
The Effects of Prenatal Exposure to Pregabalin on the Development of Ventral Midbrain Dopaminergic Neurons

 ,
,  , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
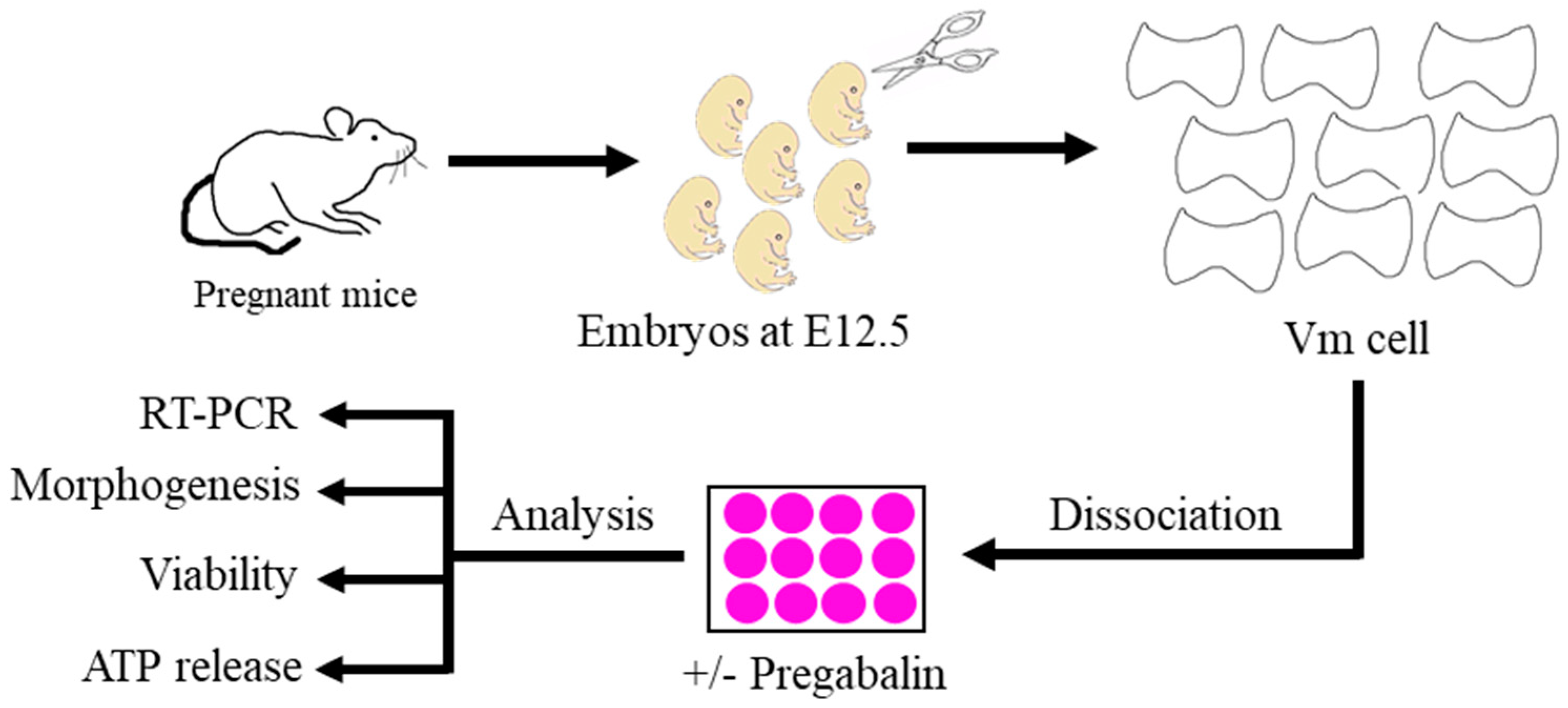
2.1. Isolation of Primary Mouse Embryonic Ventral Midbrain Dopaminergic Neurons
2.2. Three-Dimensional Neuronal CELL Culture and Pregabalin Treatment
