
Regulation of Conidiogenesis in Aspergillus flavus
by
 and
and
He-Jin Cho
1,
Sung-Hun Son
1,
Wanping Chen
2,
Ye-Eun Son
1,
Inhyung Lee
3,
Jae-Hyuk Yu
4,5 and
Hee-Soo Park
1,6,* 1
School of Food Science and Biotechnology, Kyungpook National University, Daegu 41566, Korea
2
Department of Molecular Microbiology and Genetics, University of Göttingen, 37077 Göttingen, Germany
3
Department of Bio and Fermentation Convergence Technology, Kookmin University, Seoul 02707, Korea
4
Department of Bacteriology, University of Wisconsin, Madison, WI 53706, USA
5
Department of Systems Biotechnology, Konkuk University, Seoul 05029, Korea
6
Department of Integrative Biology, Kyungpook National University, Daegu 41566, Korea
*
Author to whom correspondence should be addressed.
Cells 2022, 11(18), 2796; https://0-doi-org.brum.beds.ac.uk/10.3390/cells11182796
Submission received: 5 August 2022
/
Revised: 29 August 2022
/
Accepted: 6 September 2022
/
Published: 7 September 2022
(This article belongs to the Special Issue State-of-Art in Aspergillus)
Abstract
:Aspergillus flavus is a representative fungal species in the Aspergillus section Flavi and has been used as a model system to gain insights into fungal development and toxin production. A. flavus has several adverse effects on humans, including the production of the most carcinogenic mycotoxin aflatoxins and causing aspergillosis in immune-compromised patients. In addition, A. flavus infection of crops results in economic losses due to yield loss and aflatoxin contamination. A. flavus is a saprophytic fungus that disperses in the ecosystem mainly by producing asexual spores (conidia), which also provide long-term survival in the harsh environmental conditions. Conidia are composed of the rodlet layer, cell wall, and melanin and are produced from an asexual specialized structure called the conidiophore. The production of conidiophores is tightly regulated by various regulators, including the central regulatory cascade composed of BrlA-AbaA-WetA, the fungi-specific velvet regulators, upstream regulators, and developmental repressors. In this review, we summarize the findings of a series of recent studies related to asexual development in A. flavus and provide insights for a better understanding of other fungal species in the section Flavi.
1. Introduction
The genus Aspergillus comprises more than 300 species, divided into 19 sections [1]. Among them, fungal species belonging to Aspergillus section Flavi are important for agriculture, biotechnology, the food industry, and human health [2]. For example, Aspergillus flavus is a major virulent fungus of maize, peanuts, and corn during crop harvesting, often coupled with the production of the most potent carcinogen in the nature aflatoxins, leading to great damage to crops, and economic loss [3,4]. In addition, A. flavus is the second most common agent of invasive aspergillosis [5,6]. Along with A. flavus, other species belonging to the section Flavi also produce various mycotoxins that have detrimental effects on humans [2]. Conversely, several species have beneficial applications in fermented foods, biotechnology, and pharmaceuticals [7]. For example, A. oryzae is used in traditional fermented foods, such as meju, soy sauce, miso, and sake [8,9]. A. tamarii and A. alliaceus are used for the enzymes they produce, such as amylases, protease, and pectin-degrading enzymes [10,11].
Most species in the genus Aspergillus reproduce through the production of spores released into the environment [12]. Some Aspergillus species produce sexual spores, but most Aspergillus species reproduce via asexual spores called conidia [13,14]. The asexual spore is carried on the conidiophore, an asexual specialized structure [15,16]. Unlike hyphae, conidia have a thick cell wall structure, which affords resistance to environmental conditions as well as protection from the immune system of the host organism [14,15]. In addition, conidia contain a variety of secondary metabolites, such as melanin and mycotoxins, which play key roles in pathogenesis and development [17,18]. The important processes of asexual spore formation, maturation, dormancy, and germination are controlled by several transcription factors and regulators [19,20]. Studies of asexual development in Aspergillus have focused on the model species A. nidulans [21]. Central regulators (BrlA, AbaA, and WetA), velvet regulators, upstream regulators (FlbA-E and FluG), and other components of the signaling pathway are involved in the regulation of asexual development [20]. The results obtained for A. nidulans have been applied to other Aspergilli; however, some of these regulators play diverse roles in members of Aspergilli. Recently, owing to advances in molecular and genetic technology, research on other species of Aspergillus has gradually expanded. In particular, whole-genome genetic information for representative strains in this section was presented in 2017 [22], and genomic information for 23 species of the Aspergillus section Flavi was recently studied, laying the foundation for customized research on various fungal species [23]. Based on the whole genomic information of strains belonging to Aspergillus section Flavi, genes encoding carbohydrate-active enzymes (CAZymes) and predicted secondary metabolite backbone genes have recently been analyzed [23]. These discoveries are expected to form the basis for the industrial production of enzymes or secondary metabolites.
A.flavus, a representative fungal species in Flavi, is more useful for understanding developmental processes in the section Flavi than is A. nidulans [3]. The species belonging to the Aspergillus section Flavi and A. nidulans have a common feature of generating an asexual specialized structure called conidiophore [12]. However, there are differences in the detailed morphology of asexual and sexual developmental structures, and metabolites. The main difference is that most species belonging to the Aspergillus section Flavi produce dark-colored sclerotia containing ascospore-bearing ascocarps [2], but A. nidulans produces cleistothecia surrounded by numerous Hülle cells [24,25]. In case of the morphology of conidiophores, A. flavus produces uni- or biseriate conidiophores, while A. nidulans produces biseriate conidiophores. For secondary metabolites, most species belonging to the Aspergillus section Flavi produce aflatoxins, but A. nidulans produces sterigmatocystin, which is a precursor of aflatoxins but not aflatoxins. As a result of several recent studies, new insights into the development and metabolism of A. flavus have been published [26,27]. Accordingly, this review aims to explore the distribution of developmental regulators in the strains belonging to Flavi and summarize the roles of developmental regulators in A. flavus. Developmental regulators related to asexual development are described in detail.
2. Distribution of Key Developmental Regulators in Aspergillus
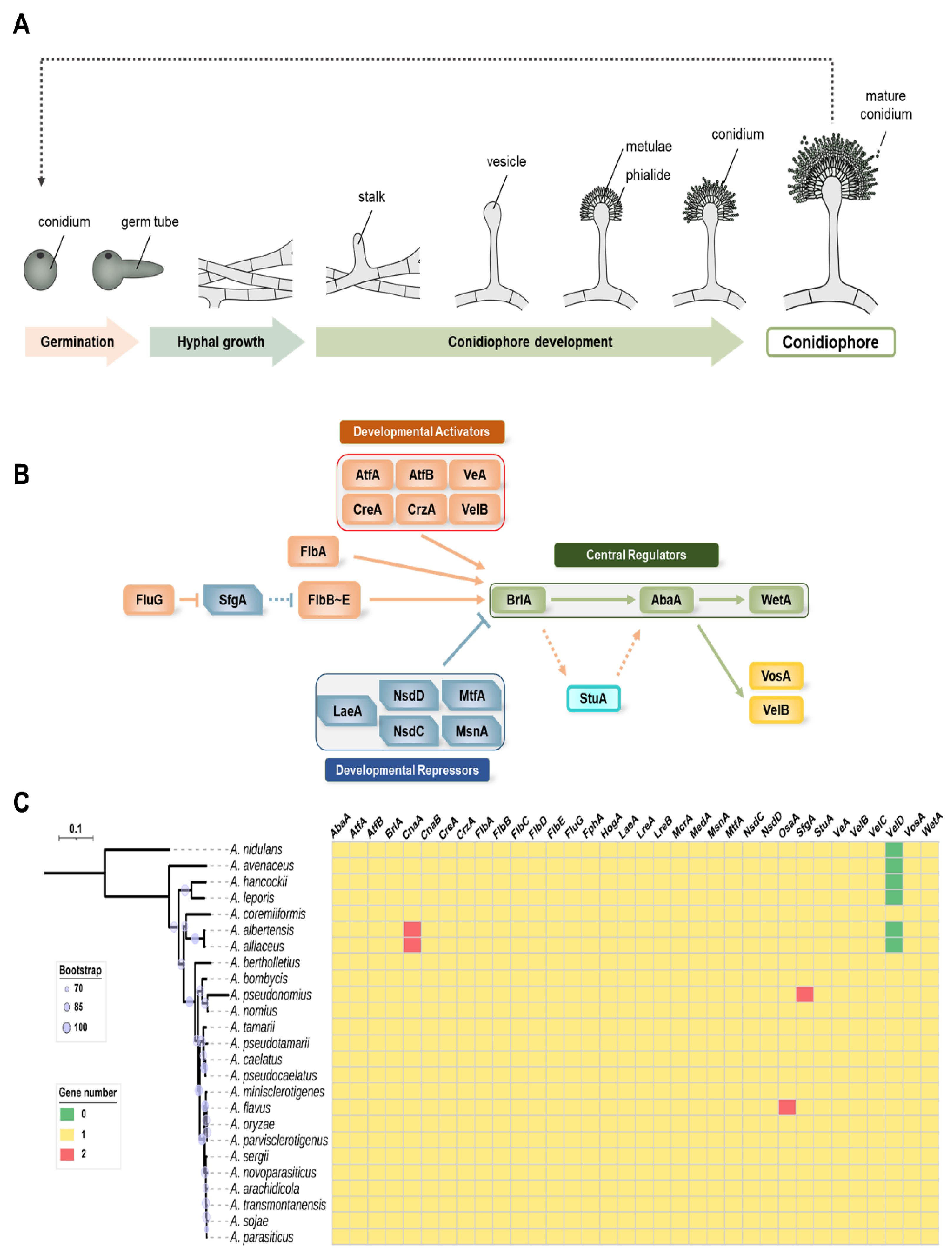
Several research groups have studied asexual development in Aspergillus spp. and have reported a variety of genes and proteins involved in the cycle of conidia formation (Figure 1A) [19,20,28]. We focused on 34 genes that are important for conidial formation, maturation, and dormancy (Figure 1B). The distribution of the 34 genes in 16 species of Aspergillus section Flavi is shown in Figure 1C. Most of the tested species of Aspergillus section Flavi contained one homolog of these genes. The genomes of A. albertensis and A. alliaceus contain two cnaA genes, encoding the catalytic subunit of calcineurin, whereas A. pseudonomius has two SfgA (suppressor of fluG A) homologs and A. flavus has two potential OsaA (orchestrator of sexual and asexual development A) homologs. Interestingly, VelD, the Velvet protein, has no homolog in A. nidulans, whereas most species belonging to Aspergillus section Flavi have it. However, VelD has not been found in several species, including A. avenaceus, A. hanockii, A. leporis, A. albertensis, and A. alliaceus. The roles of the studied genes are summarized in Table 1 and discussed below. Among 34 regulators, the roles of 26 genes were characterized in A. flavus. Other genes including cnaA, cnaB, fphA, lreA, lreB, mcrA, medA, and osaA have not been characterized, and further studies will be needed to understand A. flavus development.
