
Regulation of Nucleolar Activity by MYC



1
Department of Cell Biology, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA
2
Hamon Center for Regenerative Science and Medicine, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA
3
Harold C. Simmons Comprehensive Cancer Center, University of Texas Southwestern Medical Center, Dallas, TX 75390, USA
*
Authors to whom correspondence should be addressed.
Cells 2022, 11(3), 574; https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030574
Submission received: 7 January 2022
/
Revised: 2 February 2022
/
Accepted: 3 February 2022
/
Published: 7 February 2022
(This article belongs to the Special Issue Nucleolar Organization and Functions in Health and Disease II)
Abstract
:The nucleolus harbors the machinery necessary to produce new ribosomes which are critical for protein synthesis. Nucleolar size, shape, and density are highly dynamic and can be adjusted to accommodate ribosome biogenesis according to the needs for protein synthesis. In cancer, cells undergo continuous proliferation; therefore, nucleolar activity is elevated due to their high demand for protein synthesis. The transcription factor and universal oncogene MYC promotes nucleolar activity by enhancing the transcription of ribosomal DNA (rDNA) and ribosomal proteins. This review summarizes the importance of nucleolar activity in mammalian cells, MYC’s role in nucleolar regulation in cancer, and discusses how a better understanding (and the potential inhibition) of aberrant nucleolar activity in cancer cells could lead to novel therapeutics.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Nucleolar Structure and Organization Allow for Flexibility in the Rate of Ribosome Biogenesis
The nucleolus, one of the largest membraneless organelles, is located in the nucleus and is the home of all of the steps necessary for ribosomal biogenesis including rDNA transcription, ribosomal RNA (rRNA) processing, and the assembly of ribosomes. The lack of a membrane allows the nucleolus to be very dynamic, as its size and composition can adjust to the demands for protein synthesis. For example, in cases of hyperproliferation, a higher rate of protein synthesis and ribosome biogenesis must be achieved to allow for cell growth, and the nucleolus increases in size and density to accommodate those needs [1,2].
Nucleolar morphology and function are tightly regulated during the cell cycle. Nucleoli are most active in G2, the cell cycle phase that precedes cell division and when protein synthesis capacity is the highest [3]. Conversely, during prophase, when cells are dividing, the nucleoli are disassembled and their contents are dispersed to eventually be inherited by daughter cells produced through mitosis [4]. Nucleoli are then re-formed in the daughter cells through the activity of nucleolar organizer regions (NORs), which are 50 Kb- to 6 Mb-long regions containing rDNA copies. In humans, NORs are located on chromosomes 13, 14, 15, 21, and 22 [5]. Strikingly, there are around 200 rDNA genes per human haploid genome, making rRNA the most abundant RNA in a cell [6,7].
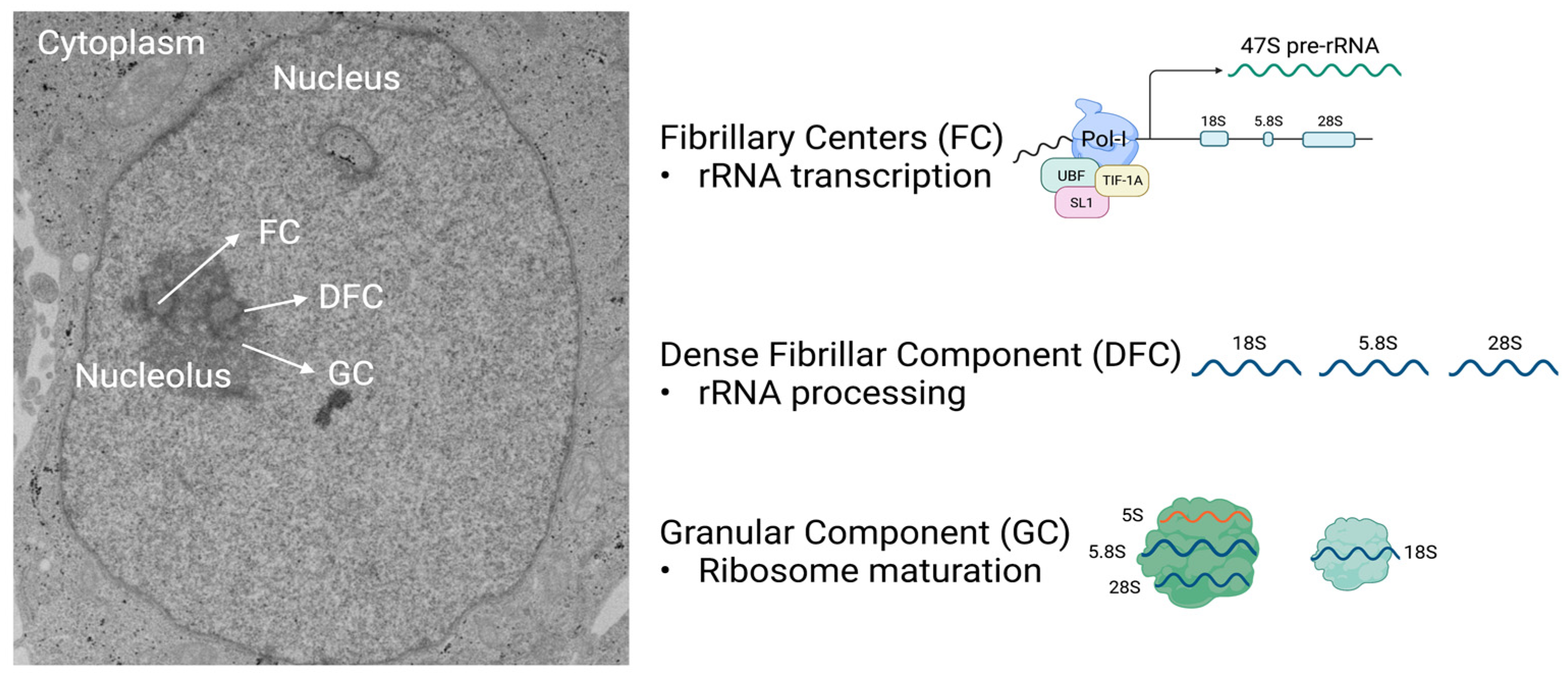
Mammalian cells may contain one or more tripartite nucleoli, and each comprise three morphologically and biochemically distinct compartments that perform specific roles to produce ribosomal subunits: the fibrillary center (FC), the dense fibrillar component (DFC), and the granular component (GC). The FCs are encased in DFCs, which are surrounded by the GC (Figure 1). These three compartments work together to perform every step necessary to generate large (60S) and small (40S) ribosome subunits that comprise rRNA and ribosomal proteins (RPs) (Figure 1) [8]. Other organisms including yeast, invertebrates, fish, and reptiles have bipartite nucleoli, where the FCs are not present and rDNA genes are dispersed within the DFC [9,10,11,12,13]. The GC, which is the outermost layer of the nucleolus, is present in all species and is the site for the final steps in ribosome biogenesis. Through interactions with the nuclear pore complex, near-mature ribosome subunits are exported to the cytoplasm, where ribosome maturation is completed, thus allowing for the formation of complete ribosomes (80S) in the presence of mRNA [14,15,16]. Final ribosome maturation occurring in the cytoplasm prevents premature translation initiation until the ribosomes reach the cytoplasm.
The initial step of ribosome biogenesis begins in the FC when upstream binding factor (UBF) and selectivity factor 1 (SL1), also known as TIF-1B, and TIF-1A, bind to RNA polymerase I (RNAPolI) at the rDNA promoter. This initiates the transcription of the 47S pre-rRNA precursor, giving rise to 28S, 18S, and 5.8S mature rRNAs [17,18,19]. The synthesis of this 47S pre-rRNA as a single transcript allows for the stoichiometric production of the small and large ribosomal subunits, required at 1:1 ratio for translation. The 5S rRNA is the only rRNA not transcribed as part of 47S pre-rRNA; it is transcribed by the RNA polymerase III (RNAPolIII) in the nucleus [20,21].
The rRNAs undergo several steps of processing including methylations and pseudouridylations [22,23]. These modifications, which are guided by small nuclear RNAs (snoRNAs), are essential for the stabilization of secondary and tertiary rRNA structures, increasing translation accuracy [24,25,26]. Most steps in pre-rRNA maturation occur in the DFC where rRNA undergoes processing events that result in the mature rRNAs 28S and 5.8S, components of the large ribosome subunit, and 18S, a component of the small ribosome subunit [27]. The addition of RPs to rRNA begin co-transcriptionally. These RPs may be important in delimiting where the rRNA is cleaved. Other RPs are attached to the 28S and 18S in the GC of the nucleolus. All RP are transcribed from nuclear genes via the activity of the RNA polymerase II (RNAPolII). Once the RPs are translated in the cytoplasm, they are imported into the nucleus, where a subset of them bind to 5S rRNA. These are then co-imported into the GC of the nucleolus to be assembled into ribosomes. The mature 40S subunit comprises 32 small RPs, and the 60S subunit comprises 47 large RPs.
Errors in rDNA transcription, rRNA processing, and ribosome assembly are the cause of several disorders collectively named ribosomopathies. These include degenerative diseases such as abdominal aortic aneurysm [28,29] and Parkinson’s disease [30], along with developmental disorders such as Treacher Collins syndrome [31,32], Diamond-Blackfan anemia [33,34,35], Bowen-Conradi syndrome [36], and autosomal recessive primary microcephaly [37].
