Utero. J. Anim. Sci. 1989, 67, 1824–1840. [Google Scholar] [CrossRef] [Green Version]
- Jansson, T.; Myatt, L.; Powell, T. The Role of Trophoblast Nutrient and Ion Transporters in the Development of Pregnancy Complications and Adult Disease. Curr. Vasc. Pharmacol. 2009, 7, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Sangkhae, V.; Nemeth, E. Placental iron transport: The mechanism and regulatory circuits. Free. Radic. Biol. Med. 2019, 133, 254–261. [Google Scholar] [CrossRef]
- Cousins, R.; Wood, C.E. Expression of organic anion transporters 1 and 3 in the ovine fetal brain during the latter half of gestation. Neurosci. Lett. 2010, 484, 22–25. [Google Scholar] [CrossRef] [Green Version]
- Hosoya, K.-I.; Tachikawa, M. Roles of organic anion/cation transporters at the blood–brain and blood–cerebrospinal fluid barriers involving uremic toxins. Clin. Exp. Nephrol. 2011, 15, 478–485. [Google Scholar] [CrossRef]
- Nigam, S.K.; Bush, K.T.; Martovetsky, G.; Ahn, S.-Y.; Liu, H.C.; Richard, E.; Bhatnagar, V.; Wu, W. The Organic Anion Transporter (OAT) Family: A Systems Biology Perspective. Physiol. Rev. 2015, 95, 83–123. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, M.V.; Stallmach, T.; Grundschober, A.F.; Dufour, J.-F.; Serrano, M.A.; Marin, J.J.G.; Sugiyama, Y.; Meier, P.J. Temporal expression profiles of organic anion transport proteins in placenta and fetal liver of the rat. Am. J. Physiol. Integr. Comp. Physiol. 2004, 287, R1505–R1516. [Google Scholar] [CrossRef] [Green Version]
- Sugiyama, D.; Kusuhara, H.; Taniguchi, H.; Ishikawa, S.; Nozaki, Y.; Aburatani, H.; Sugiyama, Y. Functional Characterization of Rat Brain-specific Organic Anion Transporter (Oatp14) at the Blood-Brain Barrier: High Affinity Transporter for Thyroxine. J. Biol. Chem. 2003, 278, 43489–43495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenhouse, C.; Hogg, C.O.; Ashworth, C.J. Associations between fetal size, sex and placental angiogenesis in the pig†. Biol. Reprod. 2019, 100, 239–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, E.M.; Jarvis, S.; Palarea-Albaladejo, J.; Edwards, S.A. The Weaker Sex? The Propensity for Male-Biased Piglet Mortality. PLoS ONE 2012, 7, e30318. [Google Scholar] [CrossRef] [PubMed]
- Dhakal, P.; Kelleher, A.M.; Behura, S.K.; Spencer, T.E. Sexually dimorphic effects of forkhead box a2 (FOXA2) and uterine glands on decidualization and fetoplacental development. Proc. Natl. Acad. Sci. USA 2020, 117, 23952–23959. [Google Scholar] [CrossRef]
- Galjaard, S.; Ameye, L.; Lees, C.C.; Pexsters, A.; Bourne, T.; Timmerman, D.; Devlieger, R. Sex differences in fetal growth and immediate birth outcomes in a low-risk Caucasian population. Biol. Sex Differ. 2019, 10, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampl, M.; Gotsch, F.; Kusanovic, J.P.; Gomez, R.; Nien, J.K.; Frongillo, E.A.; Romero, R. Sex differences in fetal growth responses to maternal height and weight. Am. J. Hum. Biol. 2009, 22, 431–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhakal, P.; Strawn, M.; Samal, A.; Behura, S.K. Fetal Brain Elicits Sexually Conflicting Transcriptional Response to the Ablation of Uterine Forkhead Box A2 (Foxa2) in Mice. Int. J. Mol. Sci. 2021, 22, 9693. [Google Scholar] [CrossRef]