2.1. Central Regulators of Conidiophore Production in Aspergillus
During asexual development, Aspergillus spp. generate conidiophores, which are composed of vesicles, one or two layers of sterigmata, and conidia [45]. A. flavus is biserate (metulae and phialide) or uniserate (phialide) species. The conidia of these species are produced by conidiogenous cell phialides [46]. Conidiophore formation in Aspergillus spp. is regulated by various regulators [20]. The expression of genes involved in conidiophore production is spatiotemporally regulated by three transcription factors: BrlA, AbaA, and WetA [13,47]. These transcription factors are highly conserved in most Aspergillus species [19]. Studies on these transcription factors have been mainly conducted in A. nidulans and A. fumigatus, but not much research has been done on other species of Aspergillus [47,48,49,50]. In A. flavus, the role of WetA, but not BrlA and AbaA, has been characterized [44,51,52]. Therefore, the roles of BrlA and AbaA were described based on our current study and studies in other species.
2.1.1. BrlA
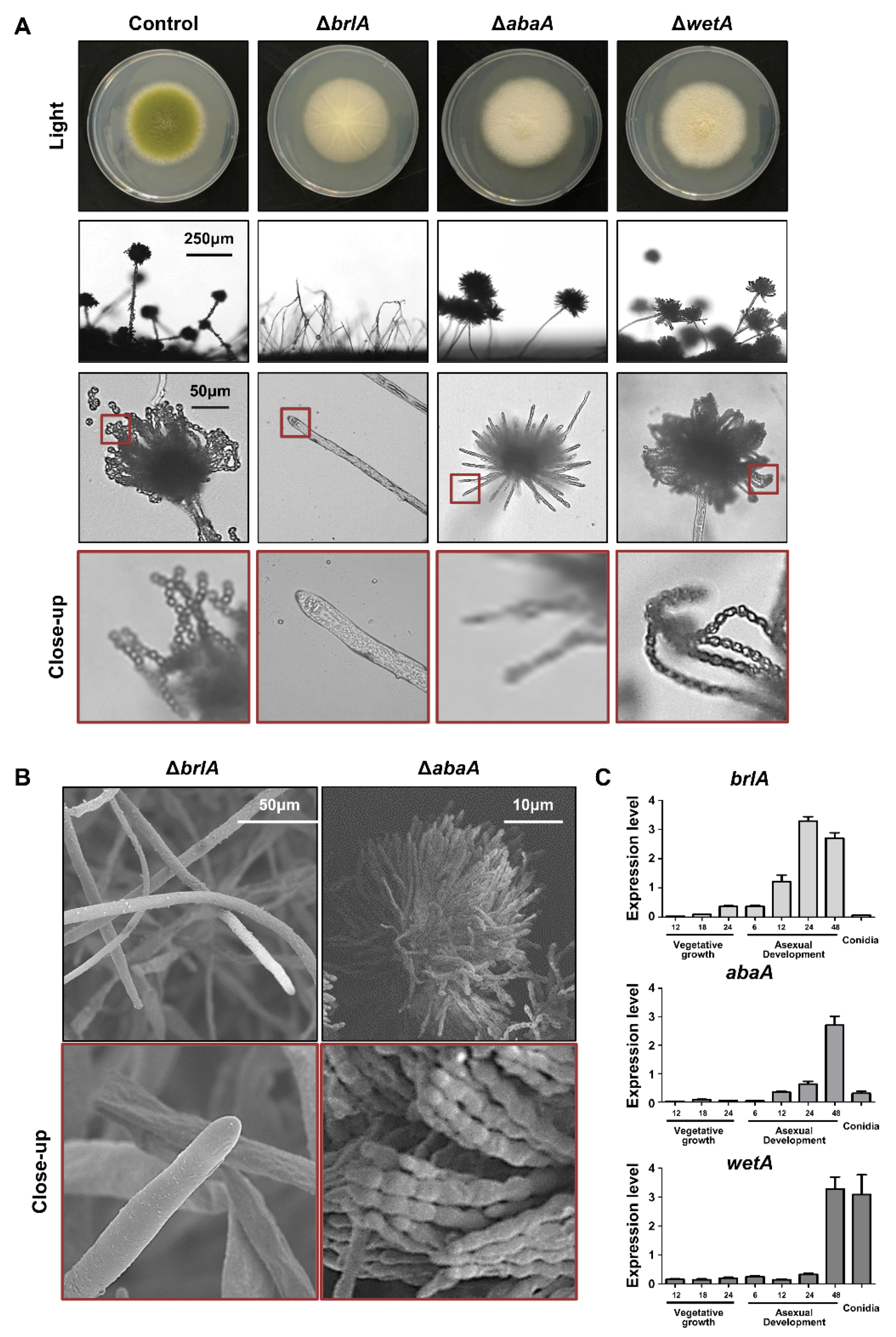
The brlA (bristle A) gene was first identified through mutational analysis of asexual development in A. nidulans [53]. The brlA mutants are aconidial, and these mutant strains exhibit a bristle-like phenotype [47]. This aconidial phenotype was similar to that of A. flavus. As shown in Figure 2A, the brlA mutant forms white colonies and cannot produce a proper conidiophore. These results are similar to those of the scanning electron microscopes image; where it can be seen that the brlA deletion mutants formed a hyphal tip but did not form a conidiophore (Figure 2B). In A. oryzae, the brlA mutant exhibits no conidiation and increased hyphal growth [54,55], suggesting that the function of brlA in conidiophore production is conserved in Aspergillus species.
BrlA is a key transcription factor for the initiation of conidiophore formation, which contains a C2H2 zinc finger DNA-binding domain [56,57,58]. In A. nidulans, BrlA is highly expressed during the initiation of conidiation and regulates genes containing the BrlA response element (BRE, 5′-CAAGGGG-3′) at the promoter region in the target genes (e.g., abaA) [57]. Similar to A. nidulans, brlA mRNA was highly expressed during the early phase of conidiation and was not detected in the conidia of A. flavus (Figure 2C). BrlA has been reported to affect sexual development and secondary metabolism [45,59]. For example, the brlA mutant cannot produce cleistothecia in A. nidulans [45]. In A. flavus, sclerotia production was shown to be decreased in the brlA mutant (Figure 3A), implying that BrlA plays a diverse role in sexual reproduction in Aspergillus spp. The production of aflatoxin was increased in the brlA mutant strains (Figure 3B). These results suggest that BrlA is conserved in Aspergillus conidiation and is involved in fungal reproduction and secondary metabolism.
2.1.2. AbaA
AbaA is a key regulator of gene expression during the middle phase of conidiation [60,61]. The typical phenotype of the abaA mutant is an abacus structure that incompletely differentiates phialides from the conidial chain in A. nidulans [60]. This phenotype has also been observed in other Aspergillus species, such as A. fumigatus and A. oryzae [49,54]. In A. flavus, the abaA mutant also exhibited white colonies and abacus-like phialides (Figure 2), similar to what is found in other species. AbaA also affects the proper production of sclerotia and aflatoxin B (Figure 3).
AbaA is a transcription factor containing a TEA/ATTS domain [43]. The abaA gene was highly expressed during the middle and late phases of conidiation but not in conidia (Figure 2C). It has been reported that AbaA recognizes the DNA sequence 5′-CATTCY-3′ (Y is a pyrimidine), called AbaA response element (ARE) [62]. AREs are present in the promoter regions of AbaA target genes such as wetA, vosA, and rodA [63]. In A. flavus, these target genes also contain AREs in their promoter regions, and mRNA expression of these target genes is regulated by AbaA. These molecular, bioinformatics, and morphological results strongly support the idea that the role of AbaA is conserved for regulating the middle phase of conidiation in Aspergillus species.
2.1.3. WetA
WetA is a DNA-binding transcription factor with an ESC1/WetA-related domain that directly or indirectly regulates genes involved in conidial formation, wall integrity, and metabolism [64]. The wetA deletion mutant exhibits a white colony, but the shape of the conidiophore is similar to that of the wild-type (Figure 2). However, when observed under a transmission electron microscope, there is a significant difference in spore size and wall integrity [44]. In addition, the amount of β-glucan increased, but the content of trehalose decreased in the mutant spores [44]. The RNA-seq results demonstrate that WetA coordinates the mRNA expression of genes related to trehalose, chitin, glucan, and melanin metabolic pathways [44]. The role of WetA is highly conserved among Aspergillus species. Wu and colleagues found a potential WetA response element (WRE) via the ChIP-seq in A. nidulans, and WREs can be found in the promoter regions of potential WetA target genes in A. fumigatus and A. flavus [52]. These results suggest that WetA is a key regulator of the gene regulatory network in Aspergillus conidia. Similar to other central regulators, the loss of wetA decreased sclerotia production but increased aflatoxin B1 production (Figure 3).
2.2. Upstream Regulators in Asexual Development
For the formation of conidiophores to start in hyphae, developmental competence, the ability to respond to developmental cues, must be acquired [65,66]. Until developmental competence is acquired, upstream developmental activators occupy the promoter region of brlA, but conidiation does not begin because several repressors bind the promoter of brlA and block the role of upstream developmental activators [67]. After developmental competence is acquired, repressors are removed from the brlA promoter and brlA expression is induced to initiate conidiation [20]. Therefore, to understand the initiation of asexual development, it is important to understand the upstream activators or repressors and the signaling pathways involved in the initiation of brlA.
2.2.1. Developmental Activators
The fluffy genes are well-studied genes that encode developmental activators involved in the expression of brlA in A. nidulans [16]. Mutational analysis identified six fluffy genes: fluG, flbA, flbB, flbC, flbD, and flbE [20,68]. The loss of function mutations in any one of these genes results in a fluffy colony.