2. Nucleolar Size and Activity Are Increased in Hyperproliferative Cells
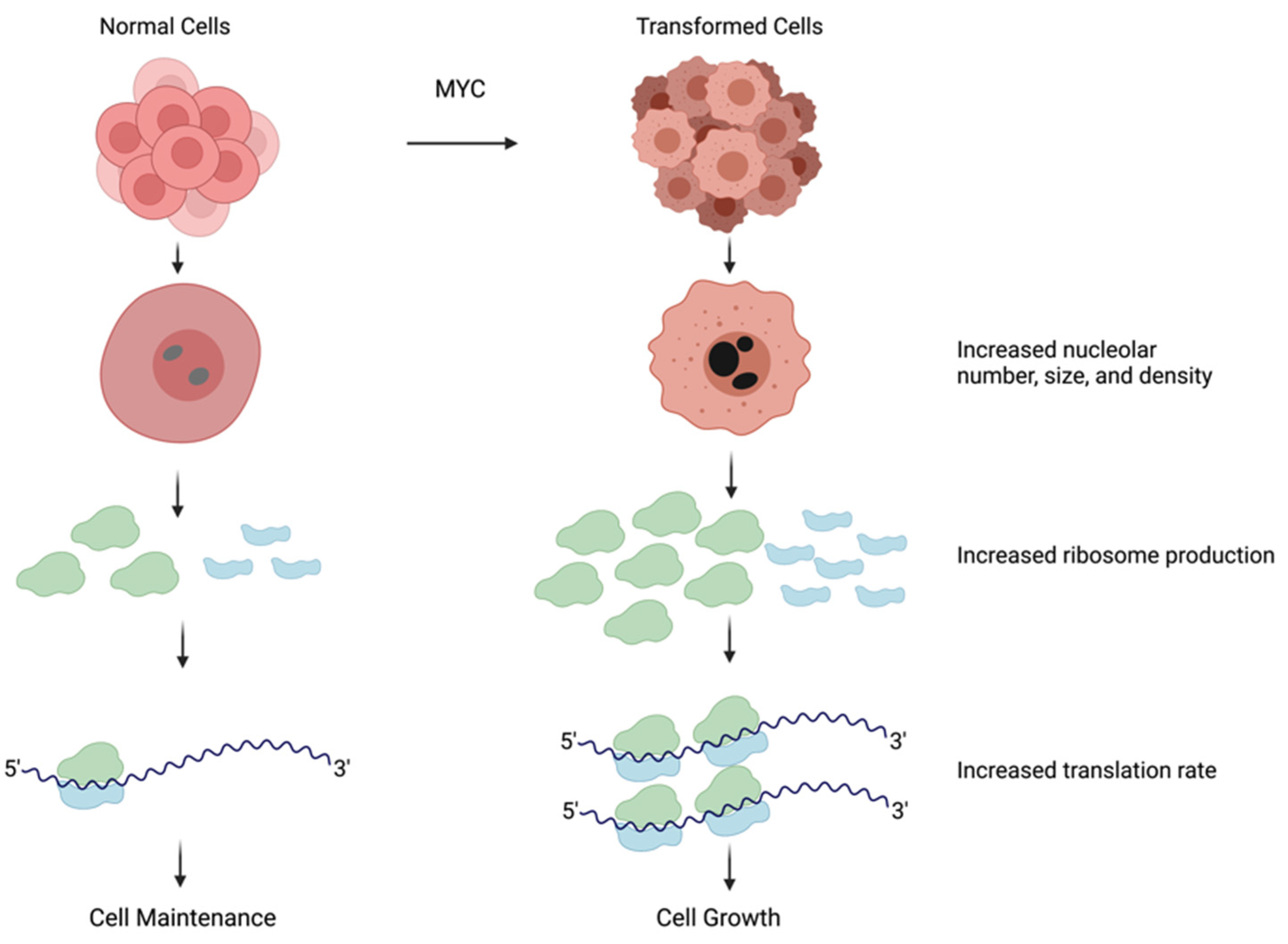
The presence of nutrients and growth factors in the cellular environment leads to the activation of signaling pathways that promote cell growth, including the production of new ribosomes. Hyperproliferative cancer cells often display activated growth factor signaling that can induce elevated RNAPolI–mediated rDNA transcription [38,39,40], leading to an increase in ribosome production. Moreover, alterations in the number of rDNA or 5S rRNA loci, documented in cancer cells, have the potential to increase ribosome production [41,42]. Elevated ribosome biogenesis causes morphological changes in the nucleoli that can be used for cancer diagnostics. Cancer cells have dark, dense, and large nucleoli, and these features are used by pathologists when grading solid tumors [2,14,43].
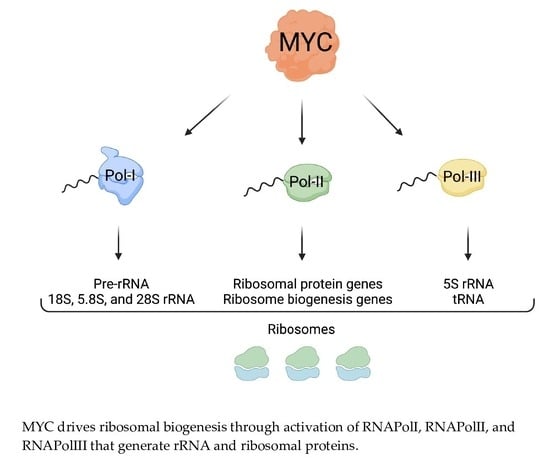
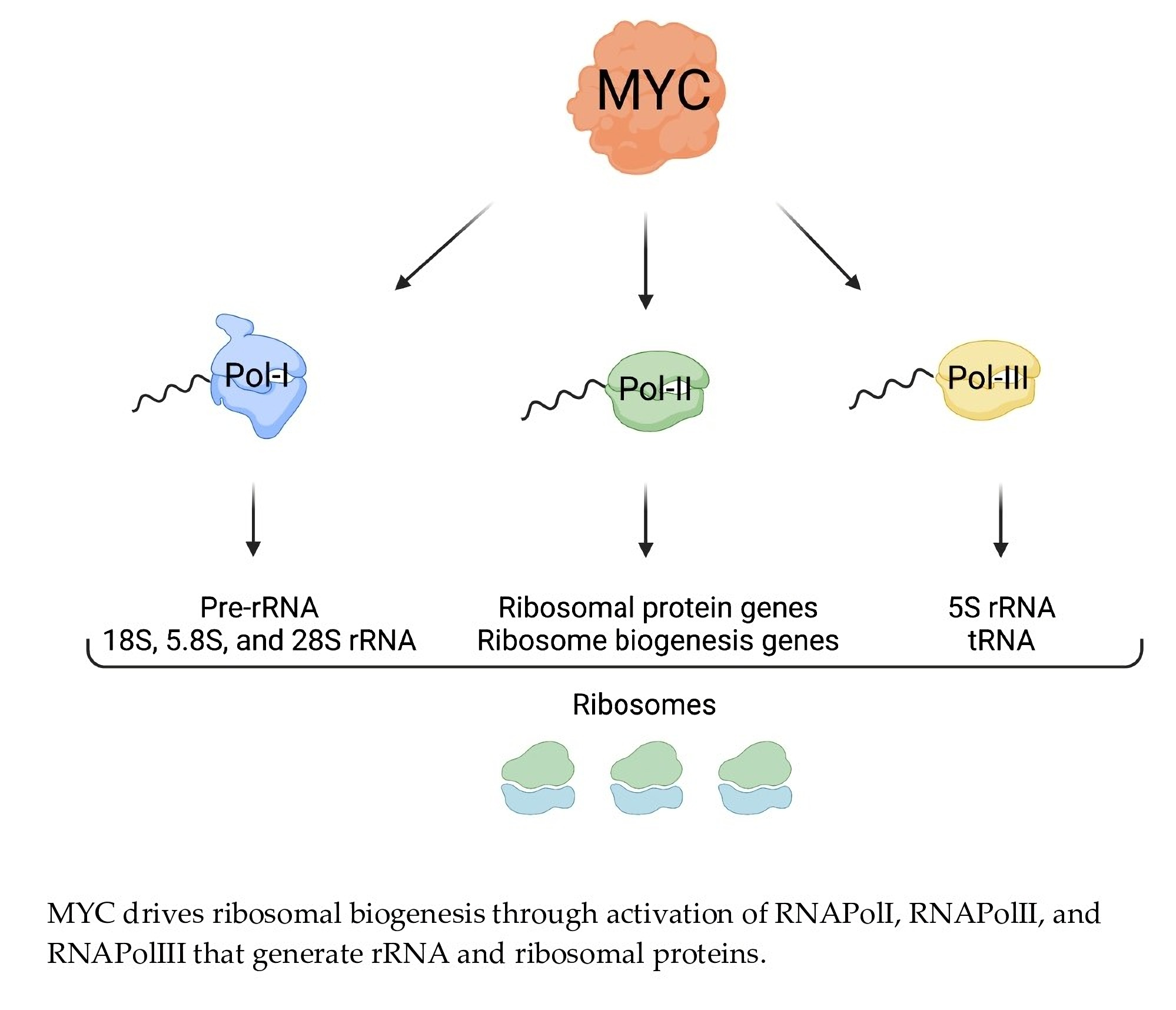
Due to the high demand for ribosomes, transformed cells hyperactivate nearly all steps in ribosome biogenesis including rDNA transcription, rRNA processing, and the expression of RPs [1]. These changes are correlated with the activation of oncogenes and the inactivation of tumor suppressors. For example, the loss of tumor suppressors, such as p53 or PTEN, may lead to an increase in rDNA transcription, thus promoting ribosome biogenesis [44]. The oncogene MYC is one of the most potent drivers of ribosome biogenesis owing to its ability to concurrently promote the transcription of rDNA as well as genes encoding for ribosomal components and key regulators of ribosome biogenesis (Figure 2) [40,45,46,47].
3. The MYC Family of Transcription Factors Are Key Regulators of Cell Growth
MYC was originally discovered as a viral oncogene (v-myc) that caused myelocytomatosis, leukemia, and sarcoma [48]. Interestingly, v-myc was acquired from a cellular oncogene dubbed c-myc, now referred as MYC [49,50,51,52]. Subsequently, increasing evidence indicated that, in humans, MYC played a role in tumorigenesis without involving a viral infection [53]. Since then, there has been a tremendous effort to better understand MYC’s role under both normal and cancerous conditions. The discovery that cancer was not a contagious disease, but that viruses had the ability to capture and carry mammalian genes was groundbreaking. The attention then turned to developing an understanding of the mechanisms by which MYC promotes deregulated growth and cell transformation.
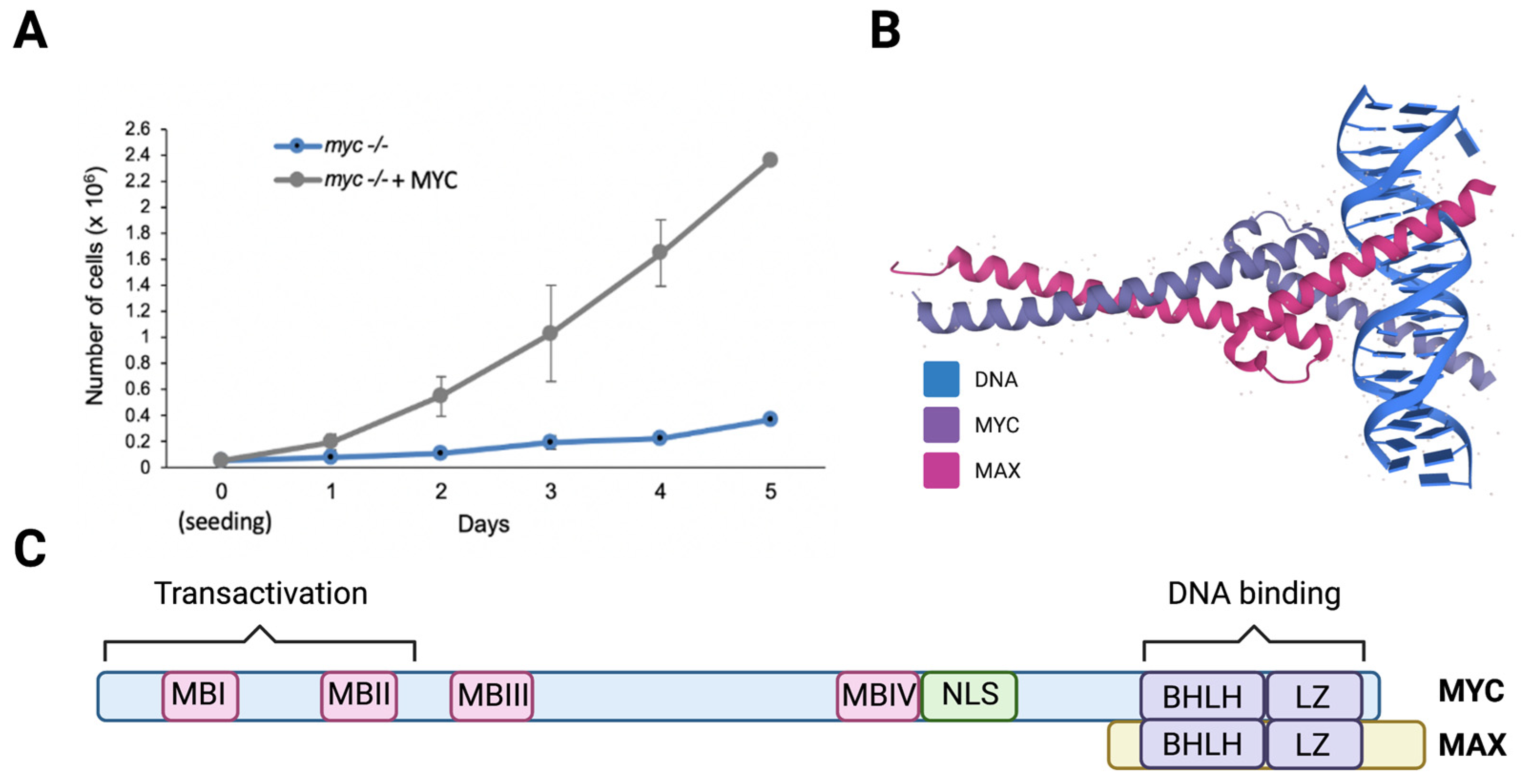
MYC was found to be elevated in 70% of human cancers, making it one of the most pervasive oncogenes [54]. Supporting the correlation between MYC and cell growth, knocking out myc in cultured cells, including fibroblasts, causes a dramatic reduction in proliferation, and the reconstitution of MYC by ectopic expression rescues this phenotype (Figure 3A). Cells expressing elevated MYC levels are highly proliferative, and exhibit enlarged and active nucleoli [55].
