
Citalopram Neuroendocrine Challenge Shows Altered Tryptophan and Kynurenine Metabolism in Migraine
 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants
2.3. Clinical Variables of Migraine
2.4. Ethics
2.5. Biological Samples
2.6. Statistical Methods
3. Results
3.1. Descriptive Characteristics of the Participants
3.2. Baseline Plasma Concentrations
3.3. Citalopram Neuroendocrine Challenge
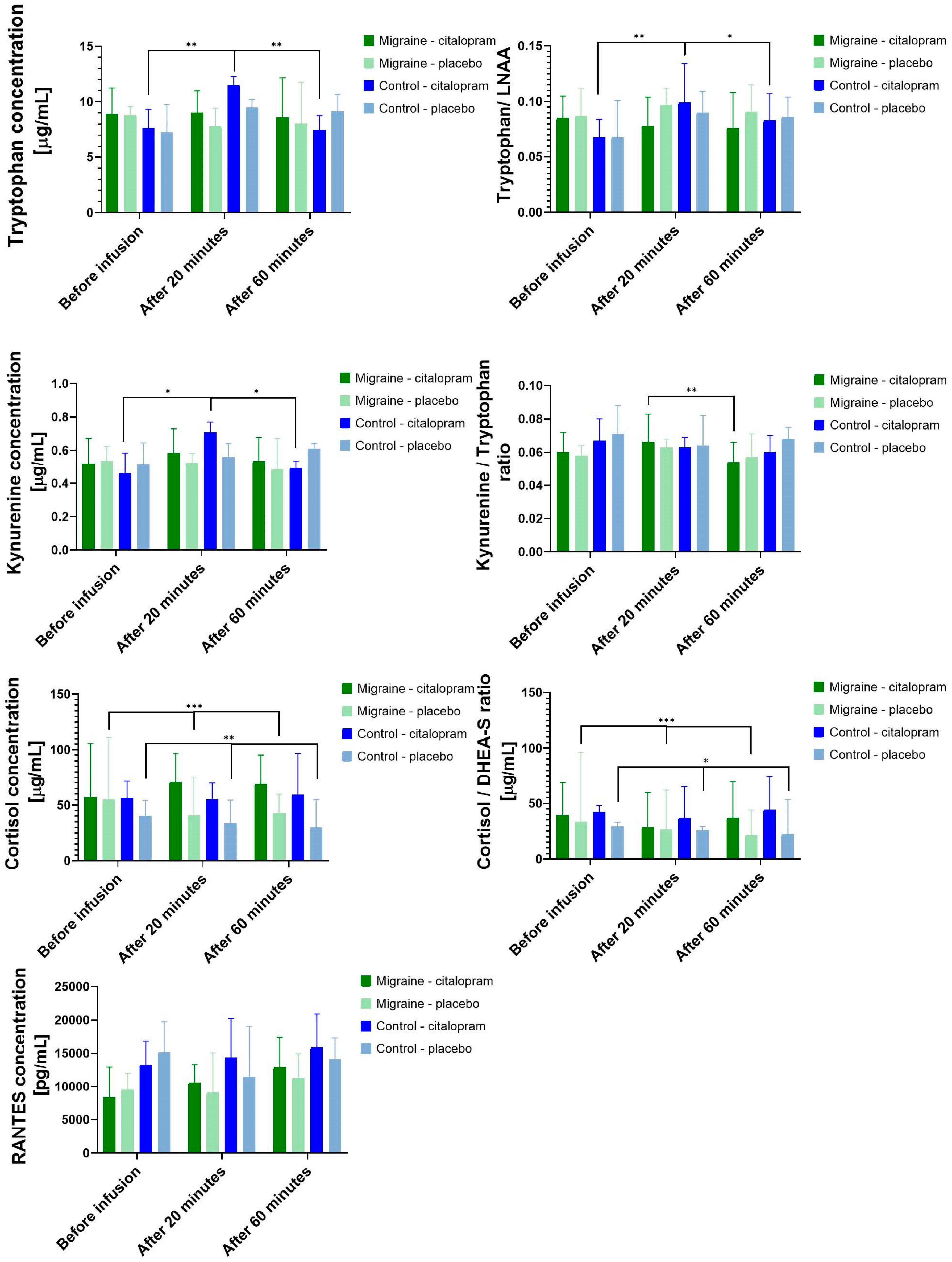
3.3.1. Main Effect of Citalopram Neuroendocrine Challenge
3.3.2. Differences between the Effects of Citalopram Neuroendocrine Challenge in Migraine Patients and Controls
3.4. The Relationship of Migraine Parameters with the Measured Biomarkers
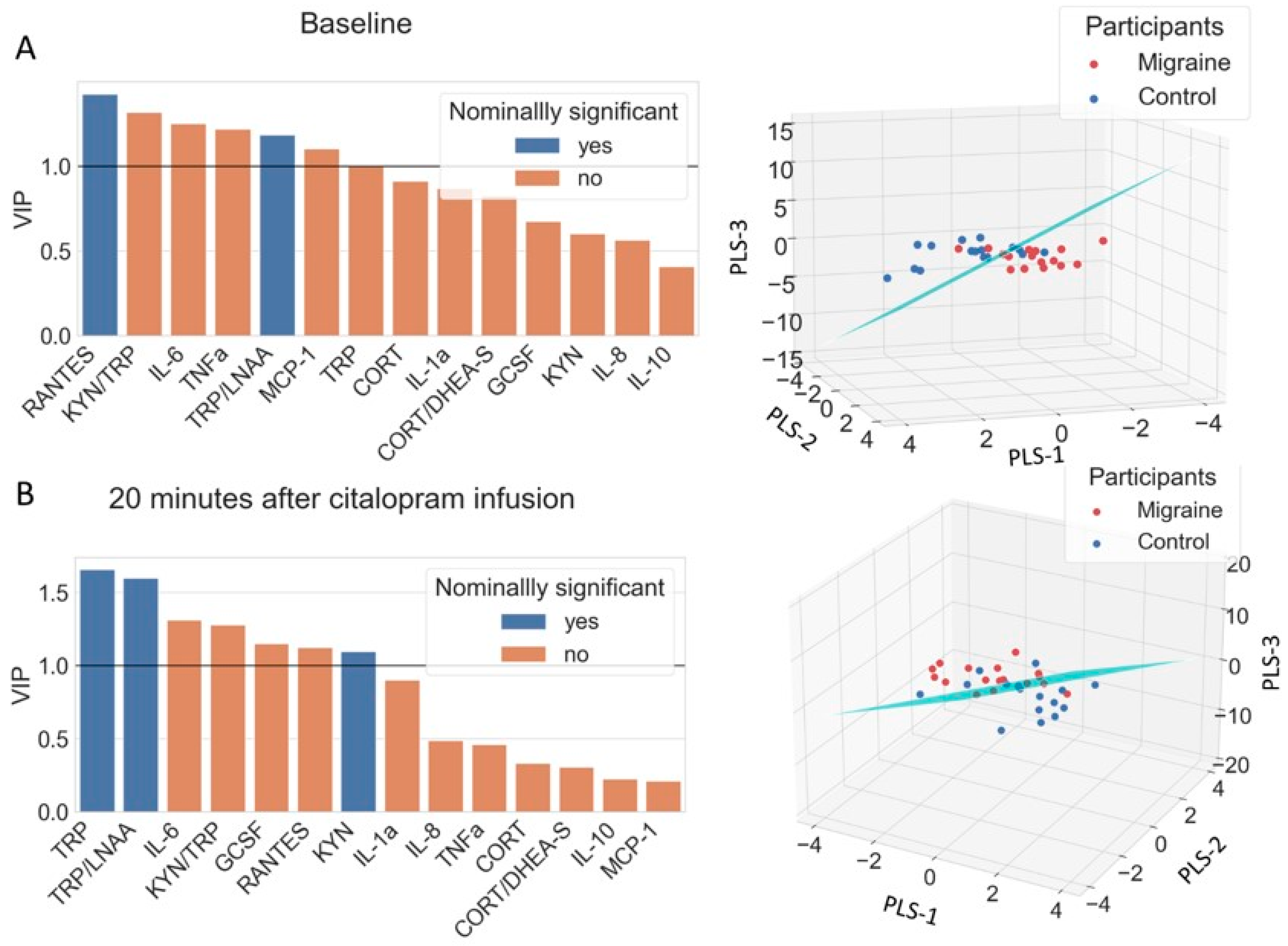
3.5. Discriminating Migraine Patients from Controls with PLS-LDA
4. Discussion
4.1. Alterations of the TRP Metabolism in Migraine
4.2. Migraine Patients Failed to Activate TRP Pathway during Citalopram Neuroendocrine Challenge
4.3. Inflammatory Biomarkers in Migraine
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD DALYs; HALE Collaborators. Global, regional, and national disability-adjusted life-years (DALYs) for 359 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1859–1922. [Google Scholar] [CrossRef] [Green Version]
- Goadsby, P.J.; Holland, P.R.; Martins-Oliveira, M.; Hoffmann, J.; Schankin, C.; Akerman, S. Pathophysiology of Migraine: A Disorder of Sensory Processing. Physiol. Rev. 2017, 97, 553–622. [Google Scholar] [CrossRef] [PubMed]
- Sicuteri, F.; Testi, A.; Anselmi, B. Biochemical Investigations in Headache: Increase in the Hydroxyindoleacetic Acid Excretion During Migraine Attacks. Int. Arch. Allergy Immunol. 1961, 19, 55–58. [Google Scholar] [CrossRef]
- Ferrari, M.D.; Odink, J.; Tapparelli, C.; Van Kempen, G.M.; Pennings, E.J.; Bruyn, G.W. Serotonin metabolism in migraine. Neurology 1989, 39, 1239–1242. [Google Scholar] [CrossRef]
- Panconesi, A. Serotonin and migraine: A reconsideration of the central theory. J. Headache Pain 2008, 9, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Hoskin, K.L.; Kaube, H.; Goadsby, P.J. Sumatriptan can inhibit trigeminal afferents by an exclusively neural mechanism. Brain 1996, 119 Pt 5, 1419–1428. [Google Scholar] [CrossRef] [Green Version]
- Cameron, C.; Kelly, S.; Hsieh, S.C.; Murphy, M.; Chen, L.; Kotb, A.; Peterson, J.; Coyle, D.; Skidmore, B.; Gomes, T.; et al. Triptans in the Acute Treatment of Migraine: A Systematic Review and Network Meta-Analysis. Headache 2015, 55 (Suppl. 4), 221–235. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.W.; Lai, P.Y.; Chen, Y.L.; Wang, Y.F.; Lirng, J.F.; Chen, S.T.; Lai, K.L.; Chen, W.T.; Wu, Y.T.; Wang, S.J. The Use of Neuroimaging for Predicting Sumatriptan Treatment Response in Patients With Migraine. Front. Neurol. 2022, 13, 798695. [Google Scholar] [CrossRef]
- de Vries, T.; Villalon, C.M.; MaassenVanDenBrink, A. Pharmacological treatment of migraine: CGRP and 5-HT beyond the triptans. Pharmacol. Ther. 2020, 211, 107528. [Google Scholar] [CrossRef]
- Palego, L.; Betti, L.; Rossi, A.; Giannaccini, G. Tryptophan Biochemistry: Structural, Nutritional, Metabolic, and Medical Aspects in Humans. J. Amino Acids 2016, 2016, 8952520. [Google Scholar] [CrossRef] [Green Version]