FluG contains two domains: an amidohydrolase domain in the N-terminal region and a γ-glutamyl ligase region in the C-terminal region [69,70,71]. FluG is involved in the production of dehydroaustinol, a diffusible activator of conidiation in A. nidulans [72,73]. Therefore, FluG is a key gene in the initiation of conidiation. Unlike A. nidulans, ΔfluG mutants can produce asexual spores of A. flavus [35]. The deletion of fluG causes reduced conidial production and brlA expression but increased sclerotia production, suggesting that FluG may act as a balancer for asexual and sexual development [35]. Further studies have found that FluG can interact with VelB or LaeA, which then controls conidia and sclerotia production [74].
FlbA is one of the regulators of G protein signaling (RGS) proteins that are involved in the G protein signaling pathway [75,76]. FlbA affects fungal development and aflatoxin production in A. flavus [34]. The deletion of flbA decreased conidia production and brlA expression. In addition, the ΔflbA mutant strains show decreased pathogenicity and toxigenicity [34]. In A. nidulans, FlbA is involved in the FadA-cAMP/PKA pathway; however, in A. flavus, FlbA does not affect the FadA signaling pathway or the regulation of intracellular cAMP concentration [34].
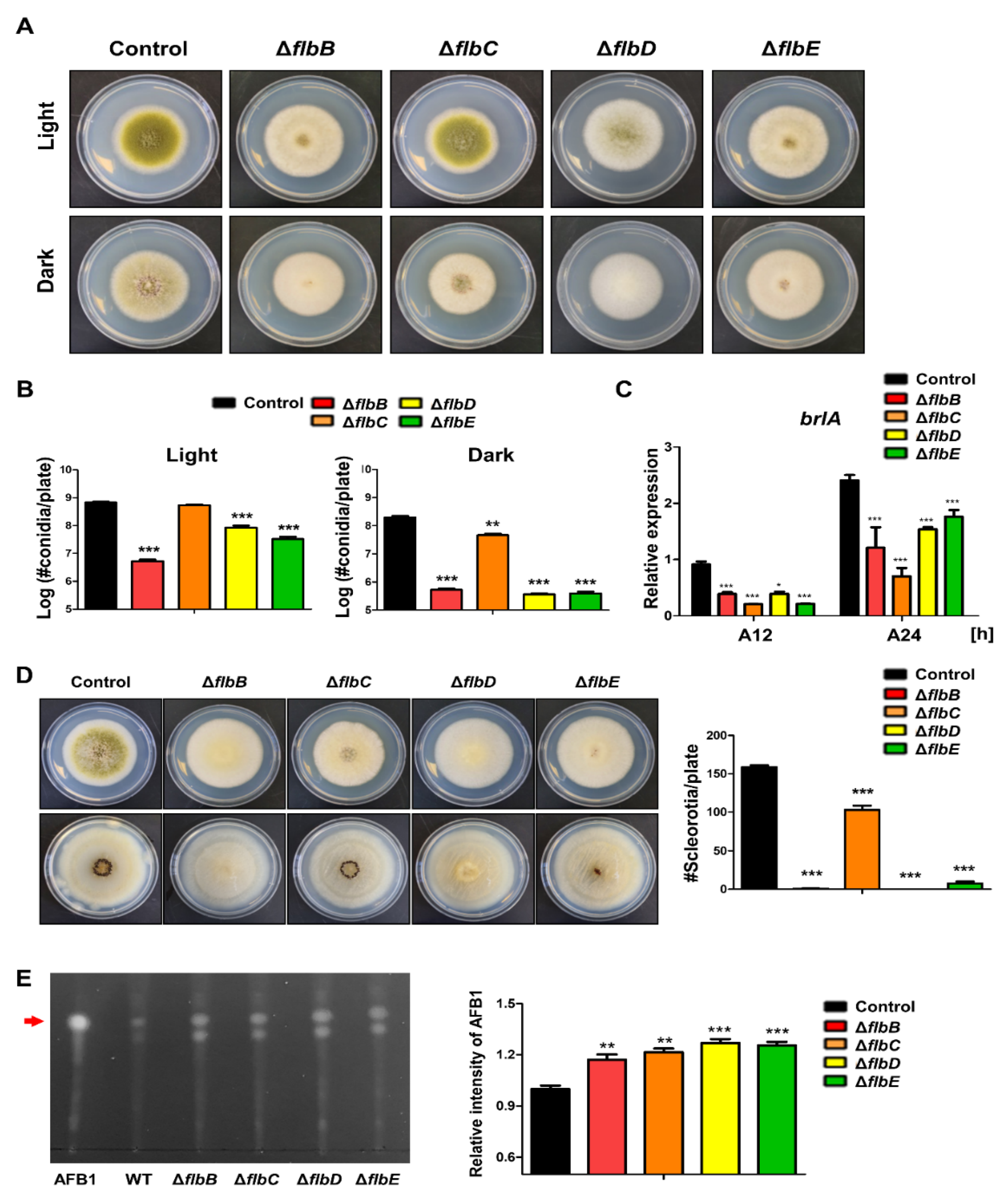
flbB–flbD are fluffy genes that encode DNA-binding transcription factors [67]. The roles of FlbB–FlbE have been studied in A. nidulans, A. fumigatus, and A. oryzae, and these transcription factors are key components for conidiophore initiation and brlA expression [54,67,77,78,79,80,81,82,83]. In A. flavus, the roles of FlbB–FlbE are similar to those in other Aspergilli, but they are slightly different from those in A. nidulans. In A. nidulans, the loss of flbB~flbE has been shown to cause delayed or decreased production of conidiophores and brlA expression. In A. flavus, each mutant strain of flbB, flbD, and flbE can produce a small number of asexual spores, the flbC mutant produces asexual spores similar to the wild-type strains in light conditions (Figure 4). In terms of brlA expression, all flbB~flbE strains express brlA to a lesser degree than the wild-type (Figure 4C), which suggests that flbB~flbE are important for the initiation of conidiophore production.
The functions of FlbB–FlbE in asexual spore formation are conserved in both A. nidulans and A. flavus (Table 2). However, sexual development has slightly different functions. In A. nidulans, the production of sexual fruiting bodies, called cleistothecia, was increased in the flbC mutant, but the amount of sclerotia was decreased in A. flavus. Overall, although their roles in sexual development and secondary metabolites are somewhat different, they are similar in asexual reproduction in Aspergillus species.
2.2.2. Velvet Regulators and LaeA
Velvet proteins contain a fungus-specific DNA-binding domain, called the velvet domain, which is a transcription factor that regulates multiple genes [84,85]. In most Aspergillus species, the velvet protein family consists of four proteins, including VeA, VelB, VelC, and VosA, but there are five proteins in A. flavus and other Flavi species (Figure 1B) [43]. Velvet proteins also form homo- or heterocomplexes that coordinate fungal reproduction and secondary metabolism [86,87]. VeA has been studied in several fungal species and plays an important role in fungal development and secondary metabolism [86,88]. In A. flavus, the deletion of veA results in decreased conidia formation, the absence of sclerotia, and aflatoxin B formation [37,74]. VeA regulates lipid degradation in seeds, which affects plant pathogenesis [37]. VeA also forms a complex with VelB and LaeA, which controls sexual development and aflatoxin production [37,74]. It has been reported that VelB, a partner protein of VeA, functions similarly to VeA in sexual reproduction, conidiophore formation, and aflatoxin production. However, VelB also interacts with other proteins, such as FluG or VosA, and these complexes are involved in other processes. It has been predicted that VelB functions together with FluG during asexual spore formation [74]. The VelB-VosA complex governs conidia maturation and stress response in conidia [43]. VelC and VelD do not appear to play a significant role in asexual spore formation, but VelD affects the aflatoxin formation process [43]. VosA is a key regulator for the viability of spores in A. nidulans. As mentioned above, VosA plays a key role in spore maturation and dormancy after conidia formation by binding to VelB. The deletion of vosA or velB causes increased stress sensitivity and decreases the amount of trehalose in A. flavus conidia [43]. VelB and VosA are representative transcription factors that regulate the mRNA expression of spore-specific genes, together with WetA, after spore formation [51]. These regulators control the genes associated with chitin and beta-glucan biosynthesis, trehalose synthesis, and secondary metabolism in A. nidulans and A. flavus conidia [51,52].
LaeA is a putative methyltransferase that has been identified as a forward genetic screen in A. nidulans [89]. It has been reported that the main function of LaeA in various fungal species is the production of sterigmatocystin and other secondary metabolites [90]. Importantly, as mentioned above, LaeA interacts with other proteins to form complexes, such as VeA-VelB-LaeA or VeA-VelB-LaeA-FluG, which coordinate aflatoxin production and fungal development [54,66]. We predicted that members of these complexes would perform similar functions in conidia production in A. flavus, but the phenotypes of each mutant were slightly different. For example, the deletion veA or velB led to decreased conidia production, but the laeA mutant exhibited increased conidia production, and the morphology of laeA mutant conidiophores was abnormal [91]. These results support the idea that LaeA and VeA/VelB play different roles in the asexual development of A. flavus compared with A. nidulans [74].
2.3. Other Key Regulators for Asexual Development
2.3.1. AtfA and AtfB
AtfA and AtfB are basic leucine zipper (bZIP) transcription factors that affect fungal development, metabolism, and stress responses [92,93,94]. Importantly, AtfA is one of the components of the high-osmolarity glycerol (HOG) MAPK cascade, which is a key signaling pathway for hyperosmotic and oxidative stress response [31]. In A. flavus, the absence of atfA or atfB leads to decreased fungal growth, conidiation, sclerotia production, and aflatoxin B1 production. In addition, AtfA and AtfB, but not osmotic and alkali stress, are required for the oxidative stress response [31]. These results provide insights into the roles of AtfA and AtfB in conidiophore production and stress response in A. flavus.
2.3.2. CreA
Carbon is the main energy source, and carbon catabolite repression (CCR) is required for the regulation of development and metabolic processes in fungi [95]. CreA contains the Cys2-His2 zinc finger domain and acts as a major transcriptional repressor in A. nidulans [96,97]. Fasoyin et al. characterized the function of CreA and found that the deletion of creA caused the production of abnormal conidiophores, decreased brlA and abaA mRNA expression, and the production of aflatoxin in A. flavus [32]. The overexpression of creA increases the number of conidia and aflatoxin B1 production in maize seeds. These results suggest that CreA acts as a positive regulator of asexual development in A. flavus.