The MYC family of proteins includes three transcription factors (MYC, MYC-N, and MYC-L) that regulate key biological processes; specifically, MYC and MYCN are essential for embryonic development [56,57]. Members of the MYC family contain a basic helix-loop-helix and leucine zipper (bHLHZip) domain in their C-terminal region that creates an alpha-helix structure through which they heterodimerize with MAX, a small bHLHZ protein [58,59,60,61] (Figure 3B). MYC-MAX heterodimers bind to and directly stimulate the transcription of genes that contain E-boxes (5′- CACGTG-3′) in their promoters [62,63]. There is evidence that MYC binds to non-E-box DNA sequences, with or without heterodimerizing MAX, via interacting with other DNA binding proteins such as RNAPolIII subunit TFIIIB [64,65,66]. MYC-NICK, a cytoplasmic proteolytic product of MYC that lacks the DNA binding domain and thus, is unable to regulate gene transcription, was also shown to play a pro-survival role in cancer cells [67,68,69].
All MYC variants contain highly conserved regions called MYC boxes (MB) (Figure 3C) that are important for transcription and protein stability [70]. MYC proteins have half-lives of around 30 min in normal cells and are considered unstable [71]. MBI contains a MYC degron, harboring the amino acids T58 and S62, which are phosphorylated preceding its targeting for ubiquitin-mediated degradation by its binding to FBWX7, a substrate adaptor protein for the SCF E3-ligase complex [72]. Mutations in MYC’s degron, or its E3 ligases, inhibits MYC degradation, causing its stabilization, which then contributes to cell transformation [73,74]. Moreover, the mutational inactivation of FBWX7 in tumors [75,76,77] causes the stabilization of MYC and other oncogenes such as Notch, cyclin E, and c-Jun that cooperate to drive cellular transformation [78,79,80,81,82,83].
MBII is a crucial region where histone acetyltransferase (HAT) complexes bind. HATs promote histone acetylation, which opens chromatin, facilitating access by the transcription machinery and ultimately activating gene transcription [84,85,86]. MYC increases local acetylation by binding to acetyltransferase complexes that acetylate histones H3 and H4 [87]. MYC specifically interacts with transcription-domain-associated-protein (TRRAP) [88], general control of amino acid synthesis protein 5 (GCN5) [66], Tat Interacting Protein, 60kDa (TIP60, also known as KAT5), and CREB-binding protein (p300/CBP) acetyltransferases [89]. MBIII and MBIV are less studied but are proposed to be important for the regulation of apoptosis and transcription by MYC [90,91].
4. MYC Promotes Transcription of rDNA
For proliferating cells to meet the ribosome and protein synthesis demand, rRNA production must be maintained at a high rate (Figure 2). The regulation of rDNA transcription by MYC has been documented in Drosophila, as well as in vertebrates [15]. MYC aids in rDNA transcription in two ways. First, MYC binds to rDNA loci together with RNAPolI, which facilitates transcriptional activation [46]. Second, MYC also enhances rDNA transcription by binding at the promoters of the RNAPolI cofactors SL1, UBF, and TIF-1A [39,56], which are often elevated in cancers, likely resulting from MYC transcriptional activity. UBF activates RNAPolI by stimulating transcriptional elongation [92] and SL1 stabilizes UBF and facilitates the pre-initiation complex formation on the rDNA promoter [93]. Therefore, elevated MYC in tumors can promote the activation of RNAPolI to transcribe rDNA into pre-rRNA (47S). Moreover, MYC facilitates the transcription of RNAPolIII–mediated 5S rRNA and transfer RNAs (tRNA), which carry amino acids to the translation machinery [65]. Additionally, MYC directly promotes the transcription of TFIIIB, critical for the activity of RNAPolIII [56]. While MYC induces transcription through RNA PolI, PolII, and PolIII activation, it also plays a role in stimulating rDNA transcription via chromatin remodeling mechanisms [94,95,96,97,98]. Through these multiple mechanisms, MYC can drive a dramatic increase in rRNA production, contributing to ribosome biogenesis in cancer cells.
5. MYC Promotes the Transcription of Genes Encoding RPs and Regulators of Nucleolar Assembly and Activity
MYC controls the expression of about 20% of the genome, including genes that are crucial for nucleolar activity [15,55,99,100,101]. By comparing the transcriptional signature of myc-/- fibroblasts expressing empty vector or reconstituted with MYC, we found that MYC promotes the expression of the regulators of nucleolar activity (Figure 4A,B), as well as the structural components of the ribosome (Figure 4C,D). The expression of 38% of the genes encoding for small RPs and 61% of the genes encoding for large RPs are increased in MYC-expressing cells. It is likely that by increasing the expression of structural components of the ribosome, MYC supports an increase in ribosome subunit assembly and in protein synthesis. Moreover, MYC regulates the expression of numerous regulators of nucleolar assembly and activity [102]. For example, MYC promotes the transcription of nucleolin (NCL), which is needed for the processing of 47S pre-rRNA [103]. Furthermore, MYC directly regulates the expression of nucleophosmin (NPM1) and fibrillarin (FBL). Due to their biophysical properties, NPM1 and FBL separate into immiscible phases, contributing to the assembly of the GC and DFC, respectively (Figure 4B) [104,105]. The expression of these proteins must scale up to increase nucleolar size and function in cancer cells [2,43]. FBL and NPM1 were shown to be necessary for rRNA processing and the transport of 40S and 60S subunits into the cytoplasm [106,107]. Whether these functions of FBL and NPM1 are independent, or a result of their properties in assembling the nucleolar phases remains to be determined. This suggests that MYC, in addition to promoting the production of rRNA and RPs (Figure 4), may participate in regulating nucleolar assembly via the induction in FBL and NPM1.
6. MYC Promotes the Expression of Genes That Activate Ribosome Biogenesis and Protein Synthesis
In addition to directly regulating the transcription of rDNA and RPs, MYC promotes the transcription of additional transcription factors that regulate one or more aspects of nucleolar function. For example, MYC binds to the promoter of the transcription factor aryl hydrocarbon receptor (AHR) and of its heterodimeric partner aryl hydrocarbon receptor nuclear translocator (ARNT) and promotes their expression [55]. AHR in turn regulates the expression of genes involved in rDNA transcription such as nucleolar and coiled-body phosphoprotein 1 (NOLC1), rRNA processing such as periodic tryptophan protein 2 homolog (PWP2), and protein synthesis such as 2-oxoglutarate and iron dependent oxygenase domain containing 1 (OGFOD1) in MYC-expressing cells [55]. Consequently, knocking down AHR in MYC-expressing cells causes decreased proliferation, nucleolar disassembly, and reduced protein synthesis. This suggests that MYC regulates nucleolar activity, and thus protein synthesis, partly through the transcriptional regulation of AHR. Moreover, AHR also regulates the production of nucleotides that are necessary for the elevated rate of rRNA production in cancer cells [108,109]. Carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase (CAD), dihydroorotate dehydrogenase (DHODH), and uridine monophosphate synthetase (UMPS), encoding for the enzymes of the de novo pyrimidine biosynthesis pathway [110], were previously found to be induced in MYC-expressing cells [101,111,112]. Recently, AHR was found to bind to the promoters of DHODH and UMPS and to cooperate with MYC to transcribe these genes. Knocking down AHR in MYC-expressing cells decreases the expression of DHODH and UMPS and the levels of the pyrimidine nucleotide uridine 5-monophosphate (UMP), which affects the number of pyrimidines needed for rRNA synthesis [101,108]. Therefore, AHR [55] activates the expression of de novo pyrimidine biosynthesis to partially support nucleolar activity. MYC was also found to regulate the transcription of genes encoding for factors responsible for translation elongation and initiation factors such as eif4e [102,113,114]. These factors are essential in translation and help carry out this process by binding to the cap structure at the 5′ end of mRNA, initiating the first step in translation [115].
7. Nucleolar Assembly and Function Regulate MYC Levels and Activity
As discussed above, MYC was shown to regulate multiple aspects of nucleolar function, including transcription of rDNA and RPs genes (Figure 5). Conversely, nucleolar components are also shown to affect MYC levels and activity, thus creating a tight positive feedback loop: MYC promotes nucleolar activity, and nucleolar components regulate MYC levels and function. For example, MYC induces the expression of NPM1 [116] and NPM1 in turn regulates MYC activity and stability through directly interacting with MYC, which stimulates its binding to target gene promoters such as eif4e, ncl, and cdk4 [107,116,117]. Furthermore, NPM1 was proposed to be necessary for the ability of MYC to induce rRNA synthesis in the nucleolus. Thus, constitutive NPM1 overexpression stimulates MYC-mediated rRNA synthesis [117]. Conversely, other studies have shown that the elevated expression of NPM1 enhances the nucleolar localization of MYC, and that this is necessary for the FBXW7-mediated degradation of MYC in the nucleolus [72,117]. FBXW7 was reported to be localized in the nucleolus where it colocalizes with the nucleolar pool of MYC. The loss of FBXW7 in cancer cells leads to MYC stabilization which likely enhances rRNA production [72].
Additionally, a feedback loop between MYC and ribosomal proteins such as RPL5 and RPL11 was also documented. MYC activity was found to be altered by RPL11, which is capable of binding to MYC, thereby inhibiting its transcriptional activity [118]. Moreover, RPL5 was shown to facilitate the degradation of MYC mRNA by linking its 3′UTR to the RISC RNA degradation complex [119]. Further studies are needed to generate a more comprehensive view of how nucleolar resident proteins and ribosomal factors control the activity of MYC and of other growth-promoting factors in normal and cancer cells.
8. Targeting Aberrant Nucleolar Activity to Inhibit Cancer Growth
As a result of the extensive effects of MYC on ribosome biogenesis, MYC-expressing cells generally have larger and hyperactive nucleoli [43]. Therefore, MYC-dependent tumors are likely sensitive to the inhibition of nucleolar activity, making the inhibition of excessive ribosome biogenesis in tumors an attractive approach for cancer therapeutics. Nevertheless, inhibiting nucleolar activity could lead to undesirable effects given the requirement for ribosomes in normal cells. Hence, identifying and inhibiting tumor-specific nucleolar regulators (possibly driven by MYC) may lead to the development of novel strategies to block cancer cell growth.
Interestingly, some drugs, including cisplatin, oxaliplatin, and doxorubicin, were shown to have off-target effects that inhibit RNAPolI activity, altering rRNA production [120,121,122,123]. The topoisomerase II inhibitor ellipticine was shown to impair SL1 promoter binding, halting RNAPolI–mediated transcription, thus causing dramatic effects on ribosome biogenesis in vitro and in pre-clinical models [56]. Nevertheless, phase I and II clinical trials revealed unacceptable cytotoxicity; therefore, this is not a safe therapeutic option [123,124]. Currently, there is a focus on identifying drugs that target the nucleolus of cancer cells in a more specific manner, initiating a promising option for the treatment of human tumors [123,125,126,127,128,129].