- Fernstrom, J.D. Large neutral amino acids: Dietary effects on brain neurochemistry and function. Amino Acids 2013, 45, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Alam, Z.; Coombes, N.; Waring, R.H.; Williams, A.C.; Steventon, G.B. Plasma levels of neuroexcitatory amino acids in patients with migraine or tension headache. J. Neurol. Sci. 1998, 156, 102–106. [Google Scholar] [CrossRef]
- Salmon, S.; Fanciullacci, M.; Bonciani, M.; Sicuteri, F. Plasma tryptophan in migraine. Headache 1978, 17, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Curto, M.; Lionetto, L.; Negro, A.; Capi, M.; Fazio, F.; Giamberardino, M.A.; Simmaco, M.; Nicoletti, F.; Martelletti, P. Altered kynurenine pathway metabolites in serum of chronic migraine patients. J. Headache Pain 2015, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- Gecse, K.; Dobos, D.; Aranyi, C.S.; Galambos, A.; Baksa, D.; Kocsel, N.; Szabo, E.; Pap, D.; Virag, D.; Ludanyi, K.; et al. Association of plasma tryptophan concentration with periaqueductal gray matter functional connectivity in migraine patients. Sci. Rep. 2022, 12, 739. [Google Scholar] [CrossRef]
- Ren, C.; Liu, J.; Zhou, J.; Liang, H.; Wang, Y.; Sun, Y.; Ma, B.; Yin, Y. Low levels of serum serotonin and amino acids identified in migraine patients. Biochem. Biophys. Res. Commun. 2018, 496, 267–273. [Google Scholar] [CrossRef]
- Tuka, B.; Nyari, A.; Cseh, E.K.; Kortesi, T.; Vereb, D.; Tomosi, F.; Kecskemeti, G.; Janaky, T.; Tajti, J.; Vecsei, L. Clinical relevance of depressed kynurenine pathway in episodic migraine patients: Potential prognostic markers in the peripheral plasma during the interictal period. J. Headache Pain 2021, 22, 60. [Google Scholar] [CrossRef]
- Drummond, P.D. Tryptophan depletion increases nausea, headache and photophobia in migraine sufferers. Cephalalgia 2006, 26, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Razeghi Jahromi, S.; Togha, M.; Ghorbani, Z.; Hekmatdoost, A.; Khorsha, F.; Rafiee, P.; Shirani, P.; Nourmohammadi, M.; Ansari, H. The association between dietary tryptophan intake and migraine. Neurol. Sci. 2019, 40, 2349–2355. [Google Scholar] [CrossRef]
- Kangasniemi, P.; Falck, B.; Långvik, V.A.; Hyyppä, M.T. Levotryptophan treatment in migraine. Headache 1978, 18, 161–165. [Google Scholar] [CrossRef]
- Hasselmark, L.; Malmgren, R.; Hannerz, J. Effect of a carbohydrate-rich diet, low in protein-tryptophan, in classic and common migraine. Cephalalgia 1987, 7, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Fejes, A.; Párdutz, A.; Toldi, J.; Vécsei, L. Kynurenine metabolites and migraine: Experimental studies and therapeutic perspectives. Curr. Neuropharmacol. 2011, 9, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecsei, L.; Szalardy, L.; Fulop, F.; Toldi, J. Kynurenines in the CNS: Recent advances and new questions. Nat. Rev. Drug Discov. 2013, 12, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Vikelis, M.; Mitsikostas, D.D. The role of glutamate and its receptors in migraine. CNS Neurol. Disord. Drug Targets 2007, 6, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Vamos, E.; Pardutz, A.; Varga, H.; Bohar, Z.; Tajti, J.; Fulop, F.; Toldi, J.; Vecsei, L. l-kynurenine combined with probenecid and the novel synthetic kynurenic acid derivative attenuate nitroglycerin-induced nNOS in the rat caudal trigeminal nucleus. Neuropharmacology 2009, 57, 425–429. [Google Scholar] [CrossRef]
