
Increased Numbers of Enteric Glial Cells in the Peyer’s Patches and Enhanced Intestinal Permeability by Glial Cell Mediators in Patients with Ileal Crohn’s Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. Quantification of GFAP and S100β by Immunofluorescence
2.3. Western Blotting for GFAP and S100β
2.4. Ussing Chamber Experiments with EGC Mediators
2.5. Statistical Analysis
3. Results
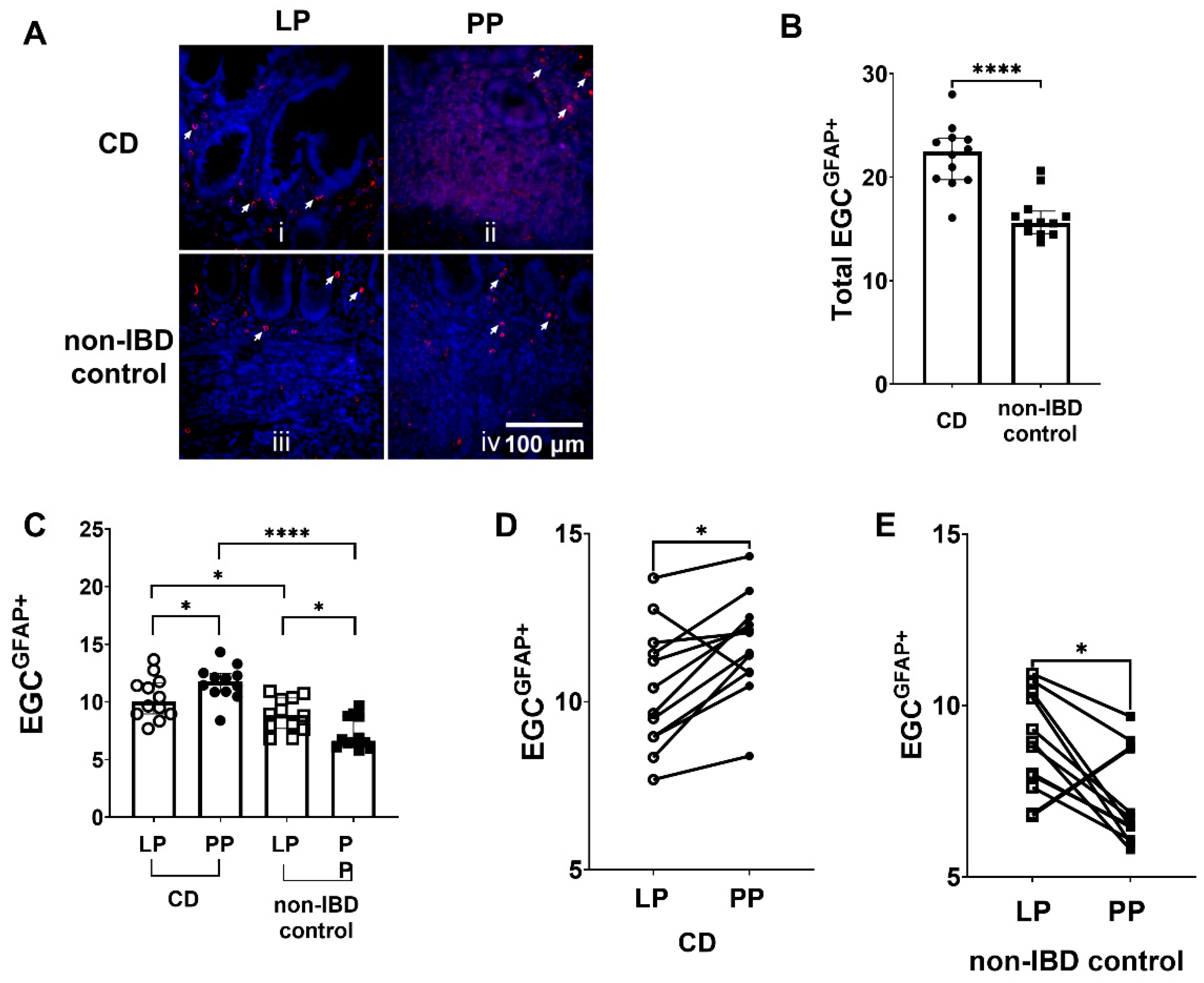
3.1. Increased Numbers of EGCGFAP+ in CD Patients Compared to Non-IBD Controls
3.2. Differences in EGCGFAP+ Distribution between Peyer’s Patches and the Lamina Propria in CD Patients and Non-IBD Controls, but Also between CD Patients and Non-IBD Groups
3.3. Western Blotting Confirmed the Differences in GFAP Expression between the Peyer’s Patches and Lamina Propria in CD Patients and Non-IBD Controls
3.4. Increased Numbers of EGCS100β+ in CD Patients Compared to Non-IBD Controls
3.5. Higher Numbers of EGCS100β+ in the Peyer’s Patches of CD Patients Compared to Non-IBD Controls, and Lower Numbers of EGCS100β+ in Peyer’s Patches Compared to Lamina Propria in Non-IBD Controls
3.6. Differences in S100β Expression between Peyer’s Patches and Lamina Propria in CD Patients and Non-IBD Controls by Western Blotting
3.7. Increased Paracellular Permeability by EGC Mediators in CD Patients While Decrease in Controls
3.8. No Effect on the Results by Patient Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.Z.; van Sommeren, S.; Huang, H.; Ng, S.C.; Alberts, R.; Takahashi, A.; Ripke, S.; Lee, J.C.; Jostins, L.; Shah, T.; et al. Association Analyses Identify 38 Susceptibility Loci for Inflammatory Bowel Disease and Highlight Shared Genetic Risk across Populations. Nat. Genet. 2015, 47, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.N.; Park, C.; Park, S.J.; Lee, C.K.; Ye, B.D.; Kim, Y.S.; Lee, S.; Chae, J.; Kim, J.-I.; Kim, Y.-H.; et al. Deep Resequencing of 131 Crohn’s Disease Associated Genes in Pooled DNA Confirmed Three Reported Variants and Identified Eight Novel Variants. Gut 2016, 65, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N. Epidemiology and Risk Factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Ramos, G.P.; Papadakis, K.A. Mechanisms of Disease: Inflammatory Bowel Diseases. Mayo Clin. Proc. 2019, 94, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, Y.; Kamoi, R.; Iida, M. Pathogenesis of Aphthoid Ulcers in Crohn’s Disease: Correlative Findings by Magnifying Colonoscopy, Electron Microscopy, and Immunohistochemistry. Gut 1996, 38, 724–732. [Google Scholar] [CrossRef]
- Keita, A.V.; Söderholm, J.D. Barrier Dysfunction and Bacterial Uptake in the Follicle-Associated Epithelium of Ileal Crohn’s Disease. Ann. N. Y. Acad. Sci. 2012, 1258, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.S.A. Mucosal Integrity and Barrier Function in the Pathogenesis of Early Lesions in Crohn’s Disease. J. Clin. Pathol. 2005, 58, 568–572. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, B.; Magnusson, M.K.; Öhman, L. Mucosal Immune System of the Gastrointestinal Tract: Maintaining Balance between the Good and the Bad. Scand. J. Gastroenterol. 2017, 52, 1–9. [Google Scholar] [CrossRef]