Interfering with rRNA and ribosome production can alter the homeostasis and integrity of the nucleolus, activating nucleolar stress that causes cell cycle arrest and apoptosis. The best-known mechanism of nucleolar stress is mediated by p53 [130]. Under normal cellular conditions, the E3 ubiquitin ligase mouse double minute 2 (MDM2) interacts with p53, targeting it for degradation [130]. In contrast, upon nucleolar stress, RPL5, RPL11, and the 5S rRNAs redistribute from the nucleolus to the nucleoplasm where they sequester MDM2. This prevents MDM2-p53 interaction, leading to p53 stabilization, which in turn causes cell cycle arrest and apoptosis [131,132]. In line with this idea, an emerging approach to cause nucleolar stress in cancer cells is to block de novo nucleotide biosynthesis, which limits nucleotide availability and thus impairs rRNA production affecting nucleolar activity. For instance, work with small-cell lung cancer (SCLC) models showed that SCLC with elevated MYC expression were more sensitive to the inhibition of the de novo purine biosynthesis enzyme inosine-5′-monophosphate dehydrogenase (IMPDH1/2), which affected rRNA production and thus, cell proliferation [133,134]. Inhibition of the de novo pyrimidine biosynthesis enzyme DHODH has been shown to decrease the production of rRNA, to induce nucleolar disassembly, and to stabilize p53 in glioblastoma, breast, and colon cancer cells, leading to a decrease in proliferation and an increase in apoptosis [108,109]. Additional pharmacologic inhibitors may be exploited to limit nucleolar activity in cancer cells. For example, MLN4924, a chemical that inhibits neddylation, a post-translational modification that regulates the activity of cullins (scaffold components for RING E3-ubiquitin ligases), has been shown to increase p53 levels by altering the MDM2-RPL11 pathway [135]. During phase 1 clinical trials, it was found that this drug held promise as a useful cancer therapeutic, although further clinical trial phases need to be conducted [135,136]. Whether MLN4924 causes nucleolar stress in patients is yet to be determined. Altogether, targeting nucleolar activity in tumors with the aim of inducing nucleolar stress may be a potent strategy to inhibit the growth of tumors that depend on MYC, and potentially, other oncogenes as well.
9. Future Directions
In recent years, the concept of ribosome heterogeneity has emerged [137], and understanding its potential role in cancer could lead to new therapeutic strategies. When ribosomes were first discovered, it was suggested that there was a universal ribosome for every protein formed [138], and the concept that ribosomes have no specificity or regulatory functions was widely accepted [137,139]. However, recent groundbreaking work has shown that ribosome heterogeneity might play an important regulatory role in cells after all [140,141,142]. There is supportive evidence that ribosomes are not always composed of the same RPs; however, the details are not fully mapped [137,143,144,145]. Additionally, several RPs have known variants and pseudogenes that may perform specific functions [146]. Some of these RP variants and pseudogenes are expressed in specific tissues, which could potentially contribute to ribosome heterogeneity [145,146]. With this concept in mind, it has also been noted that some RPs have extra-ribosomal functions including DNA repair, development regulation, cell growth and apoptosis regulation, tumor suppressor gene and proto-oncogene regulation, and RNA splicing and modification [147,148]. Many tumor types have been characterized with an increase in RPs and specific RPs mutations [149,150]. In about 10% of T-cell acute lymphoblastic leukemia cases, patients have a frameshift mutation in RPL22, which is thought to contribute to cancer progression [151]. In colorectal cancer, the elevation of some specific small RPs, including RPS3, RPS6, RPS8, and RPS12, increases ribosome biogenesis and possibly leads to the activation of extra-ribosomal functions such as DNA replication, RNA splicing and modification, and cell growth [148,152].
The concept that ribosomes can be specialized may lead to a new research avenue for the development of drugs that target cancer-specific ribosomes. Whether MYC or other oncogenes regulate the production of tumor-specific ribosomes is yet to be determined. If such ribosomes exist, this will provide a completely new area of investigation with potential for novel strategies to target tumor cells.
Author Contributions
Conceptualization, M.C.-S.; methodology I.N.B. and M.C.L.-N.; software, I.N.B.; validation, M.C.L.-N. and M.C.-S.; formal analysis, I.N.B.; investigation I.N.B., M.C.L.-N. and M.C.-S.; data curation, I.N.B.; writing—original draft preparation, I.N.B., M.C.L.-N. and M.C.-S.; writing—review and editing, I.N.B., M.C.L.-N. and M.C.-S.; visualization, I.N.B.; supervision, M.C.L.-N. and M.C.-S.; project administration, M.C.-S.; funding acquisition, M.C.-S. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by Welch I-2058-20210327, NCI R01CA245548, the UTSW Kidney Cancer SPORE Career Enhancement Program [P50CA196516/MCS], and a training fellowship supported by a Hamon Center for Regenerative Science and Medicine to M.L.N. I.N.B. is supported by training grant RP210041. M.C.S. is the Virginia Murchison Linthicum Scholar in Medical Research. The authors acknowledge the UT Southwestern Live Cell Imaging Core Facility (Harold C. Simmons Cancer Center), supported in part by an NCI Cancer Center Support Grant, 1P30 CA142543-01.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to the Electron Microscopy Core Facility at UTSW, directed by Kate Luby-Phelps.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Montanaro, L.; Treré, D.; Derenzini, M. Nucleolus, ribosomes, and cancer. Am. J. Pathol. 2008, 173, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Derenzini, M.; Trerè, D.; Pession, A.; Montanaro, L.; Sirri, V.; Ochs, R.L. Nucleolar function and size in cancer cells. Am. J. Pathol. 1998, 152, 1291–1297. [Google Scholar]
- Thomas, G. An encore for ribosome biogenesis in the control of cell proliferation. Nat. Cell Biol. 2000, 2, E71–E72. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Verdun, D. Assembly and disassembly of the nucleolus during the cell cycle. Nucleus 2011, 2, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, A.S.; Warburton, D.; Atwood, K.C. Location of ribosomal DNA in the human chromosome complement. Proc. Natl. Acad. Sci. USA 1972, 69, 3394–3398. [Google Scholar] [CrossRef] [Green Version]
- Bersaglieri, C.; Santoro, R. Genome Organization in and around the Nucleolus. Cells 2019, 8, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsekrekou, M.; Stratigi, K.; Chatzinikolaou, G. The Nucleolus: In Genome Maintenance and Repair. Int. J. Mol. Sci. 2017, 18, 1411. [Google Scholar] [CrossRef] [Green Version]
- Anger, A.M.; Armache, J.P.; Berninghausen, O.; Habeck, M.; Subklewe, M.; Wilson, D.N.; Beckmann, R. Structures of the human and Drosophila 80S ribosome. Nature 2013, 497, 80–85. [Google Scholar] [CrossRef]
- Martinez-Calvillo, S.; Florencio-Martinez, L.E.; Nepomuceno-Mejia, T. Nucleolar Structure and Function in Trypanosomatid Protozoa. Cells 2019, 8, 421. [Google Scholar] [CrossRef] [Green Version]
- Thiry, M.; Lafontaine, D.L. Birth of a nucleolus: The evolution of nucleolar compartments. Trends Cell Biol. 2005, 15, 194–199. [Google Scholar] [CrossRef]
- Thiry, M.; Lamaye, F.; Lafontaine, D.L. The nucleolus: When 2 became 3. Nucleus 2011, 2, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidian, A.; Koshel, E.; Dyomin, A.; Galkina, S.; Saifitdinova, A.; Gaginskaya, E. On some structural and evolutionary aspects of rDNA amplification in oogenesis of Trachemys scripta turtles. Cell Tissue Res. 2021, 383, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Verdun, D.; Roussel, P.; Thiry, M.; Sirri, V.; Lafontaine, D.L. The nucleolus: Structure/function relationship in RNA metabolism. Wiley Interdiscip Rev. RNA 2010, 1, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Ruggero, D. Revisiting the nucleolus: From marker to dynamic integrator of cancer signaling. Sci. Signal. 2012, 5, pe38. [Google Scholar] [CrossRef] [Green Version]
- Destefanis, F.; Manara, V.; Bellosta, P. Myc as a Regulator of Ribosome Biogenesis and Cell Competition: A Link to Cancer. Int. J. Mol. Sci. 2020, 21, 4037. [Google Scholar] [CrossRef]
- Fromont-Racine, M.; Senger, B.; Saveanu, C.; Fasiolo, F. Ribosome assembly in eukaryotes. Gene 2003, 313, 17–42. [Google Scholar] [CrossRef]
- Jantzen, H.M.; Admon, A.; Bell, S.P.; Tjian, R. Nucleolar transcription factor hUBF contains a DNA-binding motif with homology to HMG proteins. Nature 1990, 344, 830–836. [Google Scholar] [CrossRef]
- Grummt, I. Life on a planet of its own: Regulation of RNA polymerase I transcription in the nucleolus. Genes Dev. 2003, 17, 1691–1702. [Google Scholar] [CrossRef] [Green Version]
- Jin, R.; Zhou, W. TIF-IA: An oncogenic target of pre-ribosomal RNA synthesis. Biochim. Biophys. Acta 2016, 1866, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Szymanski, M.; Barciszewska, M.Z.; Erdmann, V.A.; Barciszewski, J. 5S Ribosomal RNA Database. Nucleic Acids Res. 2002, 30, 176–178. [Google Scholar] [CrossRef]
- Ciganda, M.; Williams, N. Eukaryotic 5S rRNA biogenesis. Wiley Interdiscip Rev. RNA 2011, 2, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Lafontaine, D.L.J. ‘View From A Bridge’: A New Perspective on Eukaryotic rRNA Base Modification. Trends Biochem. Sci. 2015, 40, 560–575. [Google Scholar] [CrossRef]
- Penzo, M.; Guerrieri, A.N.; Zacchini, F.; Treré, D.; Montanaro, L. RNA Pseudouridylation in Physiology and Medicine: For Better and for Worse. Genes 2017, 8, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polikanov, Y.S.; Melnikov, S.V.; Söll, D.; Steitz, T.A. Structural insights into the role of rRNA modifications in protein synthesis and ribosome assembly. Nat. Struct. Mol. Biol. 2015, 22, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Sloan, K.E.; Warda, A.S.; Sharma, S.; Entian, K.D.; Lafontaine, D.L.J.; Bohnsack, M.T. Tuning the ribosome: The influence of rRNA modification on eukaryotic ribosome biogenesis and function. RNA Biol. 2017, 14, 1138–1152. [Google Scholar] [CrossRef]