- Greco, R.; Demartini, C.; Zanaboni, A.M.; Redavide, E.; Pampalone, S.; Toldi, J.; Fulop, F.; Blandini, F.; Nappi, G.; Sandrini, G.; et al. Effects of kynurenic acid analogue 1 (KYNA-A1) in nitroglycerin-induced hyperalgesia: Targets and anti-migraine mechanisms. Cephalalgia 2017, 37, 1272–1284. [Google Scholar] [CrossRef]
- Badawy, A.A.; Guillemin, G. The Plasma [Kynurenine]/[Tryptophan] Ratio and Indoleamine 2,3-Dioxygenase: Time for Appraisal. Int. J. Tryptophan Res. 2019, 12, 1178646919868978. [Google Scholar] [CrossRef] [Green Version]
- Mithaiwala, M.N.; Santana-Coelho, D.; Porter, G.A.; O’Connor, J.C. Neuroinflammation and the Kynurenine Pathway in CNS Disease: Molecular Mechanisms and Therapeutic Implications. Cells 2021, 10, 1548. [Google Scholar] [CrossRef]
- Wu, H.; Denna, T.H.; Storkersen, J.N.; Gerriets, V.A. Beyond a neurotransmitter: The role of serotonin in inflammation and immunity. Pharmacol. Res. 2019, 140, 100–114. [Google Scholar] [CrossRef]
- Zang, X.; Zheng, X.; Hou, Y.; Hu, M.; Wang, H.; Bao, X.; Zhou, F.; Wang, G.; Hao, H. Regulation of proinflammatory monocyte activation by the kynurenine-AhR axis underlies immunometabolic control of depressive behavior in mice. FASEB J. 2018, 32, 1944–1956. [Google Scholar] [CrossRef] [Green Version]
- Walker, F.R. A critical review of the mechanism of action for the selective serotonin reuptake inhibitors: Do these drugs possess anti-inflammatory properties and how relevant is this in the treatment of depression? Neuropharmacology 2013, 67, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K. Does inflammation have a role in migraine? Nat. Rev. Neurol. 2019, 15, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Spekker, E.; Tanaka, M.; Szabo, A.; Vecsei, L. Neurogenic Inflammation: The Participant in Migraine and Recent Advancements in Translational Research. Biomedicines 2021, 10, 76. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, G.; Zsombok, T.; Jakab, B.; Nemeth, J.; Szolcsanyi, J.; Bagdy, G. Sumatriptan causes parallel decrease in plasma calcitonin gene-related peptide (CGRP) concentration and migraine headache during nitroglycerin induced migraine attack. Cephalalgia 2005, 25, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, G.; Zsombok, T.; Modos, E.A.; Olajos, S.; Jakab, B.; Nemeth, J.; Szolcsanyi, J.; Vitrai, J.; Bagdy, G. NO-induced migraine attack: Strong increase in plasma calcitonin gene-related peptide (CGRP) concentration and negative correlation with platelet serotonin release. Pain 2003, 106, 461–470. [Google Scholar] [CrossRef]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K.; Krause, D.N. CGRP as the target of new migraine therapies-successful translation from bench to clinic. Nat. Rev. Neurol. 2018, 14, 338–350. [Google Scholar] [CrossRef]
- Moskowitz, M.A. Defining a pathway to discovery from bench to bedside: The trigeminovascular system and sensitization. Headache 2008, 48, 688–690. [Google Scholar] [CrossRef]
- Afroz, S.; Arakaki, R.; Iwasa, T.; Oshima, M.; Hosoki, M.; Inoue, M.; Baba, O.; Okayama, Y.; Matsuka, Y. CGRP Induces Differential Regulation of Cytokines from Satellite Glial Cells in Trigeminal Ganglia and Orofacial Nociception. Int. J. Mol. Sci. 2019, 20, 711. [Google Scholar] [CrossRef] [Green Version]