- McPherson, R.L.; Ji, F.; Wu, G.; Blanton, J.R.; Kim, S.W. Growth and compositional changes of fetal tissues in pigs1. J. Anim. Sci. 2004, 82, 2534–2540. [Google Scholar] [CrossRef]
- Ray, S.; Tzeng, R.-Y.; DiCarlo, L.M.; Bundy, J.L.; Vied, C.; Tyson, G.; Nowakowski, R.; Arbeitman, M.N. An Examination of Dynamic Gene Expression Changes in the Mouse Brain During Pregnancy and the Postpartum Period. G3: Genes|Genomes|Genetics 2016, 6, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Holland, P.W.; Takahashi, T. The evolution of homeobox genes: Implications for the study of brain development. Brain Res. Bull. 2005, 66, 484–490. [Google Scholar] [CrossRef]
- Pronker, M.F.; Lemstra, S.; Snijder, J.; Heck, A.J.R.; Thies-Weesie, D.M.E.; Pasterkamp, R.J.; Janssen, B.J.C. Structural basis of myelin-associated glycoprotein adhesion and signalling. Nat. Commun. 2016, 7, 13584. [Google Scholar] [CrossRef] [PubMed]
- Brockschnieder, D.; Sabanay, H.; Riethmacher, D.; Peles, E. Ermin, A Myelinating Oligodendrocyte-Specific Protein That Regulates Cell Morphology. J. Neurosci. 2006, 26, 757–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde Parfet, K.A.; Lamberson, W.R.; Rieke, A.R.; Cantley, T.C.; Ganjam, V.K.; vom Saal, F.S.; Day, B.N. Intrauterine Posi-tion Effects in Male and Female Swine: Subsequent Survivability, Growth Rate, Morphology and Semen Characteristics. J. Anim. Sci. 1990, 68, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Jang, Y.D.; Ma, Y.L.; Lindemann, M.D. Intrauterine position affects fetal weight and crown-rump length throughout gestation1. J. Anim. Sci. 2014, 92, 4400–4406. [Google Scholar] [CrossRef] [Green Version]
- Sah, V.P.; Attardi, L.D.; Mulligan, G.J.; Williams, B.; Bronson, R.T.; Jacks, T. A subset of p53-deficient embryos exhibit exencephaly. Nat. Genet. 1995, 10, 175–180. [Google Scholar] [CrossRef]
- Delbridge, A.R.; Kueh, A.J.; Ke, F.; Zamudio, N.M.; El-Saafin, F.; Jansz, N.; Wang, G.-Y.; Iminitoff, M.; Beck, T.; Haupt, S.; et al. Loss of p53 Causes Stochastic Aberrant X-Chromosome Inactivation and Female-Specific Neural Tube Defects. Cell Rep. 2019, 27, 442–454.e5. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, J.; Kaufman, M.H.; Harrison, D.J.; Clarke, A. High-frequency developmental abnormalities in p53-deficient mice. Curr. Biol. 1995, 5, 931–936. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Tian, Y.; Zhang, H.; Qu, L.; Chen, Y.; Liu, Q.; Luo, Y.; Wu, X. p53 Mutant p53N236S Induces Neural Tube Defects in Female Embryos. Int. J. Biol. Sci. 2019, 15, 2006–2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickens, M.M.; Bangasser, D.A.; Briand, L.A. Sex Differences in Psychiatric Disease: A Focus on the Glutamate System. Front. Mol. Neurosci. 2018, 11, 197. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ma, Y.; Hu, J.; Cheng, W.; Jiang, H.; Zhang, X.; Li, M.; Ren, J.; Li, X. Prenatal chronic mild stress induces depression-like behavior and sex-specific changes in regional glutamate receptor expression patterns in adult rats. Neuroscience 2015, 301, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, D.; Dikranian, K.; Ishimaru, M.; Nardi, A.; Corso, T.; Tenkova, T.; Olney, J.; Fix, A. Disseminated Corticolimbic Neuronal Degeneration Induced in Rat Brain by MK-801: Potential Relevance to Alzheimer’s Disease. Neurobiol. Dis. 1998, 5, 305–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougall, S.A.; Moran, A.E.; Baum, T.J.; Apodaca, M.G.; Real, V. Effects of ketamine on the unconditioned and conditioned locomotor activity of preadolescent and adolescent rats: Impact of age, sex, and drug dose. Psychopharmacology 2017, 234, 2683–2696. [Google Scholar] [CrossRef]