- Jackman, J.E.; Alfonzo, J.D. Transfer RNA modifications: Nature’s combinatorial chemistry playground. Wiley Interdiscip Rev. RNA 2013, 4, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.; Zomerdijk, J.C. RNA-polymerase-I-directed rDNA transcription, life and works. Trends Biochem. Sci. 2005, 30, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Cheng, W.; Parlato, R.; Guo, X.; Cui, X.; Dai, C.; Xu, L.; Zhu, J.; Zhu, M.; Luo, K.; et al. Nucleolar stress induces a senescence-like phenotype in smooth muscle cells and promotes development of vascular degeneration. Aging 2020, 12, 22174–22198. [Google Scholar] [CrossRef]
- Lopez-Candales, A.; Holmes, D.R.; Liao, S.; Scott, M.J.; Wickline, S.A.; Thompson, R.W. Decreased vascular smooth muscle cell density in medial degeneration of human abdominal aortic aneurysms. Am. J. Pathol. 1997, 150, 993–1007. [Google Scholar] [PubMed]
- Kang, H.; Shin, J.H. Repression of rRNA transcription by PARIS contributes to Parkinson’s disease. Neurobiol. Dis. 2015, 73, 220–228. [Google Scholar] [CrossRef]
- Sanchez, E.; Laplace-Builhe, B.; Mau-Them, F.T.; Richard, E.; Goldenberg, A.; Toler, T.L.; Guignard, T.; Gatinois, V.; Vincent, M.; Blanchet, C.; et al. POLR1B and neural crest cell anomalies in Treacher Collins syndrome type 4. Genet. Med. 2020, 22, 547–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauwerse, J.G.; Dixon, J.; Seland, S.; Ruivenkamp, C.A.; van Haeringen, A.; Hoefsloot, L.H.; Peters, D.J.; Boers, A.C.; Daumer-Haas, C.; Maiwald, R.; et al. Mutations in genes encoding subunits of RNA polymerases I and III cause Treacher Collins syndrome. Nat. Genet. 2011, 43, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.; Gupta, A.; Dutta, A.; Potluri, P.; Batti, B. A Review of Diamond-Blackfan Anemia: Current Evidence on Involved Genes and Treatment Modalities. Cureus 2020, 12, e10019. [Google Scholar] [CrossRef]
- Farrar, J.E.; Quarello, P.; Fisher, R.; O’Brien, K.A.; Aspesi, A.; Parrella, S.; Henson, A.L.; Seidel, N.E.; Atsidaftos, E.; Prakash, S.; et al. Exploiting pre-rRNA processing in Diamond Blackfan anemia gene discovery and diagnosis. Am. J. Hematol. 2014, 89, 985–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noel, C.B. Diamond-Blackfan anemia RPL35A: A case report. J. Med. Case Rep. 2019, 13, 185. [Google Scholar] [CrossRef]
- Warda, A.S.; Freytag, B.; Haag, S.; Sloan, K.E.; Gorlich, D.; Bohnsack, M.T. Effects of the Bowen-Conradi syndrome mutation in EMG1 on its nuclear import, stability and nucleolar recruitment. Hum. Mol. Genet. 2016, 25, 5353–5364. [Google Scholar] [CrossRef]
- Farooq, M.; Lindbaek, L.; Krogh, N.; Doganli, C.; Keller, C.; Monnich, M.; Goncalves, A.B.; Sakthivel, S.; Mang, Y.; Fatima, A.; et al. RRP7A links primary microcephaly to dysfunction of ribosome biogenesis, resorption of primary cilia, and neurogenesis. Nat. Commun. 2020, 11, 5816. [Google Scholar] [CrossRef]
- Ruggero, D.; Pandolfi, P.P. Does the ribosome translate cancer? Nat. Rev. Cancer 2003, 3, 179–192. [Google Scholar] [CrossRef]
- White, R.J. RNA polymerases I and III, growth control and cancer. Nat. Rev. Mol. Cell Biol. 2005, 6, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Tsai, R.Y.; Pederson, T. Connecting the nucleolus to the cell cycle and human disease. FASEB J. 2014, 28, 3290–3296. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Lemos, B. Ribosomal DNA copy number amplification and loss in human cancers is linked to tumor genetic context, nucleolus activity, and proliferation. PLoS Genet. 2017, 13, e1006994. [Google Scholar] [CrossRef]
- Feng, L.; Du, J.; Yao, C.; Jiang, Z.; Li, T.; Zhang, Q.; Guo, X.; Yu, M.; Xia, H.; Shi, L.; et al. Ribosomal DNA copy number is associated with P53 status and levels of heavy metals in gastrectomy specimens from gastric cancer patients. Environ. Int 2020, 138, 105593. [Google Scholar] [CrossRef] [PubMed]
- Derenzini, M.; Trerè, D.; Pession, A.; Govoni, M.; Sirri, V.; Chieco, P. Nucleolar size indicates the rapidity of cell proliferation in cancer tissues. J. Pathol. 2000, 191, 181–186. [Google Scholar] [CrossRef]
- Bywater, M.J.; Poortinga, G.; Sanij, E.; Hein, N.; Peck, A.; Cullinane, C.; Wall, M.; Cluse, L.; Drygin, D.; Anderes, K.; et al. Inhibition of RNA polymerase I as a therapeutic strategy to promote cancer-specific activation of p53. Cancer Cell 2012, 22, 51–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arabi, A.; Wu, S.; Ridderstrale, K.; Bierhoff, H.; Shiue, C.; Fatyol, K.; Fahlen, S.; Hydbring, P.; Soderberg, O.; Grummt, I.; et al. c-myc associates with ribosomal DNA and activates RNA polymerase I transcription. Nat. Cell Biol. 2005, 7, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Grandori, C.; Gomez-Roman, N.; Felton-Edkins, Z.A.; Ngouenet, C.; Galloway, D.A.; Eisenman, R.N.; White, R.J. c-myc binds to human ribosomal DNA and stimulates transcription of rRNA genes by RNA polymerase I. Nat. Cell Biol. 2005, 7, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Grewal, S.S.; Li, L.; Orian, A.; Eisenman, R.N.; Edgar, B.A. Myc-dependent regulation of ribosomal RNA synthesis during Drosophila development. Nat. Cell Biol. 2005, 7, 295–302. [Google Scholar] [CrossRef]
- Lee, C.M.; Reddy, E.P. The v-myc oncogene. Oncogene 1999, 18, 2997–3003. [Google Scholar] [CrossRef]
- Roussel, M.; Saule, S.; Lagrou, C.; Rommens, C.; Beug, H.; Graf, T.; Stehelin, D. Three new types of viral oncogene of cellular origin specific for haematopoietic cell transformation. Nature 1979, 281, 452–455. [Google Scholar] [CrossRef]
- Sheiness, D.; Bishop, J.M. DNA and RNA from uninfected vertebrate cells contain nucleotide sequences related to the putative transforming gene of avian myelocytomatosis virus. J. Virol. 1979, 31, 514–521. [Google Scholar] [CrossRef] [Green Version]
- Payne, G.S.; Bishop, J.M.; Varmus, H.E. Multiple arrangements of viral DNA and an activated host oncogene in bursal lymphomas. Nature 1982, 295, 209–214. [Google Scholar] [CrossRef]
- Hayward, W.S.; Neel, B.G.; Astrin, S.M. Activation of a cellular onc gene by promoter insertion in ALV-induced lymphoid leukosis. Nature 1981, 290, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Dalla-Favera, R.; Bregni, M.; Erikson, J.; Patterson, D.; Gallo, R.C.; Croce, C.M. Human c-myc onc gene is located on the region of chromosome 8 that is translocated in Burkitt lymphoma cells. Proc. Natl. Acad. Sci. USA 1982, 79, 7824–7827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelengaris, S.; Khan, M.; Evan, G. c-myc: More than just a matter of life and death. Nat. Rev. Cancer 2002, 2, 764–776. [Google Scholar] [CrossRef] [PubMed]
- Lafita-Navarro, M.C.; Kim, M.; Borenstein-Auerbach, N.; Venkateswaran, N.; Hao, Y.H.; Ray, R.; Brabletz, T.; Scaglioni, P.P.; Shay, J.W.; Conacci-Sorrell, M. The aryl hydrocarbon receptor regulates nucleolar activity and protein synthesis in Myc-expressing cells. Genes Dev. 2018, 32, 1303–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Riggelen, J.; Yetil, A.; Felsher, D.W. Myc as a regulator of ribosome biogenesis and protein synthesis. Nat. Rev. Cancer 2010, 10, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Dang, C.V. Myc on the path to cancer. Cell 2012, 149, 22–35. [Google Scholar] [CrossRef] [Green Version]
- Cascón, A.; Robledo, M. MAX and Myc: A heritable breakup. Cancer Res. 2012, 72, 3119–3124. [Google Scholar] [CrossRef] [Green Version]
- Eilers, M.; Eisenman, R.N. Myc’s broad reach. Genes Dev. 2008, 22, 2755–2766. [Google Scholar] [CrossRef] [Green Version]
- Grandori, C.; Cowley, S.M.; James, L.P.; Eisenman, R.N. The Myc/Max/Mad network and the transcriptional control of cell behavior. Annu. Rev. Cell Dev. Biol. 2000, 16, 653–699. [Google Scholar] [CrossRef]
- Sammak, S.; Hamdani, N.; Gorrec, F.; Allen, M.D.; Freund, S.M.V.; Bycroft, M.; Zinzalla, G. Crystal Structures and Nuclear Magnetic Resonance Studies of the Apo Form of the c-myc:MAX bHLHZip Complex Reveal a Helical Basic Region in the Absence of DNA. Biochemistry 2019, 58, 3144–3154. [Google Scholar] [CrossRef] [Green Version]
- Kretzner, L.; Blackwood, E.M.; Eisenman, R.N. Myc and Max proteins possess distinct transcriptional activities. Nature 1992, 359, 426–429. [Google Scholar] [CrossRef]
- Amati, B.; Brooks, M.W.; Levy, N.; Littlewood, T.D.; Evan, G.I.; Land, H. Oncogenic activity of the c-myc protein requires dimerization with Max. Cell 1993, 72, 233–245. [Google Scholar] [CrossRef]
- Izumi, H.; Imamura, T.; Nagatani, G.; Ise, T.; Murakami, T.; Uramoto, H.; Torigoe, T.; Ishiguchi, H.; Yoshida, Y.; Nomoto, M.; et al. Y box-binding protein-1 binds preferentially to single-stranded nucleic acids and exhibits 3’-->5’ exonuclease activity. Nucleic Acids Res. 2001, 29, 1200–1207. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Roman, N.; Grandori, C.; Eisenman, R.N.; White, R.J. Direct activation of RNA polymerase III transcription by c-myc. Nature 2003, 421, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, N.S.; Ramsbottom, B.A.; Gomez-Roman, N.; Marshall, L.; Cole, P.A.; White, R.J. TRRAP and GCN5 are used by c-myc to activate RNA polymerase III transcription. Proc. Natl. Acad. Sci. USA 2007, 104, 14917–14922. [Google Scholar] [CrossRef] [Green Version]
- Conacci-Sorrell, M.; Ngouenet, C.; Eisenman, R.N. Myc-nick: A cytoplasmic cleavage product of Myc that promotes alpha-tubulin acetylation and cell differentiation. Cell 2010, 142, 480–493. [Google Scholar] [CrossRef] [Green Version]