- Empl, M.; Sostak, P.; Riedel, M.; Schwarz, M.; Muller, N.; Forderreuther, S.; Straube, A. Decreased sTNF-RI in migraine patients? Cephalalgia 2003, 23, 55–58. [Google Scholar] [CrossRef]
- Christopherson, K., II; Hromas, R. Chemokine regulation of normal and pathologic immune responses. Stem Cells 2001, 19, 388–396. [Google Scholar] [CrossRef]
- Suneson, K.; Lindahl, J.; Chamli Harsmar, S.; Soderberg, G.; Lindqvist, D. Inflammatory Depression-Mechanisms and Non-Pharmacological Interventions. Int. J. Mol. Sci. 2021, 22, 1640. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.; Yang, Z.; Xu, P.; Zhang, H. Aberrations in peripheral inflammatory cytokine levels in migraine: A systematic review and meta-analysis. J. Clin. Neurosci. 2022, 98, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Lotrich, F.E.; Bies, R.; Muldoon, M.F.; Manuck, S.B.; Smith, G.S.; Pollock, B.G. Neuroendocrine response to intravenous citalopram in healthy control subjects: Pharmacokinetic influences. Psychopharmacology 2005, 178, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Seifritz, E.; Baumann, P.; Muller, M.J.; Annen, O.; Amey, M.; Hemmeter, U.; Hatzinger, M.; Chardon, F.; Holsboer-Trachsler, E. Neuroendocrine effects of a 20-mg citalopram infusion in healthy males. A placebo-controlled evaluation of citalopram as 5-HT function probe. Neuropsychopharmacology 1996, 14, 253–263. [Google Scholar] [CrossRef] [Green Version]
- Edes, A.E.; McKie, S.; Szabo, E.; Kokonyei, G.; Pap, D.; Zsombok, T.; Hullam, G.; Gonda, X.; Kozak, L.R.; McFarquhar, M.; et al. Spatiotemporal brain activation pattern following acute citalopram challenge is dose dependent and associated with neuroticism: A human phMRI study. Neuropharmacology 2020, 170, 107807. [Google Scholar] [CrossRef] [Green Version]
- Bhagwagar, Z.; Hafizi, S.; Cowen, P.J. Acute citalopram administration produces correlated increases in plasma and salivary cortisol. Psychopharmacology 2002, 163, 118–120. [Google Scholar] [CrossRef]
- Attenburrow, M.J.; Mitter, P.R.; Whale, R.; Terao, T.; Cowen, P.J. Low-dose citalopram as a 5-HT neuroendocrine probe. Psychopharmacology 2001, 155, 323–326. [Google Scholar] [CrossRef]
- Edes, A.E.; McKie, S.; Szabo, E.; Kokonyei, G.; Pap, D.; Zsombok, T.; Magyar, M.; Csepany, E.; Hullam, G.; Szabo, A.G.; et al. Increased activation of the pregenual anterior cingulate cortex to citalopram challenge in migraine: An fMRI study. BMC Neurol. 2019, 19, 237. [Google Scholar] [CrossRef] [Green Version]
- Laudenslager, M.L.; Calderone, J.; Philips, S.; Natvig, C.; Carlson, N.E. Diurnal patterns of salivary cortisol and DHEA using a novel collection device: Electronic monitoring confirms accurate recording of collection time using this device. Psychoneuroendocrinology 2013, 38, 1596–1606. [Google Scholar] [CrossRef] [Green Version]
- Weitzman, E.D.; Fukushima, D.; Nogeire, C.; Roffwarg, H.; Gallagher, T.F.; Hellman, L. Twenty-four hour pattern of the episodic secretion of cortisol in normal subjects. J. Clin. Endocrinol. Metab. 1971, 33, 14–22. [Google Scholar] [CrossRef]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (M.I.N.I.): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry 1998, 59 (Suppl. 20), 22–33. [Google Scholar] [PubMed]