- Casado-Bedmar, M.; Heil, S.D.S.; Myrelid, P.; Söderholm, J.D.; Keita, Å.V. Upregulation of Intestinal Mucosal Mast Cells Expressing VPAC1 in Close Proximity to Vasoactive Intestinal Polypeptide in Inflammatory Bowel Disease and Murine Colitis. Neurogastroenterol. Motil. 2019, 31, e13503. [Google Scholar] [CrossRef]
- Gullberg, E.; Keita, A.V.; Salim, S.Y.; Andersson, M.; Caldwell, K.D.; Söderholm, J.D.; Artursson, P. Identification of Cell Adhesion Molecules in the Human Follicle-Associated Epithelium That Improve Nanoparticle Uptake into the Peyer’s Patches. J. Pharmacol. Exp. Ther. 2006, 319, 632–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keita, A.V.; Gullberg, E.; Ericson, A.-C.; Salim, S.Y.; Wallon, C.; Kald, A.; Artursson, P.; Söderholm, J.D. Characterization of Antigen and Bacterial Transport in the Follicle-Associated Epithelium of Human Ileum. Lab. Investig. 2006, 86, 504–516. [Google Scholar] [CrossRef] [Green Version]
- Keita, Å.V.; Alkaissi, L.Y.; Holm, E.B.; Heil, S.D.S.; Chassaing, B.; Darfeuille-Michaud, A.; McKay, D.M.; Söderholm, J.D. Enhanced E. Coli LF82 Translocation through the Follicle-Associated Epithelium in Crohn’s Disease Is Dependent on Long Polar Fimbriae and CEACAM6 Expression, and Increases Paracellular Permeability. J. Crohns Colitis 2020, 14, 216–229. [Google Scholar] [CrossRef] [PubMed]
- You, X.-Y.; Zhang, H.-Y.; Han, X.; Wang, F.; Zhuang, P.-W.; Zhang, Y.-J. Intestinal Mucosal Barrier Is Regulated by Intestinal Tract Neuro-Immune Interplay. Front. Pharmacol. 2021, 12, 659716. [Google Scholar] [CrossRef] [PubMed]
- Vergnolle, N.; Cirillo, C. Neurons and Glia in the Enteric Nervous System and Epithelial Barrier Function. Physiology 2018, 33, 269–280. [Google Scholar] [CrossRef]
- Seguella, L.; Gulbransen, B.D. Enteric Glial Biology, Intercellular Signalling and Roles in Gastrointestinal Disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 571–587. [Google Scholar] [CrossRef]
- Yu, Y.-B.; Li, Y.-Q. Enteric Glial Cells and Their Role in the Intestinal Epithelial Barrier. World J. Gastroenterol. 2014, 20, 11273–11280. [Google Scholar] [CrossRef]
- Steinkamp, M.; Gundel, H.; Schulte, N.; Spaniol, U.; Pflueger, C.; Zizer, E.; von Boyen, G.B.T. GDNF Protects Enteric Glia from Apoptosis: Evidence for an Autocrine Loop. BMC Gastroenterol. 2012, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Meira de-Faria, F.; Casado-Bedmar, M.; Mårten Lindqvist, C.; Jones, M.P.; Walter, S.A.; Keita, Å.V. Altered Interaction between Enteric Glial Cells and Mast Cells in the Colon of Women with Irritable Bowel Syndrome. Neurogastroenterol. Motil. 2021, 33, e14130. [Google Scholar] [CrossRef]
- Schoultz, I.; Keita, Å.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef]