2.3.3. CrzA
The calcineurin-Crz1 signaling pathway plays a key role in the stress response, development, and virulence of pathogenic fungi [98]. Crz1/CrzA is a C2H2-type transcription factor that is a key target of calcineurin in yeast and filamentous fungi [99,100,101,102]. In A. flavus, the loss of crzA resulted in reduced conidial production and abnormal conidiophore production [33]. The mRNA levels of brlA, abaA, and wetA significantly decreased in the crzA deletion mutant strains during asexual development. Fungal growth and sclerotia production were decreased in crzA deletion mutants [82]. Although the crzA mutant strain was generated, it was difficult to generate cnaA or cnaB deletion mutants. In addition, it was difficult to generate calcineurin mutants in other Aspergillus, or calcineurin mutant strains grow very slowly [103], implying that the calcineurin-Crz1 signaling pathway is crucial for fungal growth and the initiation of asexual and sexual development.
2.3.4. HogA (SakA)
The high-osmolarity glycerol (HOG) pathway is necessary for tolerance to a variety of environmental stresses and immune responses in the host system [104,105]. HogA (also called SakA) is a key kinase in the HOG pathway in fungi [106]. In A. flavus, HogA is a key factor for osmotic stress response, aflatoxin B1 production, and virulence [36]. HogA also affects conidiation. The deletion of hogA led to decreased conidiophore production under normal conditions but increased the number of conidia under osmotic stress conditions or in seeds, suggesting that HogA plays a diverse role in asexual development depending on the environmental conditions.
2.3.5. MsnA
MsnA is an ortholog of S. cerevisiae Msn2, which contains a C2H2-type zinc-finger domain [107]. MsnA is associated with the HOG pathway, which is crucial for stress response in A. nidulans [89]. In A. flavus and A. parasiticus, MsnA plays a similar role in the stress response [38]. In addition, the deletion of msnA decreased fungal growth but increased conidia production [38]. MsnA also affects secondary metabolism, and the msnA deletion mutant produces more aflatoxin B1 and kojic acid. MsnA plays a key role in both stress response and fungal development.
2.3.6. MtfA
MtfA is a master transcription factor for secondary metabolite production in Aspergillus species [108,109]. This protein contains a C2H2 DNA-binding domain that governs the production of secondary metabolites. In A. flavus, mtfA overexpression mutants cannot produce aflatoxin and can affect the production of several secondary metabolites [39]. For asexual development, the deletion of mtfA causes increased conidia production and induces brlA expression, but the overexpression of mtfA causes a decrease in the number of conidia and brlA expression. These results strongly support the idea that MtfA negatively regulates A. flavus asexual development.
2.3.7. NsdC and NsdD
NsdC (never in sexual development) is a transcription factor containing a C2H2 zinc finger-type DNA-binding domain [110]. NsdC is a gene identified through genetic screening, together with NsdD, which contains a GATA-type zinc finger domain. nsdC or nsdD deletion mutants cannot produce sexual structures in A. nidulans [111]. NsdD acts as a repressor of brlA expression, which is exerted by directly binding to the promoter of brlA in A. nidulans [112]. Both genes affect asexual development and aflatoxin production in A. flavus [40]. The deletion of nsdC or nsdD causes the production of abnormal conidiophores, with shortens stipes and alters conidial heads. In addition, the ΔnsdC and ΔnsdD mutants exhibited abnormal increases in the expression of brlA and abaA. In the case of NsdC, the ΔnsdC strain produced more conidiophores than the control strain. RNA-seq analysis demonstrated that NsdC affects the mRNA expression of asexual developmental genes such as brlA, abaA, wetA, vosA, and yA [113]. Lee and colleagues found that nsdD deletion mutants produced abnormal conidiophores in liquid submerged cultures and formed abundant conidiophores embedded in agar [112]. Overall, these results strongly suggest that NsdC and NsdD are key negative regulators of conidiation and may directly regulate brlA expression in A. flavus.
2.3.8. SfgA
SfgA (suppressors of fluG), which contains a Zn2Cys6 DNA-binding domain, is a negative regulator of asexual development functioning downstream of FluG in A. nidulans [114,115]. The role of SfgA in A. flavus has been examined by Yuan et al. [41]. Phenotypic analyses of the sfgA deletion or overexpression mutant demonstrated that SfgA plays a similar role in asexual development in Aspergillus species. The sfgA deletion mutant can produce asexual developmental structures in liquid submerged culture, implying that SfgA functions as a repressor of conidiophore production. Genetic analysis has revealed that SfgA is a downstream transcription factor of FluG. However, it is unclear whether SfgA is an upstream regulator of FlbB-E, and further studies are needed to confirm this.
2.3.9. StuA
The stuA (stunted) gene encodes a protein that is a member of the APSES transcription factors (Asm1p, Phd1p, Sok2p, Efg1p, and StuAp) [116,117] In A. nidulans, StuA controls the mRNA expression of brlA and abaA, which then modulate asexual sporulation [117]. In A. fumigatus, stuA mRNA is expressed after the acquisition of developmental competence. In addition, the ΔstuA mutant strains produced abnormal conidiophores and a small number of conidia, suggesting that StuA is crucial for proper conidiophore production in A. fumigatus [118]. Yao et al. demonstrated that StuA plays a key role in fungal development and secondary metabolism in A. flavus [42]. The stuA mutants exhibited impaired conidiophore production. StuA affects brlA and abaA expression, which affects the expression of downstream genes, thereby resulting in conidiophore formation. Moreover, the absence of stuA results in defects in conidiation and aflatoxin production in peanut and maize seeds. Overall, these results suggested that StuA is a key regulator of conidiation and metabolism in A. flavus.
3. Conclusions
With recent advances in next-generation sequencing, the genomes of various Aspergillus species have been reported, but knowledge based on biological and molecular research is required to understand fungal biology in detail. For this purpose, A. nidulans was primarily used as a model fungus for all Aspergillus species. However, due to the development of genetic and molecular techniques in recent decades, a variety of studies have been conducted on A. flavus, which have provided useful information for understanding the fungi belonging to Aspergillus section Flavi. A. flavus has been studied mainly due to its toxin production and pathogenicity, but to understand all of this, it is necessary to understand asexual development, the major reproductive mode of Aspergillus spp. In this review, the functions of the various developmental regulators involved in asexual reproduction were described and discussed. With these developmental regulators, the function of other transcription factors was published (Table 3). However, most studies have derived functions based on phenotypic analyses. In the near future, it will be necessary to systematically understand asexual development using new techniques such as transcriptomic, metabolomic, and metagenomic analyses. It is hoped that this will provide insights to understand fungal development in A. flavus and other fungal species in the Aspergillus section Flavi.
Author Contributions
Data curation, H.-J.C., S.-H.S., W.C. and Y.-E.S.; conceptualization, H.-S.P. and J.-H.Y.; writing—original draft preparation, H.-J.C., I.L. and H.-S.P.; supervision, project administration; and funding acquisition, H.-S.P. and J.-H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a National Research Foundation of Korea (NRF) grant to HSP funded by the Korean government (NRF-2020R1C1C1004473) and a project to train professional personnel in biological materials by the Ministry of Environment. The work at UW-Madison was supported by Food Research Institute.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.; Hubka, V.; Ezekiel, C.; Hong, S.-B.; Nováková, A.; Chen, A.; Arzanlou, M.; Larsen, T.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus section Flavi and their production of aflatoxins, ochratoxins and other mycotoxins. Stud. Mycol. 2018, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Aspergillus flavus . Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Klich, M.A. Aspergillus flavus: The major producer of aflatoxin. Mol. Plant Pathol. 2007, 8, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Manavathu, E.K.; Chandrasekar, P.H. Aspergillus flavus: An emerging non-fumigatus Aspergillus species of significance. Mycoses 2009, 52, 206–222. [Google Scholar] [CrossRef]
- Rudramurthy, S.M.; Paul, R.A.; Chakrabarti, A.; Mouton, J.W.; Meis, J.F. Invasive aspergillosis by Aspergillus flavus: Epidemiology, diagnosis, antifungal resistance, and management. J. Fungi 2019, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Jun, S.-C.; Han, K.-H.; Hong, S.-B.; Yu, J.-H. Diversity, application, and synthetic biology of industrially important Aspergillus fungi. Adv. Appl. Microbiol. 2017, 100, 161–202. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.-J.; Hu, S.; Wang, B.-T.; Jin, L. Advances in genetic engineering technology and its application in the industrial fungus Aspergillus oryzae. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Abe, K.; Asai, K.; Gomi, K.; Juvvadi, P.R.; Kato, M.; Kitamoto, K.; Takeuchi, M.; Machida, M. Genomics of Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2007, 71, 646–670. [Google Scholar] [CrossRef] [PubMed]