- Arnold, M. Headache Classification Committee of the International Headache Society (IHS) The International Classification of Headache Disorders. Cephalalgia 2018, 38, 1–211. [Google Scholar] [CrossRef] [PubMed]
- Virág, D.; Király, M.; Drahos, L.; Édes, A.E.; Gecse, K.; Bagdy, G.; Juhász, G.; Antal, I.; Klebovich, I.; Dalmadi Kiss, B.; et al. Development, validation and application of LC-MS/MS method for quantification of amino acids, kynurenine and serotonin in human plasma. J. Pharm. Biomed. Anal. 2020, 180, 113018. [Google Scholar] [CrossRef] [PubMed]
- Kokonyei, G.; Galambos, A.; Kocsel, N.; Szabo, E.; Edes, A.E.; Gecse, K.; Baksa, D.; Pap, D.; Kozak, L.R.; Bagdy, G.; et al. Inter-individual differences in pain anticipation and pain perception in migraine: Neural correlates of migraine frequency and cortisol-to-dehydroepiandrosterone sulfate (DHEA-S) ratio. PLoS ONE 2021, 16, e0261570. [Google Scholar] [CrossRef] [PubMed]
- Pyrillou, K.; Burzynski, L.C.; Clarke, M.C.H. Alternative Pathways of IL-1 Activation, and Its Role in Health and Disease. Front. Immunol. 2020, 11, 613170. [Google Scholar] [CrossRef]
- Panopoulos, A.D.; Watowich, S.S. Granulocyte colony-stimulating factor: Molecular mechanisms of action during steady state and ‘emergency’ hematopoiesis. Cytokine 2008, 42, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Tóth, K.; Lénárt, N.; Berki, P.; Fekete, R.; Szabadits, E.; Pósfai, B.; Cserép, C.; Alatshan, A.; Benkő, S.; Kiss, D.; et al. The NKCC1 ion transporter modulates microglial phenotype and inflammatory response to brain injury in a cell-autonomous manner. PLoS Biol. 2022, 20, e3001526. [Google Scholar] [CrossRef]
- Kamin, H.S.; Kertes, D.A. Cortisol and DHEA in development and psychopathology. Horm. Behav. 2017, 89, 69–85. [Google Scholar] [CrossRef]
- Anderson, I.M.; Juhasz, G.; Thomas, E.; Downey, D.; McKie, S.; Deakin, J.F.; Elliott, R. The effect of acute citalopram on face emotion processing in remitted depression: A pharmacoMRI study. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2011, 21, 140–148. [Google Scholar] [CrossRef]
- Del-Ben, C.M.; Deakin, J.F.W.; McKie, S.; Delvai, N.A.; Williams, S.R.; Elliott, R.; Dolan, M.; Anderson, I.M. The Effect of Citalopram Pretreatment on Neuronal Responses to Neuropsychological Tasks in Normal Volunteers: An fMRI Study. Neuropsychopharmacology 2005, 30, 1724–1734. [Google Scholar] [CrossRef] [Green Version]
- Klomp, A.; van Wingen, G.A.; de Ruiter, M.B.; Caan, M.W.A.; Denys, D.; Reneman, L. Test–retest reliability of task-related pharmacological MRI with a single-dose oral citalopram challenge. NeuroImage 2013, 75, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.Z.; He, J.; Liang, Y.Z.; Yuan, D.L.; Chau, F.T. Plasma fatty acid metabolic profiling and biomarkers of type 2 diabetes mellitus based on GC/MS and PLS-LDA. FEBS Lett. 2006, 580, 6837–6845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corregiari, F.M.; Gattaz, W.F.; Bernik, M. Acute hormonal changes after IV citalopram and treatment response in OCD. Psychopharmacology 2007, 193, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Hyypp, M.T.; Kangasniemi, P. Variation of plasma free tryptophan and CSF 5-HIAA during migraine. Headache 1977, 17, 25–27. [Google Scholar] [CrossRef] [PubMed]