- Conacci-Sorrell, M.; Ngouenet, C.; Anderson, S.; Brabletz, T.; Eisenman, R.N. Stress-induced cleavage of Myc promotes cancer cell survival. Genes Dev. 2014, 28, 689–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.; Poudel, K.R.; Roh-Johnson, M.; Brabletz, T.; Yu, M.; Borenstein-Auerbach, N.; Grady, W.N.; Bai, J.; Moens, C.B.; Eisenman, R.N.; et al. Myc-nick promotes cell migration by inducing fascin expression and Cdc42 activation. Proc. Natl. Acad. Sci. USA 2016, 113, E5481–E5490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flinn, E.M.; Busch, C.M.; Wright, A.P. Myc boxes, which are conserved in Myc family proteins, are signals for protein degradation via the proteasome. Mol. Cell. Biol. 1998, 18, 5961–5969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hann, S.R.; Thompson, C.B.; Eisenman, R.N. c-myc oncogene protein synthesis is independent of the cell cycle in human and avian cells. Nature 1985, 314, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Welcker, M.; Orian, A.; Grim, J.E.; Grim, J.A.; Eisenman, R.N.; Clurman, B.E. A nucleolar isoform of the Fbw7 ubiquitin ligase regulates c-myc and cell size. Curr. Biol. 2004, 14, 1852–1857. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Wright, J.B.; Gerber, S.A.; Cole, M.D. Myc protein is stabilized by suppression of a novel E3 ligase complex in cancer cells. Genes Dev. 2010, 24, 1236–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahram, F.; von der Lehr, N.; Cetinkaya, C.; Larsson, L.G. c-myc hot spot mutations in lymphomas result in inefficient ubiquitination and decreased proteasome-mediated turnover. Blood 2000, 95, 2104–2110. [Google Scholar] [CrossRef] [PubMed]
- Ekholm-Reed, S.; Spruck, C.H.; Sangfelt, O.; van Drogen, F.; Mueller-Holzner, E.; Widschwendter, M.; Zetterberg, A.; Reed, S.I.; Reed, S.E. Mutation of hCDC4 leads to cell cycle deregulation of cyclin E in cancer. Cancer Res. 2004, 64, 795–800. [Google Scholar] [CrossRef] [Green Version]
- Spruck, C.H.; Strohmaier, H.; Sangfelt, O.; Müller, H.M.; Hubalek, M.; Müller-Holzner, E.; Marth, C.; Widschwendter, M.; Reed, S.I. hCDC4 gene mutations in endometrial cancer. Cancer Res. 2002, 62, 4535–4539. [Google Scholar]
- Rajagopalan, H.; Jallepalli, P.V.; Rago, C.; Velculescu, V.E.; Kinzler, K.W.; Vogelstein, B.; Lengauer, C. Inactivation of hCDC4 can cause chromosomal instability. Nature 2004, 428, 77–81. [Google Scholar] [CrossRef]
- Koepp, D.M.; Schaefer, L.K.; Ye, X.; Keyomarsi, K.; Chu, C.; Harper, J.W.; Elledge, S.J. Phosphorylation-dependent ubiquitination of cyclin E by the SCFFbw7 ubiquitin ligase. Science 2001, 294, 173–177. [Google Scholar] [CrossRef]
- Gupta-Rossi, N.; Le Bail, O.; Gonen, H.; Brou, C.; Logeat, F.; Six, E.; Ciechanover, A.; Israël, A. Functional interaction between SEL-10, an F-box protein, and the nuclear form of activated Notch1 receptor. J. Biol. Chem. 2001, 276, 34371–34378. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Lyapina, S.; Das, I.; Li, J.; Gurney, M.; Pauley, A.; Chui, I.; Deshaies, R.J.; Kitajewski, J. SEL-10 is an inhibitor of notch signaling that targets notch for ubiquitin-mediated protein degradation. Mol. Cell. Biol. 2001, 21, 7403–7415. [Google Scholar] [CrossRef] [Green Version]
- Nateri, A.S.; Riera-Sans, L.; Da Costa, C.; Behrens, A. The ubiquitin ligase SCFFbw7 antagonizes apoptotic JNK signaling. Science 2004, 303, 1374–1378. [Google Scholar] [CrossRef] [Green Version]
- Welcker, M.; Orian, A.; Jin, J.; Grim, J.E.; Grim, J.A.; Harper, J.W.; Eisenman, R.N.; Clurman, B.E. The Fbw7 tumor suppressor regulates glycogen synthase kinase 3 phosphorylation-dependent c-myc protein degradation. Proc. Natl. Acad. Sci. USA 2004, 101, 9085–9090. [Google Scholar] [CrossRef] [Green Version]
- Yada, M.; Hatakeyama, S.; Kamura, T.; Nishiyama, M.; Tsunematsu, R.; Imaki, H.; Ishida, N.; Okumura, F.; Nakayama, K.; Nakayama, K.I. Phosphorylation-dependent degradation of c-myc is mediated by the F-box protein Fbw7. EMBO J. 2004, 23, 2116–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, J.; de Lange, T.; Ramsay, G.; Jakobovits, E.; Bishop, J.M.; Varmus, H.; Lee, W. Definition of regions in human c-myc that are involved in transformation and nuclear localization. Mol. Cell. Biol. 1987, 7, 1697–1709. [Google Scholar] [CrossRef]
- Lee, K.K.; Workman, J.L. Histone acetyltransferase complexes: One size doesn’t fit all. Nat. Rev. Mol. Cell Biol. 2007, 8, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Cole, M.D.; Cowling, V.H. Transcription-independent functions of Myc: Regulation of translation and DNA replication. Nat. Rev. Mol. Cell Biol. 2008, 9, 810–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, S.R.; Schroeder, M.; Fernandez, P.; Taubert, S.; Amati, B. Binding of c-myc to chromatin mediates mitogen-induced acetylation of histone H4 and gene activation. Genes Dev. 2001, 15, 2069–2082. [Google Scholar] [CrossRef] [Green Version]
- McMahon, S.B.; Van Buskirk, H.A.; Dugan, K.A.; Copeland, T.D.; Cole, M.D. The novel ATM-related protein TRRAP is an essential cofactor for the c-myc and E2F oncoproteins. Cell 1998, 94, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Vervoorts, J.; Lüscher-Firzlaff, J.M.; Rottmann, S.; Lilischkis, R.; Walsemann, G.; Dohmann, K.; Austen, M.; Lüscher, B. Stimulation of c-myc transcriptional activity and acetylation by recruitment of the cofactor CBP. EMBO Rep. 2003, 4, 484–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbst, A.; Hemann, M.T.; Tworkowski, K.A.; Salghetti, S.E.; Lowe, S.W.; Tansey, W.P. A conserved element in Myc that negatively regulates its proapoptotic activity. EMBO Rep. 2005, 6, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Cowling, V.H.; Chandriani, S.; Whitfield, M.L.; Cole, M.D. A conserved Myc protein domain, MBIV, regulates DNA binding, apoptosis, transformation, and G2 arrest. Mol. Cell. Biol. 2006, 26, 4226–4239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panov, K.I.; Friedrich, J.K.; Russell, J.; Zomerdijk, J.C. UBF activates RNA polymerase I transcription by stimulating promoter escape. EMBO J. 2006, 25, 3310–3322. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, J.K.; Panov, K.I.; Cabart, P.; Russell, J.; Zomerdijk, J.C. TBP-TAF complex SL1 directs RNA polymerase I pre-initiation complex formation and stabilizes upstream binding factor at the rDNA promoter. J. Biol. Chem. 2005, 280, 29551–29558. [Google Scholar] [CrossRef] [Green Version]
- Shiue, C.N.; Berkson, R.G.; Wright, A.P. c-myc induces changes in higher order rDNA structure on stimulation of quiescent cells. Oncogene 2009, 28, 1833–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiue, C.N.; Arabi, A.; Wright, A.P. Nucleolar organization, growth control and cancer. Epigenetics 2010, 5, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiue, C.N.; Nematollahi-Mahani, A.; Wright, A.P. Myc-induced anchorage of the rDNA IGS region to nucleolar matrix modulates growth-stimulated changes in higher-order rDNA architecture. Nucleic Acids Res. 2014, 42, 5505–5517. [Google Scholar] [CrossRef] [Green Version]
- Patel, J.H.; Loboda, A.P.; Showe, M.K.; Showe, L.C.; McMahon, S.B. Analysis of genomic targets reveals complex functions of Myc. Nat. Rev. Cancer 2004, 4, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Cowling, V.H.; Cole, M.D. Mechanism of transcriptional activation by the Myc oncoproteins. Semin. Cancer Biol. 2006, 16, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Perna, D.; Fagà, G.; Verrecchia, A.; Gorski, M.M.; Barozzi, I.; Narang, V.; Khng, J.; Lim, K.C.; Sung, W.K.; Sanges, R.; et al. Genome-wide mapping of Myc binding and gene regulation in serum-stimulated fibroblasts. Oncogene 2012, 31, 1695–1709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Y.H.; Lafita-Navarro, M.C.; Zacharias, L.; Borenstein-Auerbach, N.; Kim, M.; Barnes, S.; Kim, J.; Shay, J.; DeBerardinis, R.J.; Conacci-Sorrell, M. Induction of LEF1 by Myc activates the WNT pathway and maintains cell proliferation. Cell Commun. Signal. 2019, 17, 129. [Google Scholar] [CrossRef] [Green Version]
- Lafita-Navarro, M.C.; Perez-Castro, L.; Zacharias, L.G.; Barnes, S.; DeBerardinis, R.J.; Conacci-Sorrell, M. The transcription factors aryl hydrocarbon receptor and Myc cooperate in the regulation of cellular metabolism. J. Biol. Chem. 2020, 295, 12398–12407. [Google Scholar] [CrossRef]
- Ruggero, D. The role of Myc-induced protein synthesis in cancer. Cancer Res. 2009, 69, 8839–8843. [Google Scholar] [CrossRef] [Green Version]
- Ginisty, H.; Amalric, F.; Bouvet, P. Nucleolin functions in the first step of ribosomal RNA processing. EMBO J. 1998, 17, 1476–1486. [Google Scholar] [CrossRef] [Green Version]
- Feric, M.; Vaidya, N.; Harmon, T.S.; Mitrea, D.M.; Zhu, L.; Richardson, T.M.; Kriwacki, R.W.; Pappu, R.V.; Brangwynne, C.P. Coexisting Liquid Phases Underlie Nucleolar Subcompartments. Cell 2016, 165, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Mitrea, D.M.; Cika, J.A.; Stanley, C.B.; Nourse, A.; Onuchic, P.L.; Banerjee, P.R.; Phillips, A.H.; Park, C.G.; Deniz, A.A.; Kriwacki, R.W. Self-interaction of NPM1 modulates multiple mechanisms of liquid-liquid phase separation. Nat. Commun. 2018, 9, 842. [Google Scholar] [CrossRef]
- Maggi, L.B.; Kuchenruether, M.; Dadey, D.Y.; Schwope, R.M.; Grisendi, S.; Townsend, R.R.; Pandolfi, P.P.; Weber, J.D. Nucleophosmin serves as a rate-limiting nuclear export chaperone for the Mammalian ribosome. Mol. Cell. Biol. 2008, 28, 7050–7065. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hann, S.R. The Myc-nucleophosmin-ARF network: A complex web unveiled. Cell Cycle 2009, 8, 2703–2707. [Google Scholar] [CrossRef] [Green Version]
- Lafita-Navarro, M.C.; Venkateswaran, N.; Kilgore, J.A.; Kanji, S.; Han, J.; Barnes, S.; Williams, N.S.; Buszczak, M.; Burma, S.; Conacci-Sorrell, M. Inhibition of the de novo pyrimidine biosynthesis pathway limits ribosomal RNA transcription causing nucleolar stress in glioblastoma cells. PLoS Genet. 2020, 16, e1009117. [Google Scholar] [CrossRef] [PubMed]
- Hubackova, S.; Davidova, E.; Boukalova, S.; Kovarova, J.; Bajzikova, M.; Coelho, A.; Terp, M.G.; Ditzel, H.J.; Rohlena, J.; Neuzil, J. Replication and ribosomal stress induced by targeting pyrimidine synthesis and cellular checkpoints suppress p53-deficient tumors. Cell Death Dis. 2020, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.N.; Fan, T.W. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res. 2015, 43, 2466–2485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.C.; Li, F.; Handler, J.; Huang, C.R.; Xiang, Y.; Neretti, N.; Sedivy, J.M.; Zeller, K.I.; Dang, C.V. Global regulation of nucleotide biosynthetic genes by c-Myc. PLoS ONE 2008, 3, e2722. [Google Scholar] [CrossRef]