- Batomunkueva, B.P.; Egorov, N.S. Preparations of extracellular proteinases from Aspergillus ochraceus 513 and Aspergillus al-liaceus 7dN1. Mikrobiologiia 2002, 71, 56–58. [Google Scholar]
- Varga, J.; Frisvad, J.; Samson, R. Two new aflatoxin producing species, and an overview of Aspergillus section Flavi. Stud. Mycol. 2011, 69, 57–80. [Google Scholar] [CrossRef]
- Bennett, J.W. An Overview of the Genus Aspergillus. In The Aspergillus; Molecular Biology and Genomics; CRC Press: Boca Raton, FL, USA, 2010; pp. 1–17. [Google Scholar]
- Adams, T.H.; Wieser, J.K.; Yu, J.-H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Ebbole, D.J. The Conidium. In Cellular and Molecular Biology of Filamentous Fungi; American Society for Microbiology Press: Washington, DC, USA, 2010; pp. 577–590. [Google Scholar]
- Baltussen, T.J.H.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular mechanisms of conidial germination in Aspergillus spp. Microbiol. Mol. Biol. Rev. 2020, 84, e00049-19. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Yu, J.-H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Blachowicz, A.; Raffa, N.; Bok, J.W.; Choera, T.; Knox, B.; Lim, F.Y.; Huttenlocher, A.; Wang, C.C.C.; Venkateswaran, K.; Keller, N.P. Contributions of spore secondary metabolites to UV-C protection and virulence vary in different Aspergillus fumigatus strains. mBio 2020, 11, e03415-19. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.H.; Ramaswamy, A.; Sim, S.C.; Keller, N.P. Increased conidiation associated with progression along the sterig-matocystin biosynthetic pathway. Mycologia 2004, 96, 1190–1198. [Google Scholar] [CrossRef]
- Ojeda-López, M.; Chen, W.; Eagle, C.; Gutiérrez, G.; Jia, W.; Swilaiman, S.; Huang, Z.; Park, H.-S.; Yu, J.-H.; Cánovas, D.; et al. Evolution of asexual and sexual reproduction in the aspergilli. Stud. Mycol. 2018, 91, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Lee, M.-K.; Han, K.-H.; Kim, M.-J.; Yu, J.-H. Developmental decisions in Aspergillus nidulans. Biol. Fungal Cell 2019, 63–80. [Google Scholar] [CrossRef]
- Casselton, L.A.; Zolan, M.E. The art and design of genetic screens: Filamentous fungi. Nat. Rev. Genet. 2002, 3, 683–697. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Kjærbølling, I.; Vesth, T.; Frisvad, J.C.; Nybo, J.L.; Theobald, S.; Kildgaard, S.; Petersen, T.I.; Kuo, A.; Sato, A.; Lyhne, E.K.; et al. A comparative genomics study of 23 Aspergillus species from section Flavi. Nat. Commun. 2020, 11, 1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyer, P.S.; O’Gorman, C.M. Sexual development and cryptic sexuality in fungi: Insights from Aspergillus species. FEMS Microbiol. Rev. 2012, 36, 165–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Frisvad, J.; Sun, B.; Varga, J.; Kocsubé, S.; Dijksterhuis, J.; Kim, D.; Hong, S.-B.; Houbraken, J.; Samson, R. Aspergillus section Nidulantes (formerly Emericella): Polyphasic taxonomy, chemistry and biology. Stud. Mycol. 2016, 84, 1–118. [Google Scholar] [CrossRef] [PubMed]
- Affeldt, K.J.; Carrig, J.; Amare, M.; Keller, N.P. Global survey of canonical Aspergillus flavus G protein-coupled receptors. mBio 2014, 5, e01501–14. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.K.; Mack, B.M.; Moore, G.G.; Downey, D.L.; Lebar, M.; Joardar, V.; Losada, L.; Yu, J.; Nierman, W.C.; Bhatnagar, D. Whole genome comparison of Aspergillus flavus L-morphotype strain NRRL 3357 (type) and S-morphotype strain AF70. PLoS ONE 2018, 13, e0199169. [Google Scholar] [CrossRef]
- Etxebeste, O.; Otamendi, A.; Garzia, A.; Espeso, E.A.; Cortese, M.S. Rewiring of transcriptional networks as a major event leading to the diversity of asexual multicellularity in fungi. Crit. Rev. Microbiol. 2019, 45, 548–563. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Lange, L.; Lacey, A.E.; Vuong, D.; Midgley, D.J.; Greenfield, P.; Bradbury, M.; Lacey, E.; Busk, P.K.; Pilgaard, B.; et al. Aspergillus hancockii sp. nov., a biosynthetically talented fungus endemic to southeastern Australian soils. PLoS ONE 2017, 12, e0170254. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.U.; Kim, K.M.; Choi, Y.-H.; Hurh, B.-S.; Lee, I. Whole genome analysis of Aspergillus sojae SMF 134 supports its merits as a starter for soybean fermentation. J. Microbiol. 2019, 57, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Pei, H.; Zhou, X.; Zhao, K.; Yu, M.; Han, G.; Fan, J.; Tao, F. Systematic characterization of bZIP transcription factors required for development and aflatoxin generation by high-throughput gene knockout in Aspergillus flavus. J. Fungi 2022, 8, 356. [Google Scholar] [CrossRef] [PubMed]
- Fasoyin, O.E.; Wang, B.; Qiu, M.; Han, X.; Chung, K.-R.; Wang, S. Carbon catabolite repression gene creA regulates morphology, aflatoxin biosynthesis and virulence in Aspergillus flavus. Fungal Genet. Biol. 2018, 115, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-Y.; Son, Y.-E.; Lee, D.-H.; Eom, T.-J.; Kim, M.-J.; Park, H.-S. Function of crzA in fungal development and aflatoxin production in Aspergillus flavus. Toxins 2019, 11, 567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, R.; Yang, K.; Tumukunde, E.; Guo, Z.; Zhang, B.; Liu, Y.; Zhuang, Z.; Yuan, J.; Wang, S. Regulator of G protein signaling contributes to the development and aflatoxin biosynthesis in Aspergillus flavus through the regulation of Gα Activity. Appl. Environ. Microbiol. 2022, 88, e00244-22. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Scharfenstein, L.L.; Mack, B.; Ehrlich, K.C. Deletion of the Aspergillus flavus orthologue of A. nidulans fluG reduces conidiation and promotes production of sclerotia but does not abolish aflatoxin biosynthesis. Appl. Environ. Microbiol. 2012, 78, 7557–7563. [Google Scholar] [CrossRef] [PubMed]
- Tumukunde, E.; Li, D.; Qin, L.; Li, Y.; Shen, J.; Wang, S.; Yuan, J. Osmotic-adaptation response of sakA/hogA gene to aflatoxin biosynthesis, morphology development and pathogenicity in Aspergillus flavus. Toxins 2019, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Distinct roles for VeA and LaeA in development and pathogenesis of Aspergillus flavus. Eukaryot. Cell 2009, 8, 1051–1060. [Google Scholar] [CrossRef]
- Chang, P.-K.; Scharfenstein, L.L.; Luo, M.; Mahoney, N.; Molyneux, R.J.; Yu, J.; Brown, R.L.; Campbell, B.C. Loss of msnA, a putative stress regulatory gene, in Aspergillus parasiticus and Aspergillus flavus increased production of conidia, aflatoxins and kojic acid. Toxins 2011, 3, 82–104. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.; Lohmar, J.M.; Satterlee, T.; Cary, J.W.; Calvo, A.M. The master transcription factor mtfA governs aflatoxin production, morphological development and pathogenicity in the fungus Aspergillus flavus. Toxins 2016, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Harris-Coward, P.Y.; Ehrlich, K.C.; Mack, B.M.; Kale, S.P.; Larey, C.; Calvo, A.M. NsdC and NsdD affect Aspergillus flavus morphogenesis and aflatoxin production. Eukaryot. Cell 2012, 11, 1104–1111. [Google Scholar] [CrossRef]
- Yuan, X.-Y.; Li, J.-Y.; Zhi, Q.-Q.; Chi, S.-D.; Qu, S.; Luo, Y.-F.; He, Z.-M. SfgA renders Aspergillus flavus more stable to the external environment. J. Fungi 2022, 8, 638. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Zhang, F.; Nie, X.; Wang, X.; Yuan, J.; Zhuang, Z.; Wang, S. Essential APSES transcription factors for mycotoxin synthesis, fungal development, and pathogenicity in Aspergillus flavus. Front. Microbiol. 2017, 8, 2277. [Google Scholar] [CrossRef] [PubMed]
- Eom, T.-J.; Moon, H.; Yu, J.-H.; Park, H.-S. Characterization of the velvet regulators in Aspergillus flavus. J. Microbiol. 2018, 56, 893–901. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Mead, M.E.; Kim, S.-C.; Rokas, A.; Yu, J.-H. WetA bridges cellular and chemical development in Aspergillus flavus. PLoS ONE 2017, 12, e0179571. [Google Scholar] [CrossRef]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef]
- Krijgsheld, P.; Bleichrodt, R.; van Veluw, G.; Wang, F.; Müller, W.; Dijksterhuis, J.; Wösten, H. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Boylan, M.T.; Mirabito, P.M.; Willett, C.E.; Zimmerman, C.R.; Timberlake, W.E. Isolation and physical characterization of three essential conidiation genes from Aspergillus nidulans. Mol. Cell. Biol. 1987, 7, 3113–3118. [Google Scholar] [CrossRef] [PubMed]
- Shin, K.-S.; Kim, Y.H.; Yu, J.-H. Proteomic analyses reveal the key roles of BrlA and AbaA in biogenesis of gliotoxin in Aspergillus fumigatus. Biochem. Biophys. Res. Commun. 2015, 463, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Yu, J.-H. AbaA and WetA govern distinct stages of Aspergillus fumigatus development. Microbiology 2011, 157, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-H.; Mah, J.-H.; Seo, J.-A. Growth and developmental control in the model and pathogenic Aspergilli. Eukaryot. Cell 2006, 5, 1577–1584. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Mead, M.E.; Lee, M.-K.; Neuhaus, G.F.; Adpressa, D.A.; Martien, J.I.; Son, Y.-E.; Moon, H.; Amador-Noguez, D.; Han, K.-H.; et al. Transcriptomic, protein-DNA interaction, and metabolomic studies of VosA, VelB, and WetA in Aspergillus nidulans asexual spores. mBio 2021, 12, e03128-20. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.-Y.; Mead, M.; Lee, M.-K.; Loss, E.M.O.; Kim, S.-C.; Rokas, A.; Yu, J.-H. Systematic dissection of the evolutionarily conserved WetA developmental regulator across a genus of filamentous fungi. mBio 2018, 9, e01130-18. [Google Scholar] [CrossRef] [PubMed]