- Wurtman, R.J.; Rose, C.M.; Chou, C.; Larin, F.F. Daily rhythms in the concentrations of various amino acids in human plasma. N. Engl. J. Med. 1968, 279, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Vecsei, L.; Ashina, M. The L-kynurenine signalling pathway in trigeminal pain processing: A potential therapeutic target in migraine? Cephalalgia 2011, 31, 1029–1038. [Google Scholar] [CrossRef]
- Nagy-Grocz, G.; Laborc, K.F.; Veres, G.; Bajtai, A.; Bohar, Z.; Zadori, D.; Fejes-Szabo, A.; Spekker, E.; Vecsei, L.; Pardutz, A. The Effect of Systemic Nitroglycerin Administration on the Kynurenine Pathway in the Rat. Front. Neurol. 2017, 8, 278. [Google Scholar] [CrossRef] [Green Version]
- Borsook, D.; Maleki, N.; Becerra, L.; McEwen, B. Understanding migraine through the lens of maladaptive stress responses: A model disease of allostatic load. Neuron 2012, 73, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Maleki, N.; Becerra, L.; Borsook, D. Migraine: Maladaptive brain responses to stress. Headache 2012, 52 (Suppl. 2), 102–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, H.; Ozaki, N.; Sawada, M.; Isobe, K.; Ohta, T.; Nagatsu, T. A link between stress and depression: Shifts in the balance between the kynurenine and serotonin pathways of tryptophan metabolism and the etiology and pathophysiology of depression. Stress 2008, 11, 198–209. [Google Scholar] [CrossRef]
- Ruddick, J.P.; Evans, A.K.; Nutt, D.J.; Lightman, S.L.; Rook, G.A.; Lowry, C.A. Tryptophan metabolism in the central nervous system: Medical implications. Expert Rev. Mol. Med. 2006, 8, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.J.; Welch, J. Stress- and endotoxin-induced increases in brain tryptophan and serotonin metabolism depend on sympathetic nervous system activity. J. Neurochem. 1991, 57, 1615–1622. [Google Scholar] [CrossRef]
- Fernstrom, J.D.; Fernstrom, M.H. Exercise, serum free tryptophan, and central fatigue. J. Nutr. 2006, 136, 553S–559S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoglund, E.; Overli, O.; Winberg, S. Tryptophan Metabolic Pathways and Brain Serotonergic Activity: A Comparative Review. Front. Endocrinol. 2019, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Ara, I.; Bano, S. Citalopram decreases tryptophan 2,3-dioxygenase activity and brain 5-HT turnover in swim stressed rats. Pharmacol. Rep. 2012, 64, 558–566. [Google Scholar] [CrossRef]
- Bano, S.; Gitay, M.; Ara, I.; Badawy, A. Acute effects of serotonergic antidepressants on tryptophan metabolism and corticosterone levels in rats. Pak. J. Pharm. Sci. 2010, 23, 266–272. [Google Scholar]
- Herhaus, B.; Joisten, N.; Wessels, I.; Zimmer, P.; Petrowski, K. How acute physical and psychological stress differentially influence the kynurenine pathway: A randomized cross-over trial. Psychoneuroendocrinology 2021, 134, 105433. [Google Scholar] [CrossRef]
- O’Farrell, K.; Harkin, A. Stress-related regulation of the kynurenine pathway: Relevance to neuropsychiatric and degenerative disorders. Neuropharmacology 2017, 112, 307–323. [Google Scholar] [CrossRef] [Green Version]