- Eberhardy, S.R.; Farnham, P.J. c-myc mediates activation of the cad promoter via a post-RNA polymerase II recruitment mechanism. J. Biol. Chem. 2001, 276, 48562–48571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahl, P.B.; Young, R.A. Myc and transcription elongation. Cold Spring Harb. Perspect. Med. 2014, 4, a020990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Li, T.; Price, D.H. RNA polymerase II elongation control. Annu. Rev. Biochem. 2012, 81, 119–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, E.V. The role of c-myc in regulation of translation initiation. Oncogene 2004, 23, 3217–3221. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Boone, D.; Hann, S.R. Nucleophosmin interacts directly with c-myc and controls c-myc-induced hyperproliferation and transformation. Proc. Natl. Acad. Sci. USA 2008, 105, 18794–18799. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Hann, S.R. Nucleophosmin is essential for c-myc nucleolar localization and c-myc-mediated rDNA transcription. Oncogene 2013, 32, 1988–1994. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.S.; Arnold, H.; Sun, X.X.; Sears, R.; Lu, H. Inhibition of c-myc activity by ribosomal protein L11. EMBO J. 2007, 26, 3332–3345. [Google Scholar] [CrossRef]
- Liao, J.M.; Zhou, X.; Gatignol, A.; Lu, H. Ribosomal proteins L5 and L11 co-operatively inactivate c-myc via RNA-induced silencing complex. Oncogene 2014, 33, 4916–4923. [Google Scholar] [CrossRef] [Green Version]
- Burger, K.; Muhl, B.; Harasim, T.; Rohrmoser, M.; Malamoussi, A.; Orban, M.; Kellner, M.; Gruber-Eber, A.; Kremmer, E.; Holzel, M.; et al. Chemotherapeutic drugs inhibit ribosome biogenesis at various levels. J. Biol. Chem. 2010, 285, 12416–12425. [Google Scholar] [CrossRef] [Green Version]
- Hannan, K.M.; Sanij, E.; Rothblum, L.I.; Hannan, R.D.; Pearson, R.B. Dysregulation of RNA polymerase I transcription during disease. Biochim. Biophys. Acta 2013, 1829, 342–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drygin, D.; Rice, W.G.; Grummt, I. The RNA polymerase I transcription machinery: An emerging target for the treatment of cancer. Annu. Rev. Pharm. Toxicol. 2010, 50, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Hein, N.; Hannan, K.M.; George, A.J.; Sanij, E.; Hannan, R.D. The nucleolus: An emerging target for cancer therapy. Trends Mol. Med. 2013, 19, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Andrews, W.J.; Panova, T.; Normand, C.; Gadal, O.; Tikhonova, I.G.; Panov, K.I. Old drug, new target: Ellipticines selectively inhibit RNA polymerase I transcription. J. Biol. Chem. 2013, 288, 4567–4582. [Google Scholar] [CrossRef] [Green Version]
- Catez, F.; Dalla Venezia, N.; Marcel, V.; Zorbas, C.; Lafontaine, D.L.J.; Diaz, J.J. Ribosome biogenesis: An emerging druggable pathway for cancer therapeutics. Biochem. Pharm. 2019, 159, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Derenzini, M.; Montanaro, L.; Trere, D. What the nucleolus says to a tumour pathologist. Histopathology 2009, 54, 753–762. [Google Scholar] [CrossRef]
- Woods, S.J.; Hannan, K.M.; Pearson, R.B.; Hannan, R.D. The nucleolus as a fundamental regulator of the p53 response and a new target for cancer therapy. Biochim. Biophys. Acta 2015, 1849, 821–829. [Google Scholar] [CrossRef]
- Boisvert, F.M.; van Koningsbruggen, S.; Navascues, J.; Lamond, A.I. The multifunctional nucleolus. Nat. Rev. Mol. Cell Biol. 2007, 8, 574–585. [Google Scholar] [CrossRef]
- Carotenuto, P.; Pecoraro, A.; Palma, G.; Russo, G.; Russo, A. Therapeutic Approaches Targeting Nucleolus in Cancer. Cells 2019, 8, 1090. [Google Scholar] [CrossRef] [Green Version]
- Boulon, S.; Westman, B.J.; Hutten, S.; Boisvert, F.M.; Lamond, A.I. The nucleolus under stress. Mol. Cell 2010, 40, 216–227. [Google Scholar] [CrossRef]
- Horn, H.F.; Vousden, K.H. Coping with stress: Multiple ways to activate p53. Oncogene 2007, 26, 1306–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, A.; Wang, Y.; Raje, H.; Rosby, R.; DiMario, P. Nucleolar stress with and without p53. Nucleus 2014, 5, 402–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kofuji, S.; Hirayama, A.; Eberhardt, A.O.; Kawaguchi, R.; Sugiura, Y.; Sampetrean, O.; Ikeda, Y.; Warren, M.; Sakamoto, N.; Kitahara, S.; et al. IMP dehydrogenase-2 drives aberrant nucleolar activity and promotes tumorigenesis in glioblastoma. Nat. Cell Biol. 2019, 21, 1003–1014. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Ni, M.; Chalishazar, M.D.; Huffman, K.E.; Kim, J.; Cai, L.; Shi, X.; Cai, F.; Zacharias, L.G.; Ireland, A.S.; et al. Inosine Monophosphate Dehydrogenase Dependence in a Subset of Small Cell Lung Cancers. Cell Metab. 2018, 28, 369–382 e365. [Google Scholar] [CrossRef] [Green Version]
- Bailly, A.; Perrin, A.; Bou Malhab, L.J.; Pion, E.; Larance, M.; Nagala, M.; Smith, P.; O’Donohue, M.F.; Gleizes, P.E.; Zomerdijk, J.; et al. The NEDD8 inhibitor MLN4924 increases the size of the nucleolus and activates p53 through the ribosomal-Mdm2 pathway. Oncogene 2016, 35, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Swords, R.T.; Erba, H.P.; DeAngelo, D.J.; Bixby, D.L.; Altman, J.K.; Maris, M.; Hua, Z.; Blakemore, S.J.; Faessel, H.; Sedarati, F.; et al. Pevonedistat (MLN4924), a First-in-Class NEDD8-activating enzyme inhibitor, in patients with acute myeloid leukaemia and myelodysplastic syndromes: A phase 1 study. Br. J. Haematol. 2015, 169, 534–543. [Google Scholar] [CrossRef] [Green Version]
- Genuth, N.R.; Barna, M. The Discovery of Ribosome Heterogeneity and Its Implications for Gene Regulation and Organismal Life. Mol. Cell 2018, 71, 364–374. [Google Scholar] [CrossRef] [Green Version]
- CRICK, F.H. On protein synthesis. Symp. Soc. Exp. Biol. 1958, 12, 138–163. [Google Scholar]
- BRENNER, S.; JACOB, F.; MESELSON, M. An unstable intermediate carrying information from genes to ribosomes for protein synthesis. Nature 1961, 190, 576–581. [Google Scholar] [CrossRef]
- Dinman, J.D. Pathways to Specialized Ribosomes: The Brussels Lecture. J. Mol. Biol. 2016, 428, 2186–2194. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Barna, M. Translating the genome in time and space: Specialized ribosomes, RNA regulons, and RNA-binding proteins. Annu. Rev. Cell Dev. Biol. 2015, 31, 31–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, S.; Barna, M. Specialized ribosomes: A new frontier in gene regulation and organismal biology. Nat. Rev. Mol. Cell Biol. 2012, 13, 355–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Röst, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide. Mol. Cell 2017, 67, 71–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guimaraes, J.C.; Zavolan, M. Patterns of ribosomal protein expression specify normal and malignant human cells. Genome Biol. 2016, 17, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.; Warner, J.R. Ribosome-omics of the human ribosome. RNA 2014, 20, 1004–1013. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.; Thomas, G.; Volarević, S. Ribosome biogenesis in cancer: New players and therapeutic avenues. Nat. Rev. Cancer 2018, 18, 51–63. [Google Scholar] [CrossRef]
- Wool, I.G. Extraribosomal functions of ribosomal proteins. Trends Biochem. Sci. 1996, 21, 164–165. [Google Scholar] [CrossRef]
- Lai, M.D.; Xu, J. Ribosomal proteins and colorectal cancer. Curr. Genom. 2007, 8, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Golomb, L.; Volarevic, S.; Oren, M. p53 and ribosome biogenesis stress: The essentials. FEBS Lett. 2014, 588, 2571–2579. [Google Scholar] [CrossRef]
- Russo, A.; Russo, G. Ribosomal Proteins Control or Bypass p53 during Nucleolar Stress. Int. J. Mol. Sci. 2017, 18, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.; Lee, S.Y.; Gutierrez, A.; Perrigoue, J.; Thapa, R.J.; Tu, Z.; Jeffers, J.R.; Rhodes, M.; Anderson, S.; Oravecz, T.; et al. Inactivation of ribosomal protein L22 promotes transformation by induction of the stemness factor, Lin28B. Blood 2012, 120, 3764–3773. [Google Scholar] [CrossRef] [PubMed]
- Pogue-Geile, K.; Geiser, J.R.; Shu, M.; Miller, C.; Wool, I.G.; Meisler, A.I.; Pipas, J.M. Ribosomal protein genes are overexpressed in colorectal cancer: Isolation of a cDNA clone encoding the human S3 ribosomal protein. Mol. Cell. Biol. 1991, 11, 3842–3849. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The nucleolus contains three distinct compartments responsible for pre-rRNA transcription, rRNA processing, and ribosome subunit assembly. The nucleoli are located within the nucleus (shown in the electron microscopy photo to the left) and are comprised of three sub-compartments: the fibrillary centers (FC), the dense fibrillar component (DFC), and the granular component (GC). The transcription of rDNA occurs in the FC upon the binding of selectivity factor 1 (SL1), which leads to the activation of the cofactors upstream binding factor (UBF) and TIF-1A, initiating RNAPolI to transcribe rDNA into 47S pre-rRNA. The 47S pre-rRNA is processed into 18S, 5.8S, and 28S rRNA in the DFC. Ribosome maturation proceeds in the GC, where additional RPs are wrapped around rRNAs. Figure created with Biorender.com (accessed on 31 January 2022).
Figure 1.
The nucleolus contains three distinct compartments responsible for pre-rRNA transcription, rRNA processing, and ribosome subunit assembly. The nucleoli are located within the nucleus (shown in the electron microscopy photo to the left) and are comprised of three sub-compartments: the fibrillary centers (FC), the dense fibrillar component (DFC), and the granular component (GC). The transcription of rDNA occurs in the FC upon the binding of selectivity factor 1 (SL1), which leads to the activation of the cofactors upstream binding factor (UBF) and TIF-1A, initiating RNAPolI to transcribe rDNA into 47S pre-rRNA. The 47S pre-rRNA is processed into 18S, 5.8S, and 28S rRNA in the DFC. Ribosome maturation proceeds in the GC, where additional RPs are wrapped around rRNAs. Figure created with Biorender.com (accessed on 31 January 2022).

Figure 2.
Transformed cells have larger nucleoli and increased ribosome production. The nucleolus increases in size and density to accommodate the need for ribosome production. In transformed cells frequently due to hyperactivation of MYC, nucleoli are larger in size and darker in color, indicating higher activity, which results in an increase in ribosome number. Containing a larger number of ribosomes amplifies mRNA translation and ultimately leads to an increase in cell growth. Figure created with Biorender.com (accessed on 5 January 2022).
Figure 2.