- Clutterbuck, A.J. A Mutational analysis of conidial development in Aspergillus nidulans. Genetics 1969, 63, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Tokuoka, M.; Jin, F.J.; Takahashi, T.; Koyama, Y. Genetic analysis of conidiation regulatory pathways in koji-mold Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 10–18. [Google Scholar] [CrossRef]
- Yamada, O.; Lee, B.R.; Gomi, K.; Iimura, Y. Cloning and functional analysis of the Aspergillus oryzae conidiation regulator gene brlA by its disruption and misscheduled expression. J. Biosci. Bioeng. 1999, 87, 424–429. [Google Scholar] [CrossRef]
- Prade, R.; Timberlake, W. The Aspergillus nidulans brlA regulatory locus consists of overlapping transcription units that are individually required for conidiophore development. EMBO J. 1993, 12, 2439–2447. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Timberlake, W.E. Identification of Aspergillus brlA response elements (BREs) by genetic selection in yeast. Genetics 1993, 133, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Etxebeste, O. Transcription factors in the fungus Aspergillus nidulans: Markers of genetic innovation, network rewiring and conflict between genomics and transcriptomics. J. Fungi 2021, 7, 600. [Google Scholar] [CrossRef]
- Twumasi-Boateng, K.; Yu, Y.; Chen, D.; Gravelat, F.N.; Nierman, W.C.; Sheppard, D.C. Transcriptional profiling identifies a role for BrlA in the response to nitrogen depletion and for StuA in the regulation of secondary metabolite clusters in Aspergillus fumigatus. Eukaryot. Cell 2009, 8, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Sewall, T.C.; Mims, C.W.; Timberlake, W.E. abaA controls phialide differentiation in Aspergillus nidulans. Plant Cell 1990, 2, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Andrianopoulos, A.; Timberlake, W.E. The Aspergillus nidulans abaA gene encodes a transcriptional activator that acts as a genetic switch to control development. Mol. Cell. Biol. 1994, 14, 2503–2515. [Google Scholar] [CrossRef] [PubMed]
- Aramayo, R.; Timberlake, W. The Aspergillus nidulans yA gene is regulated by abaA. EMBO J. 1993, 12, 2039–2048. [Google Scholar] [CrossRef]
- Mead, M.E.; Borowsky, A.T.; Joehnk, B.; Steenwyk, J.L.; Shen, X.-X.; Sil, A.; Rokas, A. Recurrent loss of abaA, a master regulator of asexual development in filamentous fungi, correlates with changes in genomic and morphological traits. Genome Biol. Evol. 2020, 12, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Marshall, M.A.; Timberlake, W.E. Aspergillus nidulans wetA activates spore-specific gene expression. Mol. Cell. Biol. 1991, 11, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Noble, L.M.; Andrianopoulos, A. Reproductive competence: A recurrent logic module in eukaryotic development. Proc. R. Soc. B Boil. Sci. 2013, 280, 20130819. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, D.; Gealt, M.; Pastushok, M. Gene control of developmental competence in Aspergillus nidulans. Dev. Biol. 1973, 34, 9–15. [Google Scholar] [CrossRef]
- Etxebeste, O.; Garzia, A.; Espeso, E.A.; Ugalde, U. Aspergillus nidulans asexual development: Making the most of cellular modules. Trends Microbiol. 2010, 18, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Wieser, J.; Lee, B.N.; Fondon, J.W.; Adams, T.H. Genetic requirements for initiating asexual development in Aspergillus nidulans. Curr. Genet. 1994, 27, 62–69. [Google Scholar] [CrossRef]
- Lee, B.N.; Adams, T.H. The Aspergillus nidulans fluG gene is required for production of an extracellular developmental signal and is related to prokaryotic glutamine synthetase I. Genes Dev. 1994, 8, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.N.; Adams, T.H. FluG and flbA function interdependently to initiate conidiophore development in Aspergillus nidulans through brlA beta activation. EMBO J. 1996, 15, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.I.; García, L.T.; Cortese, M.; Ugalde, U. The Early asexual development regulator fluG codes for a putative bifunctional enzyme. Front. Microbiol. 2019, 10, 778. [Google Scholar] [CrossRef]
- Lo, H.-C.; Entwistle, R.; Guo, C.-J.; Ahuja, M.; Szewczyk, E.; Hung, J.-H.; Chiang, Y.-M.; Oakley, B.R.; Wang, C.C.C. Two separate gene clusters encode the biosynthetic pathway for the meroterpenoids austinol and dehydroaustinol in Aspergillus nidulans. J. Am. Chem. Soc. 2012, 134, 4709–4720. [Google Scholar] [CrossRef]
- Rodríguez-Urra, A.B.; Jiménez, C.; Nieto, M.I.; Rodríguez, J.; Hayashi, H.; Ugalde, U. Signaling the induction of sporulation involves the interaction of two secondary metabolites in Aspergillus nidulans. ACS Chem. Biol. 2012, 7, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Scharfenstein, L.L.; Li, P.; Ehrlich, K.C. Aspergillus flavus VelB acts distinctly from VeA in conidiation and may coordinate with FluG to modulate sclerotial production. Fungal Genet. Biol. 2013, 58-59, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Na Lee, B.; Adams, T.H. Overexpression offIbA, an early regulator of Aspergillus asexual sporulation, leads to activation of brIA and premature initiation of development. Mol. Microbiol. 1994, 14, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Kim, M.-J.; Yu, J.-H.; Shin, K.-S. Heterotrimeric G-protein signalers and RGSs in Aspergillus fumigatus. Pathogens 2020, 9, 902. [Google Scholar] [CrossRef]
- Etxebeste, O.; Ni, M.; Garzia, A.; Kwon, N.-J.; Fischer, R.; Yu, J.-H.; Espeso, E.A.; Ugalde, U. Basic-zipper-type transcription factor FlbB controls asexual development in Aspergillus nidulans. Eukaryot. Cell 2008, 7, 38–48. [Google Scholar] [CrossRef]
- Garzia, A.; Etxebeste, O.; Herrero-Garcia, E.; Fischer, R.; Espeso, E.A.; Ugalde, U. Aspergillus nidulansFlbE is an upstream developmental activator of conidiation functionally associated with the putative transcription factor FlbB. Mol. Microbiol. 2008, 71, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Arratia-Quijada, J.; Sánchez, O.; Scazzocchio, C.; Aguirre, J. FlbD, a Myb transcription factor of Aspergillus nidulans, is uniquely involved in both asexual and sexual differentiation. Eukaryot. Cell 2012, 11, 1132–1142. [Google Scholar] [CrossRef]
- Kwon, N.-J.; Garzia, A.; Espeso, E.A.; Ugalde, U.; Yu, J.-H. FlbC is a putative nuclear C2H2 transcription factor regulating development in Aspergillus nidulans. Mol. Microbiol. 2010, 77, 1203–1219. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Shin, K.-S.; Wang, T.; Yu, J.-H. Aspergillus fumigatus flbB encodes two basic leucine zipper domain (bZIP) proteins required for proper asexual development and gliotoxin production. Eukaryot. Cell 2010, 9, 1711–1723. [Google Scholar] [CrossRef] [PubMed]
- Otamendi, A.; Perez-De-Nanclares-Arregi, E.; Oiartzabal-Arano, E.; Cortese, M.S.; Espeso, E.A.; Etxebeste, O. Developmental regulators FlbE/D orchestrate the polarity site-to-nucleus dynamics of the fungal bZIP transcription factor FlbB. Experientia 2019, 76, 4369–4390. [Google Scholar] [CrossRef]
- Herrero-Garcia, E.; Perez-De-Nanclares-Arregi, E.; Cortese, M.; Markina-Iñarrairaegui, A.; Oiartzabal-Arano, E.; Etxebeste, O.; Ugalde, U.; Espeso, E.A. Tip-to-nucleus migration dynamics of the asexual development regulator FlbB in vegetative cells. Mol. Microbiol. 2015, 98, 607–624. [Google Scholar] [CrossRef] [PubMed]
- Bayram, Ö.; Braus, G.H. Coordination of secondary metabolism and development in fungi: The velvet family of regulatory proteins. FEMS Microbiol. Rev. 2012, 36, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Yu, J.-H. Velvet regulators in Aspergillus spp. Microbiol. Biotechnol. Lett. 2016, 44, 409–419. [Google Scholar] [CrossRef]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.-J.; Keller, N.P.; Yu, J.-H.; et al. VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef] [PubMed]
- Bayram, S.; Bayram, Ö.; Valerius, O.; Park, H.S.; Irniger, S.; Gerke, J.; Ni, M.; Han, K.-H.; Yu, J.-H.; Braus, G.H. LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity. PLoS Genet. 2010, 6, e1001226. [Google Scholar] [CrossRef]
- Kim, H.-S.; Han, K.-Y.; Kim, K.-J.; Han, D.-M.; Jahng, K.-Y.; Chae, K.-S. The veA gene activates sexual development in Aspergillus nidulans. Fungal Genet. Biol. 2002, 37, 72–80. [Google Scholar] [CrossRef]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.; Bok, J.; Chung, D.; Perrin, R.M.; Shwab, E.K. LaeA, a global regulator of Aspergillus toxins. Med Mycol. 2006, 44, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.-K.; Scharfenstein, L.L.; Ehrlich, K.C.; Wei, Q.; Bhatnagar, D.; Ingber, B.F. Effects of laeA deletion on Aspergillus flavus conidial development and hydrophobicity may contribute to loss of aflatoxin production. Fungal Biol. 2011, 116, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Lara-Rojas, F.; Sánchez, O.; Kawasaki, L.; Aguirre, J. Aspergillus nidulans transcription factor AtfA interacts with the MAPK SakA to regulate general stress responses, development and spore functions. Mol. Microbiol. 2011, 80, 436–454. [Google Scholar] [CrossRef] [PubMed]
- Leiter, É.; Emri, T.; Pákozdi, K.; Hornok, L.; Pócsi, I. The impact of bZIP Atf1ortholog global regulators in fungi. Appl. Microbiol. Biotechnol. 2021, 105, 5769–5783. [Google Scholar] [CrossRef]
- Garrido-Bazán, V.; Jaimes-Arroyo, R.; Sánchez, O.; Lara-Rojas, F.; Aguirre, J. SakA and MpkC stress MAPKs show opposite and common functions during stress responses and development in Aspergillus nidulans. Front. Microbiol. 2018, 9, 2518. [Google Scholar] [CrossRef]
- Bailey, C.; Arst, H.N. Carbon Catabolite Repression in Aspergillus nidulans. JBIC J. Biol. Inorg. Chem. 1975, 51, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon Catabolite Repression in Filamentous Fungi. Int. J. Mol. Sci. 2017, 19, 48. [Google Scholar] [CrossRef]