- Nagy-Grocz, G.; Tar, L.; Bohar, Z.; Fejes-Szabo, A.; Laborc, K.F.; Spekker, E.; Vecsei, L.; Pardutz, A. The modulatory effect of anandamide on nitroglycerin-induced sensitization in the trigeminal system of the rat. Cephalalgia 2016, 36, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Vamos, E.; Fejes, A.; Koch, J.; Tajti, J.; Fulop, F.; Toldi, J.; Pardutz, A.; Vecsei, L. Kynurenate derivative attenuates the nitroglycerin-induced CamKIIalpha and CGRP expression changes. Headache 2010, 50, 834–843. [Google Scholar] [CrossRef]
- Levine, E.S.; Jacobs, B.L. Neurochemical afferents controlling the activity of serotonergic neurons in the dorsal raphe nucleus: Microiontophoretic studies in the awake cat. J. Neurosci. 1992, 12, 4037–4044. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Behbehani, M.M. Physiological characteristics of the projection pathway from the medial preoptic to the nucleus raphe magnus of the rat and its modulation by the periaqueductal gray. Pain 2001, 94, 139–147. [Google Scholar] [CrossRef]
- Conti, P.; D’Ovidio, C.; Conti, C.; Gallenga, C.E.; Lauritano, D.; Caraffa, A.; Kritas, S.K.; Ronconi, G. Progression in migraine: Role of mast cells and pro-inflammatory and anti-inflammatory cytokines. Eur. J. Pharmacol. 2019, 844, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Fidan, I.; Yuksel, S.; Ymir, T.; Irkec, C.; Aksakal, F.N. The importance of cytokines, chemokines and nitric oxide in pathophysiology of migraine. J. Neuroimmunol. 2006, 171, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Kogelman, L.J.A.; Falkenberg, K.; Buil, A.; Erola, P.; Courraud, J.; Laursen, S.S.; Michoel, T.; Olesen, J.; Hansen, T.F. Changes in the gene expression profile during spontaneous migraine attacks. Sci. Rep. 2021, 11, 8294. [Google Scholar] [CrossRef]
- Sarchielli, P.; Alberti, A.; Vaianella, L.; Pierguidi, L.; Floridi, A.; Mazzotta, G.; Floridi, A.; Gallai, V. Chemokine levels in the jugular venous blood of migraine without aura patients during attacks. Headache 2004, 44, 961–968. [Google Scholar] [CrossRef]
- Domingues, R.B.; Duarte, H.; Senne, C.; Bruniera, G.; Brunale, F.; Rocha, N.P.; Teixeira, A.L. Serum levels of adiponectin, CCL3/MIP-1alpha, and CCL5/RANTES discriminate migraine from tension-type headache patients. Arq. Neuropsiquiatr. 2016, 74, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, P.P. The discovery of a new drug class for the acute treatment of migraine. Headache 2007, 47 (Suppl. 1), S10–S19. [Google Scholar] [CrossRef]
- Mikolajczyk, T.P.; Nosalski, R.; Szczepaniak, P.; Budzyn, K.; Osmenda, G.; Skiba, D.; Sagan, A.; Wu, J.; Vinh, A.; Marvar, P.J.; et al. Role of chemokine RANTES in the regulation of perivascular inflammation, T-cell accumulation, and vascular dysfunction in hypertension. FASEB J. 2016, 30, 1987–1999. [Google Scholar] [CrossRef] [Green Version]
- Vernieri, F.; Moro, L.; Altamura, C.; Palazzo, P.; Antonelli Incalzi, R.; Rossini, P.M.; Pedone, C. Patients with migraine with aura have increased flow mediated dilation. BMC Neurol. 2010, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Larsen, J.S.; Skaug, E.A.; Wisloff, U.; Ellingsen, O.; Stovner, L.J.; Linde, M.; Hagen, K. Migraine and endothelial function: The HUNT3 Study. Cephalalgia 2016, 36, 1341–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hautakangas, H.; Winsvold, B.S.; Ruotsalainen, S.E.; Bjornsdottir, G.; Harder, A.V.E.; Kogelman, L.J.A.; Thomas, L.F.; Noordam, R.; Benner, C.; Gormley, P.; et al. Genome-wide analysis of 102,084 migraine cases identifies 123 risk loci and subtype-specific risk alleles. Nat. Genet. 2022, 54, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Bruder-Nascimento, A.; Belin de Chantemele, E.J.; Bruder-Nascimento, T. CCR5 antagonist treatment inhibits vascular injury by regulating NADPH oxidase 1. Biochem. Pharmacol. 2022, 195, 114859. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Before Citalopram Neuroendocrine Challenge | p-Value | Before Placebo Infusion | p-Value | |||