- Carthy, E.; Ellender, T. Histamine, Neuroinflammation and Neurodevelopment: A Review. Front. Neurosci. 2021, 15, 870. [Google Scholar] [CrossRef] [PubMed]
- Kalish, B.T.; Kim, E.; Finander, B.; Duffy, E.E.; Kim, H.; Gilman, C.K.; Yim, Y.S.; Tong, L.; Kaufman, R.J.; Griffith, E.C.; et al. Maternal immune activation in mice disrupts proteostasis in the fetal brain. Nat. Neurosci. 2021, 24, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Bilgüvar, K.; Ishigame, K.; Sestan, N.; Günel, M.; Louvi, A. Functional Synergy between Cholecystokinin Receptors CCKAR and CCKBR in Mammalian Brain Development. PLoS ONE 2015, 10, e0124295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshibu, K.; Levitt, P. Sex differences in expression of transforming growth factor-α and epidermal growth factor receptor mRNA in waved-1 and C57Bl6 mice. Neuroscience 2005, 134, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.J.; Pohl, S.Ö.-G.; Deshmukh, A.; Visweswaran, M.; Ward, N.C.; Arfuso, F.; Agostino, M.; Dharmarajan, A. The Role of Wnt Signalling in Angiogenesis. Clin. Biochem. Rev. 2017, 38, 131–142. [Google Scholar] [PubMed]
- Wray, S. From Nose to Brain: Development of Gonadotropin-Releasing Hormone−1 Neurons. J. Neuroendocrinol. 2010, 22, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Gulati, A. Endothelin Receptors, Mitochondria and Neurogenesis in Cerebral Ischemia. Curr. Neuropharmacol. 2016, 14, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, J.B.; Beyer, C.; Hutchison, R.E.; Wozniak, A. Sexual dimorphism in the developmental regulation of brain aromatase. J. Steroid Biochem. Mol. Biol. 1995, 53, 307–313. [Google Scholar] [CrossRef]
- Wheelock, M.; Hect, J.; Hernandez-Andrade, E.; Hassan, S.; Romero, R.; Eggebrecht, A.; Thomason, M. Sex differences in functional connectivity during fetal brain development. Dev. Cogn. Neurosci. 2019, 36, 100632. [Google Scholar] [CrossRef] [PubMed]
- Charvet, C.J.; Striedter, G.F.; Finlay, B.L. Evo-Devo and Brain Scaling: Candidate Developmental Mechanisms for Variation and Constancy in Vertebrate Brain Evolution. Brain Behav. Evol. 2011, 78, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Feltes, B.C.; Poloni, J.D.F.; Bonatto, D. Development and Aging: Two Opposite but Complementary Phenomena. Aging 2014, 40, 74–84. [Google Scholar] [CrossRef]
- Goyal, M.S.; Blazey, T.M.; Su, Y.; Couture, L.E.; Durbin, T.J.; Bateman, R.J.; Benzinger, T.L.-S.; Morris, J.C.; Raichle, M.E.; Vlassenko, A.G. Persistent metabolic youth in the aging female brain. Proc. Natl. Acad. Sci. USA 2019, 116, 3251–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | Number of DEGs | Differential Expression |
|---|---|---|
| F.45 vs. F.60 | 801 | Upregulated |
| F.45 vs. F.60 | 1258 | Downregulated |