- Ries, L.N.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse Regulation of the CreA Carbon Catabolite Repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Lee, S.C.; Cardenas, M.E.; Heitman, J. Calcium-calmodulin-calcineurin signaling: A globally conserved virulence cascade in eukaryotic microbial pathogens. Cell Host Microbe 2019, 26, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Cramer, R.A.; Perfect, B.Z.; Pinchai, N.; Park, S.; Perlin, D.S.; Asfaw, Y.G.; Heitman, J.; Perfect, J.R.; Steinbach, W.J. Calcineurin target CrzA regulates conidial germination, hyphal growth, and pathogenesis of Aspergillus fumigatus. Eukaryot. Cell 2008, 7, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Juvvadi, P.R.; Lamoth, F.; Steinbach, W.J. Calcineurin as a multifunctional regulator: Unraveling novel functions in fungal stress responses, hyphal growth, drug resistance, and pathogenesis. Fungal Biol. Rev. 2014, 28, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Spielvogel, A.; Findon, H.; Arst, H.N.; Araújo-Bazán, L.; Hernández-Ortíz, P.; Stahl, U.; Meyer, V.; Espeso, E.A. Two zinc finger transcription factors, CrzA and SltA, are involved in cation homoeostasis and detoxification in Aspergillus nidulans. Biochem. J. 2008, 414, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ortiz, P.; Espeso, E.A. Spatiotemporal dynamics of the calcineurin target CrzA. Cell. Signal. 2017, 29, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Cramer, R.A.; Perfect, B.Z.; Asfaw, Y.G.; Sauer, T.C.; Najvar, L.K.; Kirkpatrick, W.R.; Patterson, T.F.; Benjamin, D.K.; Heitman, J.; et al. Calcineurin controls growth, morphology, and pathogenicity in Aspergillus fumigatus. Eukaryot. Cell 2006, 5, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- De Nadal, E.; Posas, F. The HOG pathway and the regulation of osmoadaptive responses in yeast. FEMS Yeast Res. 2022, 22, foac013. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Li, R. Current Understanding of HOG-MAPK pathway in Aspergillus fumigatus. Mycopathologia 2012, 175, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.A.; Morgan, B.A.; Quinn, J. Stress signalling to fungal stress-activated protein kinase pathways. FEMS Microbiol. Lett. 2010, 306, 1–8. [Google Scholar] [CrossRef]
- Emri, T.; Szarvas, V.; Orosz, E.; Antal, K.; Park, H.; Han, K.-H.; Yu, J.-H.; Pócsi, I. Core oxidative stress response in Aspergillus nidulans. BMC Genom. 2015, 16, 478. [Google Scholar] [CrossRef]
- Ramamoorthy, V.; Dhingra, S.; Kincaid, A.; Shantappa, S.; Feng, X.; Calvo, A.M. The putative C2H2 transcription factor MtfA is a novel regulator of secondary metabolism and morphogenesis in Aspergillus nidulans. PLoS ONE 2013, 8, e74122. [Google Scholar] [CrossRef]
- Smith, T.D.; Calvo, A.M. The mtfA transcription factor gene controls morphogenesis, gliotoxin production, and virulence in the opportunistic human pathogen Aspergillus fumigatus. Eukaryot. Cell 2014, 13, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-R.; Chae, K.-S.; Han, K.-H.; Han, D.-M. The nsdC Gene Encoding a Putative C2H2-Type Transcription Factor Is a Key Activator of Sexual Development in Aspergillus nidulans. Genetics 2009, 182, 771–783. [Google Scholar] [CrossRef]
- Han, K.; Yu, J.; Chae, K.; Jahng, K. The nsdD gene encodes a putative GATA-type transcription factor necessary for sexual development of Aspergillus nidulans. Mol. Microbiol. 2001, 41, 299–309. [Google Scholar] [CrossRef]
- Lee, M.-K.; Kwon, N.-J.; Lee, I.-S.; Jung, S.; Kim, S.-C.; Yu, J.-H. Negative regulation and developmental competence in Aspergillus. Sci. Rep. 2016, 6, 28874. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.K.; Mack, B.M.; Wei, Q.; Bland, J.; Bhatnagar, D.; Cary, J.W. RNA sequencing of an nsdC mutant reveals global regulation of secondary metabolic gene clusters in Aspergillus flavus. Microbiol. Res. 2016, 182, 150–161. [Google Scholar] [CrossRef]
- Seo, J.-A.; Guan, Y.; Yu, J.-H. Suppressor Mutations Bypass the Requirement of fluG for Asexual Sporulation and Sterigmatocystin Production in Aspergillus nidulans. Genetics 2003, 165, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.-A.; Guan, Y.; Yu, J.-H. FluG-Dependent Asexual Development in Aspergillus nidulans Occurs via Derepression. Genetics 2006, 172, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.Y.; Toennis, T.M.; Adams, T.H.; Miller, B.L. Isolation and transcriptional characterization of a morphological modifier: The Aspergillus nidulans stunted (stuA) gene. Mol. Gen. Genet. MGG 1991, 227, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.Y.; Wu, J.; Miller, B.L. StuA is required for cell pattern formation in Aspergillus. Genes Dev. 1992, 6, 1770–1782. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.C.; Doedt, T.; Chiang, L.Y.; Kim, H.; Chen, D.; Nierman, W.C.; Filler, S.G. The Aspergillus fumigatus StuA protein governs the up-regulation of a discrete transcriptional program during the acquisition of developmental competence. Mol. Biol. Cell 2005, 16, 5866–5879. [Google Scholar] [CrossRef]
- Wang, P.; Xu, J.; Chang, P.-K.; Liu, Z.; Kong, Q. New Insights of transcriptional regulator AflR in Aspergillus flavus physiology. Microbiol. Spectr. 2022, 10, e00791-21. [Google Scholar] [CrossRef]
- Han, X.; Qiu, M.; Wang, B.; Yin, W.-B.; Nie, X.; Qin, Q.; Ren, S.; Yang, K.; Zhang, F.; Zhuang, Z.; et al. Functional analysis of the nitrogen metabolite repression regulator gene nmrA in Aspergillus flavus. Front. Microbiol. 2016, 7, 1794. [Google Scholar] [CrossRef] [PubMed]
- Mengjuan, Z.; Guanglan, L.; Xiaohua, P.; Weitao, S.; Can, T.; Xuan, C.; Yanling, Y.; Zhenhong, Z. The PHD transcription factor Cti6 is involved in the fungal colonization and aflatoxin B1 biological synthesis of Aspergillus flavus. IMA Fungus 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Harris-Coward, P.; Scharfenstein, L.; Mack, B.M.; Chang, P.-K.; Wei, Q.; Lebar, M.; Carter-Wientjes, C.; Majumdar, R.; Mitra, C.; et al. The Aspergillus flavus homeobox gene, hbx1, is required for development and aflatoxin production. Toxins 2017, 9, 315. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Wiemann, P.; Garvey, G.S.; Lim, F.Y.; Haas, B.; Wortman, J.; Keller, N.P. Illumina identification of RsrA, a conserved C2H2 transcription factor coordinating the NapA mediated oxidative stress signaling pathway in Aspergillus. BMC Genom. 2014, 15, 1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Yang, G.; Zhang, D.; Liu, Y.; Li, Y.; Lin, G.; Guo, Z.; Wang, S.; Zhuang, Z. The PHD transcription factor Rum1 regulates morphogenesis and aflatoxin biosynthesis in Aspergillus flavus. Toxins 2018, 10, 301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xu, G.; Geng, L.; Lu, X.; Yang, K.; Yuan, J.; Nie, X.; Zhuang, Z.; Wang, S. The stress response regulator AflSkn7 influences morphological development, stress response, and pathogenicity in the fungus Aspergillus flavus. Toxins 2016, 8, 202. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.-E.; Cho, H.-J.; Lee, M.-K.; Park, H.-S. Characterizing the role of Zn cluster family transcription factor ZcfA in governing development in two Aspergillus species. PLoS ONE 2020, 15, e0228643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
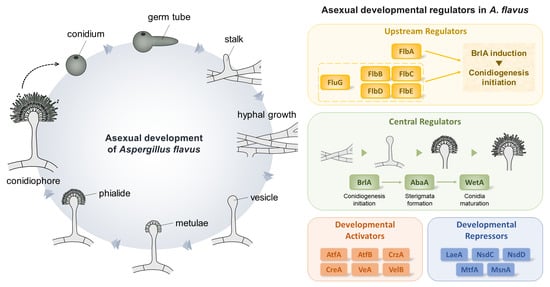
Figure 1.
Asexual developmental regulators in Aspergillus spp. (A) A morphogenic model of conidiophore development of A. flavus. (B) A genetic model of the regulation of conidiogenesis in A. flavus. (C) Distribution of regulators involved in conidiogenesis in Aspergillus section Flavi. Distribution of 34 important regulators were investigated in 24 representative genomes from the section Flavi. The genomic data of A. albertensis, A. alliaceus CBS 536.65, A. arachidicola, A. avenaceus IBT 18842, A. bertholletius IBT 29228, A. bombycis NRRL 26010, A. caelatus CBS 763.97, A. coremiiformis CBS 553.77, A. flavus NRRL3357, A. leporis CBS 151.66, A. minisclerotigenes CBS 117635, A. nomius IBT 12657, A. novoparasiticus CBS 126849, A. oryzae RIB40, A. parasiticus CBS 117618, A. parvisclerotigenus CBS 121.62, A. pseudocaelatus CBS 117616, A. pseudonomius CBS 119388, A. pseudotamarii CBS 117625, A. sergii CBS 130017, A. tamarii CBS 117626, and A. transmontanensis CBS 130015 are obtained from Joint Genome Institute fungal genome portal MycoCosm (http://jgi.doe.gov/fungi, accessed on 4 August 2022). The genomic data of A. hancockii and A. sojae SMF134 were previously published [29,30]. The homologs were searched by BlastP using the regulators of A. flavus NRRL3357 as queries.
Figure 1.
Asexual developmental regulators in Aspergillus spp. (A) A morphogenic model of conidiophore development of A. flavus. (B) A genetic model of the regulation of conidiogenesis in A. flavus. (C) Distribution of regulators involved in conidiogenesis in Aspergillus section Flavi. Distribution of 34 important regulators were investigated in 24 representative genomes from the section Flavi. The genomic data of A. albertensis, A. alliaceus CBS 536.65, A. arachidicola, A. avenaceus IBT 18842, A. bertholletius IBT 29228, A. bombycis NRRL 26010, A. caelatus CBS 763.97, A. coremiiformis CBS 553.77, A. flavus NRRL3357, A. leporis CBS 151.66, A. minisclerotigenes CBS 117635, A. nomius IBT 12657, A. novoparasiticus CBS 126849, A. oryzae RIB40, A. parasiticus CBS 117618, A. parvisclerotigenus CBS 121.62, A. pseudocaelatus CBS 117616, A. pseudonomius CBS 119388, A. pseudotamarii CBS 117625, A. sergii CBS 130017, A. tamarii CBS 117626, and A. transmontanensis CBS 130015 are obtained from Joint Genome Institute fungal genome portal MycoCosm (http://jgi.doe.gov/fungi, accessed on 4 August 2022). The genomic data of A. hancockii and A. sojae SMF134 were previously published [29,30]. The homologs were searched by BlastP using the regulators of A. flavus NRRL3357 as queries.