- Kinchen, J.; Chen, H.H.; Parikh, K.; Antanaviciute, A.; Jagielowicz, M.; Fawkner-Corbett, D.; Ashley, N.; Cubitt, L.; Mellado-Gomez, E.; Attar, M.; et al. Structural Remodeling of the Human Colonic Mesenchyme in Inflammatory Bowel Disease. Cell 2018, 175, 372–386.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.J.; Kieffer, E.J.; Powley, T.L. Loss of Glia and Neurons in the Myenteric Plexus of the Aged Fischer 344 Rat. Anat. Embryol. 2004, 209, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.J.; Powley, T.L. Innervation of the Gastrointestinal Tract: Patterns of Aging. Auton. Neurosci. 2007, 136, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogilevski, T.; Burgell, R.; Aziz, Q.; Gibson, P.R. The Role of the Autonomic Nervous System in the Pathogenesis and Therapy of IBD. Aliment. Pharmacol. Ther. 2019, 50, 720–737. [Google Scholar] [CrossRef] [PubMed]
- Villanacci, V.; Bassotti, G.; Nascimbeni, R.; Antonelli, E.; Cadei, M.; Fisogni, S.; Salerni, B.; Geboes, K. Enteric Nervous System Abnormalities in Inflammatory Bowel Diseases. Neurogastroenterol. Motil. 2008, 20, 1009–1016. [Google Scholar] [CrossRef]
- von Boyen, G.B.T.; Schulte, N.; Pflüger, C.; Spaniol, U.; Hartmann, C.; Steinkamp, M. Distribution of Enteric Glia and GDNF during Gut Inflammation. BMC Gastroenterol. 2011, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Cornet, A.; Savidge, T.C.; Cabarrocas, J.; Deng, W.L.; Colombel, J.F.; Lassmann, H.; Desreumaux, P.; Liblau, R.S. Enterocolitis Induced by Autoimmune Targeting of Enteric Glial Cells: A Possible Mechanism in Crohn’s Disease? Proc. Natl. Acad. Sci. USA 2001, 98, 13306–13311. [Google Scholar] [CrossRef] [Green Version]
- Steinkamp, M.; Schulte, N.; Spaniol, U.; Pflüger, C.; Hartmann, C.; Kirsch, J.; von Boyen, G.B. Brain Derived Neurotrophic Factor Inhibits Apoptosis in Enteric Glia during Gut Inflammation. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2012, 18, BR117–BR122. [Google Scholar] [CrossRef]
- Boesmans, W.; Lasrado, R.; Berghe, P.V.; Pachnis, V. Heterogeneity and Phenotypic Plasticity of Glial Cells in the Mammalian Enteric Nervous System. Glia 2015, 63, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Pochard, C.; Coquenlorge, S.; Freyssinet, M.; Naveilhan, P.; Bourreille, A.; Neunlist, M.; Rolli-Derkinderen, M. The Multiple Faces of Inflammatory Enteric Glial Cells: Is Crohn’s Disease a Gliopathy? Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G1–G11. [Google Scholar] [CrossRef] [Green Version]
- Bertheloot, D.; Latz, E. HMGB1, IL-1α, IL-33 and S100 Proteins: Dual-Function Alarmins. Cell. Mol. Immunol. 2017, 14, 43–64. [Google Scholar] [CrossRef] [Green Version]
- Xia, C.; Braunstein, Z.; Toomey, A.C.; Zhong, J.; Rao, X. S100 Proteins as an Important Regulator of Macrophage Inflammation. Front. Immunol. 2018, 8, 1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drokhlyansky, E.; Smillie, C.S.; Van Wittenberghe, N.; Ericsson, M.; Griffin, G.K.; Eraslan, G.; Dionne, D.; Cuoco, M.S.; Goder-Reiser, M.N.; Sharova, T.; et al. The Human and Mouse Enteric Nervous System at Single-Cell Resolution. Cell 2020, 182, 1606–1622.e23. [Google Scholar] [CrossRef]
- Meir, M.; Flemming, S.; Burkard, N.; Bergauer, L.; Metzger, M.; Germer, C.-T.; Schlegel, N. Glial Cell Line-Derived Neurotrophic Factor Promotes Barrier Maturation and Wound Healing in Intestinal Epithelial Cells in Vitro. Am. J. Physiol.-Gastrointest. Liver Physiol. 2015, 309, G613–G624. [Google Scholar] [CrossRef] [Green Version]