|---|---|---|---|---|---|---|
| Migraine | Control | Migraine | Control | |||
| Tryptophan, kynurenine(µg/mL), cortisol (ng/mL) and cortisol/DHEA-S (µg/mL) | ||||||
| TRP | 8.92 (6.75–10.18) | 7.12 (5.55–9.28) | 0.075 | 8.20 (6.85–9.47) | 7.15 (5.59–8.11) | 0.080 |
| TRP/LNAA | 0.09 (0.06–0.11) | 0.06 (0.05–0.08) | 0.012 | 0.08 (0.07–0.11) | 0.06 (0.05–0.09) | 0.033 |
| KYN | 0.57 (0.40–0.67) | 0.46 (0.39–0.54) | 0.308 | 0.54 (0.46–0.62) | 0.47 (0.39–0.64) | 0.274 |
| KYN/TRP | 0.07 (0.05–0.07)) | 0.07 (0.06–0.07) | 0.264 | 0.06 (0.06 to 0.08) | 0.07 (0.06–0.08) | 0.481 |
| CORT | 58.74 (40.23–118.0) | 56.72 (38.15–80.86) | 0.512 | 64.21 (51.13–111.0) | 40.86 (31.23–66.54) | 0.382 |
| CORT/DHEA-S | 46.49 (37.62–85.32) | 42.10 (33.41–58.25) | 0.662 | 34.48 (35.86–82.30) | 28.88 (23.90–53.65) | 0.423 |
| Cytokines and chemokines (pg/mL) | ||||||
| GCSF | 4.19 (3.64–4.83) | 3.95 (3.54–4.91) | 0.894 | 4.18 (3.94–4.94) | 4.18 (3.39–5.13) | 0.423 |
| RANTES | 10641 (6336–11,588) | 13456 (10,192–16,846) | 0.017 | 10593 (7279–12,588) | 12670 (10,091–19,710) | 0.049 |
| MCP-1 | 15.01 (10.70–29.94) | 13.96 (8.70–19.99) | 0.538 | 16.94 (9.24–32.08) | 15.74 (8.32–22.04) | 0.388 |
| IL1a | 2.69 (2.20–3.62) | 2.42 (1.92–3.00) | 0.224 | 2.96 (2.22–3.11) | 2.85 (1.36–3.47) | 0.696 |
| IL1b | 0 (0.00–0.00) | 0 (0.00–0.00) | 1.000 | 0 (0.00–0.00) | 0 (0.00–0.00) | 1.000 |
| IL6 | 0 (0.00–0.00) | 0 (0.00–0.00) | 0.163 | 0 (0.00–0.00) | 0 (0.00–0.00) | 0.143 |
| IL8 | 0 (0.00–7.85) | 0 (0.00–6.81) | 0.748 | 7.13 (0.00–8.67) | 0 (0.00–0.00) | 0.014 |
| IL10 | 3.20 (3.14–3.39) | 3.26 (3.14–3.46) | 0.473 | 3.46 (3.32–3.61) | 3.27 (3.11–3.60) | 0.193 |
| TNFa | 1.96 (1.50–2.20) | 1.41 (0.89–2.40) | 0.233 | 1.96 (0.79–2.22) | 1.26 (0.66–2.22) | 0.752 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gecse, K.; Édes, A.E.; Nagy, T.; Demeter, A.K.; Virág, D.; Király, M.; Dalmadi Kiss, B.; Ludányi, K.; Környei, Z.; Denes, A.; et al. Citalopram Neuroendocrine Challenge Shows Altered Tryptophan and Kynurenine Metabolism in Migraine. Cells 2022, 11, 2258. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11142258
Gecse K, Édes AE, Nagy T, Demeter AK, Virág D, Király M, Dalmadi Kiss B, Ludányi K, Környei Z, Denes A, et al. Citalopram Neuroendocrine Challenge Shows Altered Tryptophan and Kynurenine Metabolism in Migraine. Cells. 2022; 11(14):2258. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11142258
Chicago/Turabian StyleGecse, Kinga, Andrea Edit Édes, Tamás Nagy, Adrienn Katalin Demeter, Dávid Virág, Márton Király, Borbála Dalmadi Kiss, Krisztina Ludányi, Zsuzsanna Környei, Adam Denes, and et al. 2022. "Citalopram Neuroendocrine Challenge Shows Altered Tryptophan and Kynurenine Metabolism in Migraine" Cells 11, no. 14: 2258. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11142258