Figure 2.
Summary of the central regulators in A. flavus. (A) Phenotypes of ΔbrlA, ΔabaA, and ΔwetA mutant strains. Colony phenotypes of ΔbrlA, ΔabaA, and ΔwetA strains point-inoculated on solid glucose minimal medium with 0.1% yeast extract media and grown at 37 °C (Upper). Morphology of ΔbrlA, ΔabaA, and ΔwetA conidiophores observed under a light microscope at 48 h after inoculation onto solid MMYE media at 37 °C (Bottom). (B) Scanning electron micrographs of ΔbrlA and ΔabaA strains. (C) mRNA levels of brlA, abaA, and wetA during A. flavus life cycle. Samples for RT-qPCR analysis were collected from 12, 18, and 24 h of vegetative growth; 6, 12, 24, and 48 h of asexual development, and in conidia.
Figure 2.
Summary of the central regulators in A. flavus. (A) Phenotypes of ΔbrlA, ΔabaA, and ΔwetA mutant strains. Colony phenotypes of ΔbrlA, ΔabaA, and ΔwetA strains point-inoculated on solid glucose minimal medium with 0.1% yeast extract media and grown at 37 °C (Upper). Morphology of ΔbrlA, ΔabaA, and ΔwetA conidiophores observed under a light microscope at 48 h after inoculation onto solid MMYE media at 37 °C (Bottom). (B) Scanning electron micrographs of ΔbrlA and ΔabaA strains. (C) mRNA levels of brlA, abaA, and wetA during A. flavus life cycle. Samples for RT-qPCR analysis were collected from 12, 18, and 24 h of vegetative growth; 6, 12, 24, and 48 h of asexual development, and in conidia.

Figure 3.
Roles of the central regulators in A. flavus sclerotia and aflatoxin production. (A) Ethanol-washed colony photographs of ΔbrlA, ΔabaA, and ΔwetA strains grown on solid minimal media with 1% glucose and 0.1% yeast extract (MMYE) for 7 days. Quantitative analysis of sclerotia of these strains. (B) Image of thin-layer chromatography (TLC) of aflatoxin B1 from ΔbrlA, ΔabaA, and ΔwetA strains under dark conditions. To extract aflatoxin B1 from each strain, about 107 conidia were inoculated into liquid complete media and incubated for 7 days at 30°C in dark condition. To extract aflatoxin B1, chloroform was used. The samples were spotted onto a TLC silica plate, and the plate was placed into a chamber containing chloroform: acetone (9:1, v/v). Densitometry of the TLC analysis results. Statistical differences between control and mutant strains were evaluated using Student’s unpaired t-tests. Data are reported as the mean ± standard deviation. ** p < 0.01, *** p < 0.001.
Figure 3.
Roles of the central regulators in A. flavus sclerotia and aflatoxin production. (A) Ethanol-washed colony photographs of ΔbrlA, ΔabaA, and ΔwetA strains grown on solid minimal media with 1% glucose and 0.1% yeast extract (MMYE) for 7 days. Quantitative analysis of sclerotia of these strains. (B) Image of thin-layer chromatography (TLC) of aflatoxin B1 from ΔbrlA, ΔabaA, and ΔwetA strains under dark conditions. To extract aflatoxin B1 from each strain, about 107 conidia were inoculated into liquid complete media and incubated for 7 days at 30°C in dark condition. To extract aflatoxin B1, chloroform was used. The samples were spotted onto a TLC silica plate, and the plate was placed into a chamber containing chloroform: acetone (9:1, v/v). Densitometry of the TLC analysis results. Statistical differences between control and mutant strains were evaluated using Student’s unpaired t-tests. Data are reported as the mean ± standard deviation. ** p < 0.01, *** p < 0.001.

Figure 4.
Roles of the fluffy genes in A. flavus. (A) Colony morphology of ΔflbB, ΔflbC, ΔflbD, and ΔflbE mutant strains. (B) Quantitative analysis of the number of conidia per plate shown in (A). (C) mRNA level of brlA in wild-type and mutant strains. (D) Ethanol-washed colony photographs of fluffy mutant strains. (E) Image of TLC of aflatoxin B1 from fluffy mutant strains. Densitometry of TLC analysis results. Statistical differences between control and mutant strains were evaluated using Student’s unpaired t-tests. Data are reported as the mean ± standard deviation. * p < 0.05, ** p < 0.01, and *** p < 0.001.
Figure 4.
Roles of the fluffy genes in A. flavus. (A) Colony morphology of ΔflbB, ΔflbC, ΔflbD, and ΔflbE mutant strains. (B) Quantitative analysis of the number of conidia per plate shown in (A). (C) mRNA level of brlA in wild-type and mutant strains. (D) Ethanol-washed colony photographs of fluffy mutant strains. (E) Image of TLC of aflatoxin B1 from fluffy mutant strains. Densitometry of TLC analysis results. Statistical differences between control and mutant strains were evaluated using Student’s unpaired t-tests. Data are reported as the mean ± standard deviation. * p < 0.05, ** p < 0.01, and *** p < 0.001.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The function of the major developmental regulators in studied A. flavus conidiation.
| Genes | Domain(s) | Description | Ref. |
|---|---|---|---|
| AbaA | TEA/ATTS | Regulator for sterigmata formation | - |
| AtfA | bZIP | Developmental activator | [31] |
| AtfB | bZIP | Developmental activator | [31] |
| BrlA | C2H2 zinc finger | Initiator for conidiogenesis | - |
| CreA | C2H2 zinc finger | Developmental activator | [32] |
| CrzA | C2H2 zinc finger | Developmental activator | [33] |
| FlbA | RGS | Upstream developmental activator | [34] |
| FlbB | Basic leucine zipper | Upstream developmental activator | - |
| FlbC | C2H2 zinc finger | Upstream dev. activator | - |
| FlbD | Myb-like DNA binding | Upstream dev. activator | - |
| FlbE | - | Upstream dev. activator | - |
| FluG | Amidohydrolase and GS | Upstream dev. activator | [35] |
| HogA | Protein kinase | Modulator for conidiation | [36] |
| LaeA | SAM | Developmental repressor | [37] |
| MsnA | C2H2 zinc finger | Developmental repressor | [38] |
| MtfA | C2H2 zinc finger | Developmental repressor | [39] |
| NsdC | C2H2 zinc finger | Developmental repressor | [40] |
| NsdD | GATA-type zinc-finger | Developmental repressor | [40] |
| SfgA | Zn2Cys6 | Developmental repressor | [41] |
| StuA | APSES | Modulator for conidiophore formation | [42] |
| VeA | Velvet | Developmental activator | [37] |
| VelB | Velvet | Developmental activator Regulator for conidial maturation | [43] |
| VelC | Velvet | - | [43] |
| VelD | Velvet | - | [43] |
| VosA | Velvet | Regulator for conidial maturation | [43] |
| WetA | ESC1/WetA-related | Regulator for conidial maturation | [44] |
bZIP, basic leucine zipper; RGS, regulator of G protein signaling; SAM, S-adenosyl-l-methionine; GS, glutamine synthetase; and APSES, Asm1p, Phd1p, Sok2p, Efg1p, and StuAp.
Table 2.
Comparison of the developmental phenotypes of the Δflb mutant strains in A. nidulans and A. flavus.
Table 2.
Comparison of the developmental phenotypes of the Δflb mutant strains in A. nidulans and A. flavus.
| Genes | Asexual Development | Sexual Structure Formation | Ref. | ||
|---|---|---|---|---|---|
| Conidia Formation | brlA Expression | ||||
| FlbB | ΔAniflbB | decrease | absence | not determined | [77] |
| ΔAflflbB | decrease | decrease | decrease | - | |
| FlbC | ΔAniflbC | decrease | delay | increase | [80] |
| ΔAflflbC | decrease | decrease | decrease | - | |
| FlbD | ΔAniflbD | decrease | delay | decrease | [79] |
| ΔAflflbD | decrease | decrease | decrease | - | |
| FlbE | ΔAniflbE | decrease | delay | not determined | [78] |
| ΔAflflbE | decrease | decrease | decrease | - | |
Table 3.
The function of transcription factors related to A. flavus conidiation.
| Genes | Domain(s) | Phenotype of Deletion Mutant | Ref. |
|---|---|---|---|
| AflR | Zn2Cys6 domain | Decrease conidiophore production | [119] |
| AreA | GATA zinc finger domain | Decrease conidiophore production and brlA expression | [120] |
| Cti6 | PHD domain | Decrease conidiophore production and brlA expression | [121] |
| Hbx1 | Homeodomain | Loss of conidiophore, decrease brlA expression | [122] |
| RafA | APSES | Decrease conidiophore production, increase brlA expression | [42] |
| RsrA | C2H2 zinc finger | Decrease conidiophore production | [123] |
| Rum1 | PHD domain | Increase conidiophore production and brlA expression | [124] |
| Skn7 | Heat-shock transcription factor-like DNA-binding domain | Abnormal conidiophore, decrease conidiophore production | [125] |
| ZcfA | Zn2Cys6 domain | Increase conidiophore production | [126] |
PHD, plant homeodomain; APSES, Asm1p, Phd1p, Sok2p, Efg1p, and StuAp.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cho, H.-J.; Son, S.-H.; Chen, W.; Son, Y.-E.; Lee, I.; Yu, J.-H.; Park, H.-S. Regulation of Conidiogenesis in Aspergillus flavus. Cells 2022, 11, 2796. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11182796
AMA Style
Cho H-J, Son S-H, Chen W, Son Y-E, Lee I, Yu J-H, Park H-S. Regulation of Conidiogenesis in Aspergillus flavus. Cells. 2022; 11(18):2796. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11182796
Chicago/Turabian StyleCho, He-Jin, Sung-Hun Son, Wanping Chen, Ye-Eun Son, Inhyung Lee, Jae-Hyuk Yu, and Hee-Soo Park. 2022. "Regulation of Conidiogenesis in Aspergillus flavus" Cells 11, no. 18: 2796. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11182796
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.