- Meir, M.; Burkard, N.; Ungewiß, H.; Diefenbacher, M.; Flemming, S.; Kannapin, F.; Germer, C.-T.; Schweinlin, M.; Metzger, M.; Waschke, J.; et al. Neurotrophic Factor GDNF Regulates Intestinal Barrier Function in Inflammatory Bowel Disease. J. Clin. Investig. 2019, 129, 2824–2840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Landeghem, L.; Mahé, M.M.; Teusan, R.; Léger, J.; Guisle, I.; Houlgatte, R.; Neunlist, M. Regulation of Intestinal Epithelial Cells Transcriptome by Enteric Glial Cells: Impact on Intestinal Epithelial Barrier Functions. BMC Genom. 2009, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Hagbom, M.; De Faria, F.M.; Winberg, M.E.; Westerberg, S.; Nordgren, J.; Sharma, S.; Keita, Å.V.; Loitto, V.; Magnusson, K.-E.; Svensson, L. Neurotrophic Factors Protect the Intestinal Barrier from Rotavirus Insult in Mice. mBio 2020, 11, e02834-19. [Google Scholar] [CrossRef] [Green Version]
- Xie, Q.; Chen, X.; Meng, Z.M.; Huang, X.L.; Zhang, Q.; Zhou, J.Q.; Zhang, L.; He, F.Q.; Zou, Y.P.; Gan, H.T. Glial-Derived Neurotrophic Factor Regulates Enteric Mast Cells and Ameliorates Dextran Sulfate Sodium-Induced Experimental Colitis. Int. Immunopharmacol. 2020, 85, 106638. [Google Scholar] [CrossRef]
- Keita, A.V.; Carlsson, A.H.; Cigéhn, M.; Ericson, A.-C.; McKay, D.M.; Söderholm, J.D. Vasoactive Intestinal Polypeptide Regulates Barrier Function via Mast Cells in Human Intestinal Follicle-Associated Epithelium and during Stress in Rats. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2013, 25, e406–e417. [Google Scholar] [CrossRef] [PubMed]
- Keita, A.V.; Salim, S.Y.; Jiang, T.; Yang, P.-C.; Franzén, L.; Söderkvist, P.; Magnusson, K.-E.; Söderholm, J.D. Increased Uptake of Non-Pathogenic E. Coli via the Follicle-Associated Epithelium in Longstanding Ileal Crohn’s Disease. J. Pathol. 2008, 215, 135–144. [Google Scholar] [CrossRef]
- Arranz, A.; Juarranz, Y.; Leceta, J.; Gomariz, R.P.; Martínez, C. VIP Balances Innate and Adaptive Immune Responses Induced by Specific Stimulation of TLR2 and TLR4. Peptides 2008, 29, 948–956. [Google Scholar] [CrossRef]
- Schratzberger, P.; Geiseler, A.; Dunzendorfer, S.; Reinisch, N.; Kähler, C.M.; Wiedermann, C.J. Similar Involvement of VIP Receptor Type I and Type II in Lymphocyte Chemotaxis. J. Neuroimmunol. 1998, 87, 73–81. [Google Scholar] [CrossRef]
- Yadav, M.; Rosenbaum, J.; Goetzl, E.J. Cutting Edge: Vasoactive Intestinal Peptide (VIP) Induces Differentiation of Th17 Cells with a Distinctive Cytokine Profile. J. Immunol. 2008, 180, 2772–2776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toumi, F.; Neunlist, M.; Denis, M.G.; Oreshkova, T.; Laboisse, C.L.; Galmiche, J.-P.; Jarry, A. Vasoactive Intestinal Peptide Induces IL-8 Production in Human Colonic Epithelial Cells via MAP Kinase-Dependent and PKA-Independent Pathways. Biochem. Biophys. Res. Commun. 2004, 317, 187–191. [Google Scholar] [CrossRef]