| F.60 vs. F.90 | 2665 | Upregulated |
| F.60 vs. F.90 | 2660 | Downregulated |
| M.45 vs. M.60 | 571 | Upregulated |
| M.45 vs. M.60 | 752 | Downregulated |
| M.60 vs. M.90 | 3163 | Upregulated |
| M.60 vs. M.90 | 3442 | Downregulated |
| Female (GD45 vs. GD60) | Female (GD60 vs. GD90) | Male (GD45 vs. GD60) | Male (GD60 vs. GD90) | |
|---|---|---|---|---|
| Downregulated | 132 (99) | 694 (491) | 36 (29) | 274 (215) |
| Upregulated | 205 (148) | 268 (200) | 14 (12) | 1772 (1222) |
| Total | 337 (247) | 962 (691) | 50 (41) | 2046 (1437) |
| Pathway | Number of DEGs | Fold Enrichment | FDR | Fetal Se× | Gestation Days |
|---|---|---|---|---|---|
| Ionotropic glutamate receptor pathway (P00037) | 15 | 3.32 | 7.4 × 10−3 | F | 45 vs. 60 |
| p53 pathway (P00059) | 20 | 2.49 | 1.9 × 10−2 | F | 45 vs. 60 |
| Endothelin signaling pathway (P00019) | 19 | 2.47 | 2.3 × 10−2 | F | 45 vs. 60 |
| Gonadotropin−releasing hormone receptor pathway (P06664) | 48 | 2.3 | 1.3 × 10−3 | F | 45 vs. 60 |
| Angiogenesis (P00005) | 35 | 2.21 | 3.9 × 10−3 | F | 45 vs. 60 |
| Wnt signaling pathway (P00057) | 52 | 1.81 | 7.4 × 10−3 | F | 45 vs. 60 |
| Gonadotropin−releasing hormone receptor pathway (P06664) | 99 | 1.82 | 1.6 × 10−4 | F | 60 vs. 90 |
| Angiogenesis (P00005) | 68 | 1.65 | 4.0 × 10−2 | F | 60 vs. 90 |
| Wnt signaling pathway (P00057) | 30 | 1.99 | 3.4 × 10−2 | M | 45 vs. 60 |
| Histamine H1 receptor mediated signaling pathway (P04385) | 27 | 2.13 | 4.8 × 10−2 | M | 60 vs. 90 |
| Endothelin signaling pathway (P00019) | 48 | 1.96 | 1.1 × 10−2 | M | 60 vs. 90 |
| Metabotropic glutamate receptor group III pathway (P00039) | 39 | 1.93 | 4.0 × 10−2 | M | 60 vs. 90 |
| Heterotrimeric G−protein signaling pathway−Gq alpha and Go alpha mediated pathway (P00027) | 67 | 1.88 | 3.0 × 10−3 | M | 60 vs. 90 |
| Angiogenesis (P00005) | 88 | 1.75 | 3.2 × 10−3 | M | 60 vs. 90 |
| Heterotrimeric G−protein signaling pathway−Gi alpha and Gs alpha mediated pathway (P00026) | 81 | 1.69 | 6.5 × 10−3 | M | 60 vs. 90 |
| EGF receptor signaling pathway (P00018) | 65 | 1.6 | 5.0 × 10−2 | M | 60 vs. 90 |
| CCKR signaling map (P06959) | 76 | 1.53 | 4.5 × 10−2 | M | 60 vs. 90 |
| Gonadotropin−releasing hormone receptor pathway (P06664) | 102 | 1.53 | 1.2 × 10−2 | M | 60 vs. 90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strawn, M.; Moraes, J.G.N.; Safranski, T.J.; Behura, S.K. Sexually Dimorphic Transcriptomic Changes of Developing Fetal Brain Reveal Signaling Pathways and Marker Genes of Brain Cells in Domestic Pigs. Cells 2021, 10, 2439. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092439
Strawn M, Moraes JGN, Safranski TJ, Behura SK. Sexually Dimorphic Transcriptomic Changes of Developing Fetal Brain Reveal Signaling Pathways and Marker Genes of Brain Cells in Domestic Pigs. Cells. 2021; 10(9):2439. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092439
Chicago/Turabian StyleStrawn, Monica, Joao G. N. Moraes, Timothy J. Safranski, and Susanta K. Behura. 2021. "Sexually Dimorphic Transcriptomic Changes of Developing Fetal Brain Reveal Signaling Pathways and Marker Genes of Brain Cells in Domestic Pigs" Cells 10, no. 9: 2439. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10092439