Transformed cells have larger nucleoli and increased ribosome production. The nucleolus increases in size and density to accommodate the need for ribosome production. In transformed cells frequently due to hyperactivation of MYC, nucleoli are larger in size and darker in color, indicating higher activity, which results in an increase in ribosome number. Containing a larger number of ribosomes amplifies mRNA translation and ultimately leads to an increase in cell growth. Figure created with Biorender.com (accessed on 5 January 2022).

Figure 3.
MYC heterodimerizes with MAX and increases cell proliferation. (A) Proliferation curve of HO15.19 myc-/- rat fibroblasts expressing empty vector or MYC. Cells were seeded and counted for 5 days. (B) The structure of the basic helix-loop-helix and leucine zipper (bHLH-LZ) domains of the heterodimer MYC-MAX and DNA (PDBe-KD), https://www.ebi.ac.uk/pdbe/pdbe-kb/proteins/P01106/interactions, (accessed on 28 November 2021). (C) Schematic representation of the MYC protein sequence with MYC Boxes (MBI, MBII, MBIII, MBIV), nuclear localization signal (NLS), and bHLHLZip domain on the C-terminus where DNA and MAX interacts. Figure created using Biorender.com (accessed on 28 January 2022).
Figure 3.
MYC heterodimerizes with MAX and increases cell proliferation. (A) Proliferation curve of HO15.19 myc-/- rat fibroblasts expressing empty vector or MYC. Cells were seeded and counted for 5 days. (B) The structure of the basic helix-loop-helix and leucine zipper (bHLH-LZ) domains of the heterodimer MYC-MAX and DNA (PDBe-KD), https://www.ebi.ac.uk/pdbe/pdbe-kb/proteins/P01106/interactions, (accessed on 28 November 2021). (C) Schematic representation of the MYC protein sequence with MYC Boxes (MBI, MBII, MBIII, MBIV), nuclear localization signal (NLS), and bHLHLZip domain on the C-terminus where DNA and MAX interacts. Figure created using Biorender.com (accessed on 28 January 2022).

Figure 4.
Ribosomal biogenesis and structural ribosome components are upregulated in MYC-expressing cells. (A). Schematic of the interaction of MYC and MAX with the transcription machinery that drives the expression of regulatory and structural genes necessary for ribosome biogenesis. (B). Heatmap of myc-/- expressing empty vector or reconstituted with MYC. Data are extracted from published RNAseq [55] for nucleolar genes. (C). Pie chart showing that MYC increased the transcription of 38% of the structural components of the small ribosome subunit and 61% of the large subunit. Data were obtained by comparing the expression of structural ribosome genes in myc-/- expressing empty vector or reconstituted with MYC using a cutoff of Log2 fold change of and with adjusted p-value of . (D). Heatmap of myc-/- expressing empty vector or reconstituted by MYC. Data was extracted from published RNAseq [55] for structural RPs. Heatmaps were generated by MetaboAnalyst 5.0 (https://www.metaboanalyst.ca/, accessed on 10 December 2021). Figure created using Biorender.com, (accessed on 31 January 2022).
Figure 4.
Ribosomal biogenesis and structural ribosome components are upregulated in MYC-expressing cells. (A). Schematic of the interaction of MYC and MAX with the transcription machinery that drives the expression of regulatory and structural genes necessary for ribosome biogenesis. (B). Heatmap of myc-/- expressing empty vector or reconstituted with MYC. Data are extracted from published RNAseq [55] for nucleolar genes. (C). Pie chart showing that MYC increased the transcription of 38% of the structural components of the small ribosome subunit and 61% of the large subunit. Data were obtained by comparing the expression of structural ribosome genes in myc-/- expressing empty vector or reconstituted with MYC using a cutoff of Log2 fold change of and with adjusted p-value of . (D). Heatmap of myc-/- expressing empty vector or reconstituted by MYC. Data was extracted from published RNAseq [55] for structural RPs. Heatmaps were generated by MetaboAnalyst 5.0 (https://www.metaboanalyst.ca/, accessed on 10 December 2021). Figure created using Biorender.com, (accessed on 31 January 2022).

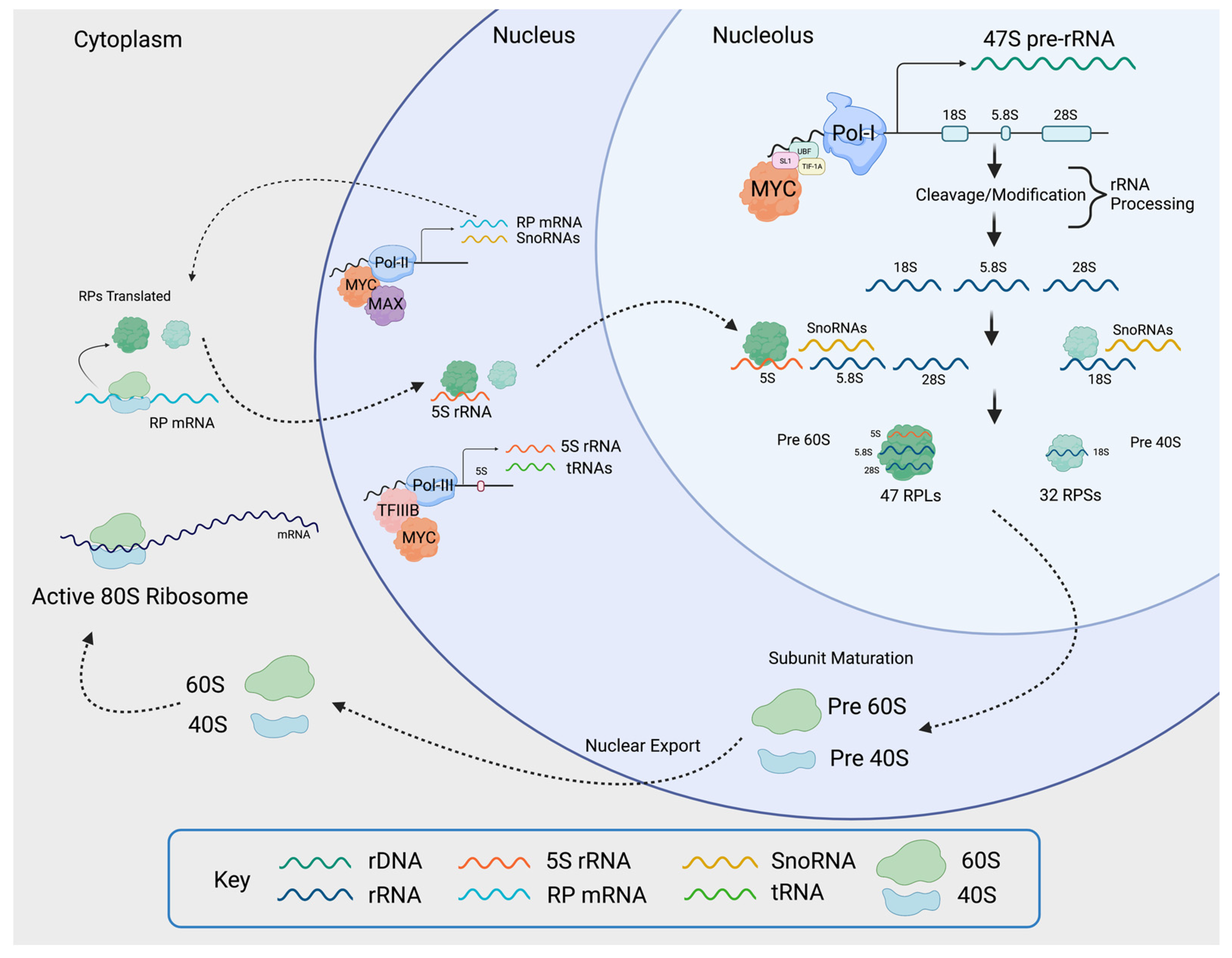
Figure 5.
MYC induces ribosomal biogenesis processes. MYC heterodimerizes with MAX and promotes RNAPolI activity by binding to the rDNA promoter, as well as by activating the expression of selectivity factor 1 (SL1), which binds other RNAPolI cofactors such as upstream binding factor (UBF) and TIF-1A. The rDNA is transcribed into 47S pre-rRNA in the nucleolus. The pre-rRNA is processed and cleaved into 18S, 5.8S, and 28S rRNA. MYC-MAX simultaneously enhances RNAPolII activity by binding to RNAPolII-regulated promoters, as well as RNAPolIII activity by inducing the expression of the RNAPolIII cofactor TFIIIB. This yields RNAPolII-driven small RPs (RPS) mRNA, large RPs (RPL) mRNA, RNAPolIII-driven snoRNAs, and 5S rRNA. The mRNAs are transported into the cytoplasm where mature ribosomes translate them into small and large RPs. Once translated, small RPs are imported into the nucleolus. Large RPs are first imported into the nucleus, where they interact with 5S rRNA, and then to the nucleolus. In the nucleolus, the pre-rRNAs are modified and processed with the help of snoRNAs. As they are maturing, the rRNA wrap around the RPs, creating the pre-40 and pre-60 subunits which are exported to the cytoplasm for the final maturation step. Once maturation is complete in the cytoplasm, the active 80S ribosomes are formed. The small 40S subunit comprises 18S rRNA and 32 small RPs, and the large 60S subunit comprises 5S, 5.8S, and 28S rRNA and 47 large RPs. Figure created using Biorender.com (accessed on 31 January 2022).
Figure 5.
MYC induces ribosomal biogenesis processes. MYC heterodimerizes with MAX and promotes RNAPolI activity by binding to the rDNA promoter, as well as by activating the expression of selectivity factor 1 (SL1), which binds other RNAPolI cofactors such as upstream binding factor (UBF) and TIF-1A. The rDNA is transcribed into 47S pre-rRNA in the nucleolus. The pre-rRNA is processed and cleaved into 18S, 5.8S, and 28S rRNA. MYC-MAX simultaneously enhances RNAPolII activity by binding to RNAPolII-regulated promoters, as well as RNAPolIII activity by inducing the expression of the RNAPolIII cofactor TFIIIB. This yields RNAPolII-driven small RPs (RPS) mRNA, large RPs (RPL) mRNA, RNAPolIII-driven snoRNAs, and 5S rRNA. The mRNAs are transported into the cytoplasm where mature ribosomes translate them into small and large RPs. Once translated, small RPs are imported into the nucleolus. Large RPs are first imported into the nucleus, where they interact with 5S rRNA, and then to the nucleolus. In the nucleolus, the pre-rRNAs are modified and processed with the help of snoRNAs. As they are maturing, the rRNA wrap around the RPs, creating the pre-40 and pre-60 subunits which are exported to the cytoplasm for the final maturation step. Once maturation is complete in the cytoplasm, the active 80S ribosomes are formed. The small 40S subunit comprises 18S rRNA and 32 small RPs, and the large 60S subunit comprises 5S, 5.8S, and 28S rRNA and 47 large RPs. Figure created using Biorender.com (accessed on 31 January 2022).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Brown, I.N.; Lafita-Navarro, M.C.; Conacci-Sorrell, M. Regulation of Nucleolar Activity by MYC. Cells 2022, 11, 574. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030574
AMA Style
Brown IN, Lafita-Navarro MC, Conacci-Sorrell M. Regulation of Nucleolar Activity by MYC. Cells. 2022; 11(3):574. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030574
Chicago/Turabian StyleBrown, Isabella N., M. Carmen Lafita-Navarro, and Maralice Conacci-Sorrell. 2022. "Regulation of Nucleolar Activity by MYC" Cells 11, no. 3: 574. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11030574
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.