- Murakami, M.; Ohta, T.; Ito, S. Lipopolysaccharides Enhance the Action of Bradykinin in Enteric Neurons via Secretion of Interleukin-1beta from Enteric Glial Cells. J. Neurosci. Res. 2009, 87, 2095–2104. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Ye, D.; Said, H.M.; Ma, T.Y. IL-1β-Induced Increase in Intestinal Epithelial Tight Junction Permeability Is Mediated by MEKK-1 Activation of Canonical NF-ΚB Pathway. Am. J. Pathol. 2010, 177, 2310–2322. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (y) | Sex | Anti-Inflammatory Medication | Indication for Surgery | Primary or Recurrent Surgery | Montreal Classification | Pre-op p-CRP |
|---|---|---|---|---|---|---|
| 61 | M | None | Stricture | Primary | A2L3B2 | 13 |
| 63 | M | None | Stricture, abscess | Recurrent | A2L3B3 | <10 |
| 20 | M | Azathioprine | Stricture, fistula | Primary | A2L1B3 | <10 |
| 38 | F | None | Stricture | Primary | A2L1B2 | 10 |
| 38 | F | None | Stricture | Primary | A2L3B2p | <10 |
| 49 | F | None | Stricture, abscess | Primary | A3L1B2 | <10 |
| 50 | M | Azathioprine, infliximab | Stricture | Primary | A2L1B2 | <10 |
| 25 | F | Azathioprine | Stricture | Recurrent | A1L1B3 | <10 |
| 49 | M | None | Stricture, abscess | Recurrent | A2L1B3 | 26 |
| 46 | M | None | Stricture | Primary | A2L1B2 | <10 |
| 43 | F | None | Stricture | Primary | A2L3B2 | 42 |
| 49 | M | None | Stricture | Recurrent | A2L1B3 | <10 |
| 49 | F | Ustekinumab | Stricture | Recurrent | A2L3B2 | <10 |
| 55 | M | Azathioprine, infliximab | Stricture | Recurrent | A2L1B2 | <10 |
| 29 | F | None | Fistulas | Recurrent | A1L1B3 | <10 |
| 29 | M | Thiopurine, adalimumab | Fistula | Recurrent | A2L3B3p | <10 |
| 43 | M | Mesalazine, infliximab | Fistula | Primary | A2L3B3 | <10 |
| 27 | M | None | Stricture, fistula, abscess | Recurrent | A1L3B3 | 19 |
| 21 | M | None | Stricture | Recurrent | A2L1B3 | <10 |
| 17 | F | Budesonide, azathioprine, infliximab | Stricture | Primary | A1L3B2 | <10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biskou, O.; Meira de-Faria, F.; Walter, S.M.; Winberg, M.E.; Haapaniemi, S.; Myrelid, P.; Söderholm, J.D.; Keita, Å.V. Increased Numbers of Enteric Glial Cells in the Peyer’s Patches and Enhanced Intestinal Permeability by Glial Cell Mediators in Patients with Ileal Crohn’s Disease. Cells 2022, 11, 335. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030335
Biskou O, Meira de-Faria F, Walter SM, Winberg ME, Haapaniemi S, Myrelid P, Söderholm JD, Keita ÅV. Increased Numbers of Enteric Glial Cells in the Peyer’s Patches and Enhanced Intestinal Permeability by Glial Cell Mediators in Patients with Ileal Crohn’s Disease. Cells. 2022; 11(3):335. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030335
Chicago/Turabian StyleBiskou, Olga, Felipe Meira de-Faria, Susanna M. Walter, Martin E. Winberg, Staffan Haapaniemi, Pär Myrelid, Johan D. Söderholm, and Åsa V. Keita. 2022. "Increased Numbers of Enteric Glial Cells in the Peyer’s Patches and Enhanced Intestinal Permeability by Glial Cell Mediators in Patients with Ileal Crohn’s Disease" Cells 11, no. 3: 335. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030335