2.3. vmDA Neuronal Viability and ATP Release Assessment
2.4. Immunocytochemistry
2.5. Morphogenetic Analysis
2.6. Quantitative PCR
2.7. Statistical Analysis
3. Results
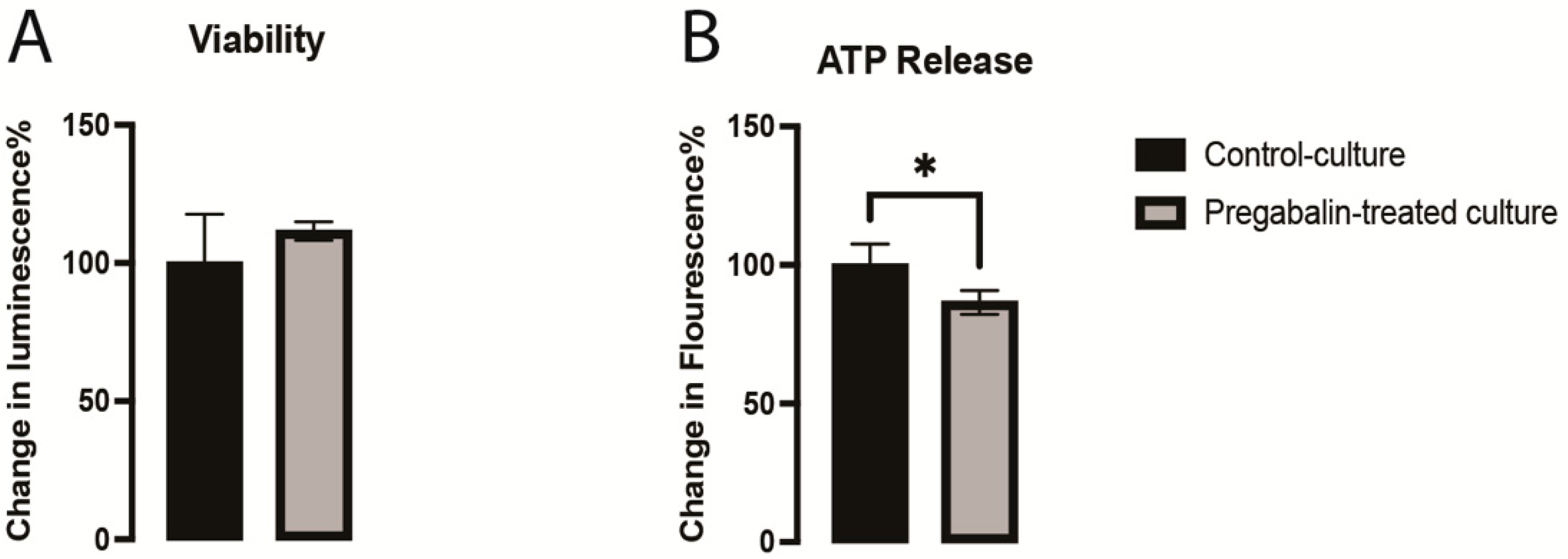
3.1. Pregabalin Disrupted Metabolic Activity of vmDA Neurons
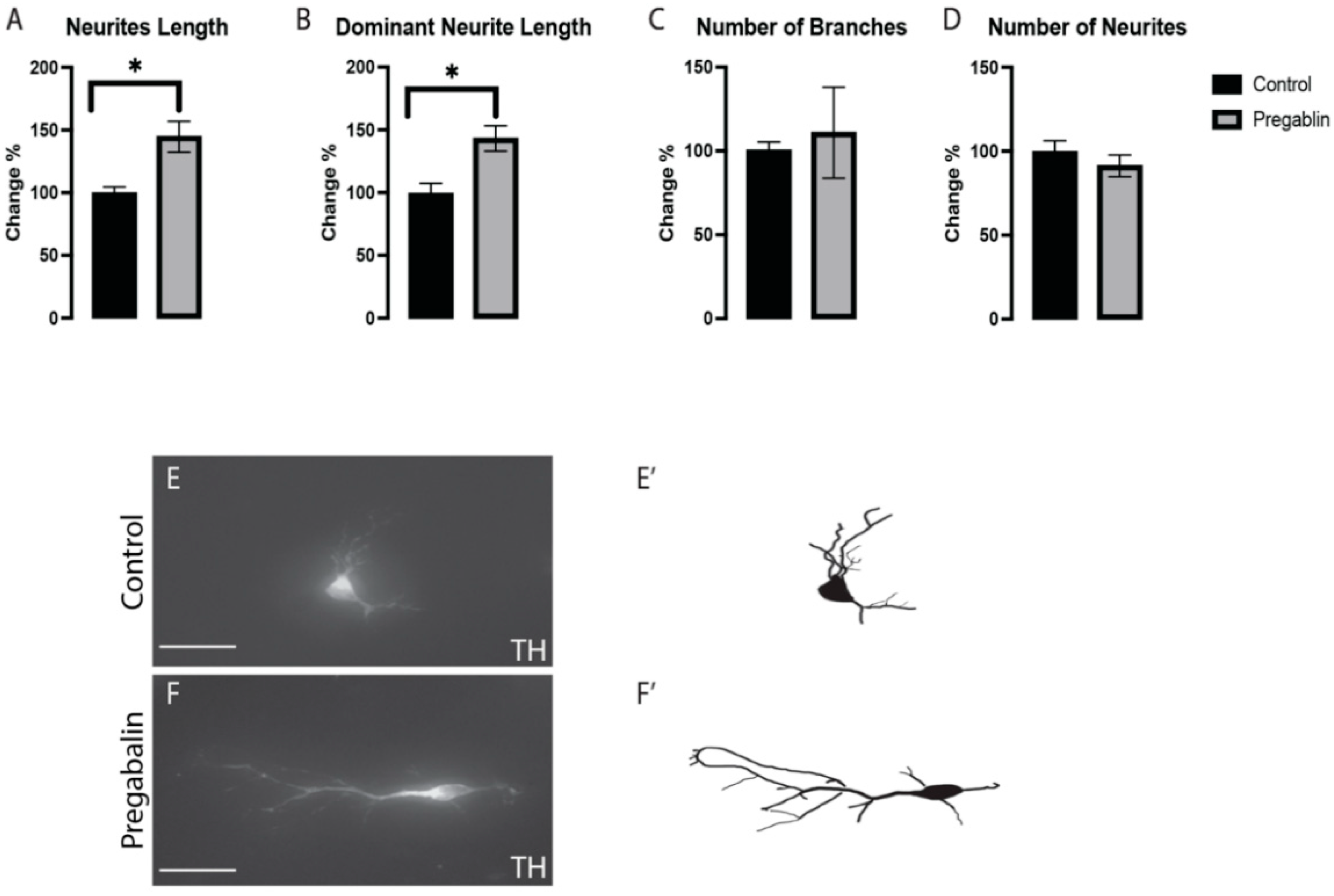
3.2. Pregabalin Affected the Morphogenesis of vmDA Neurons
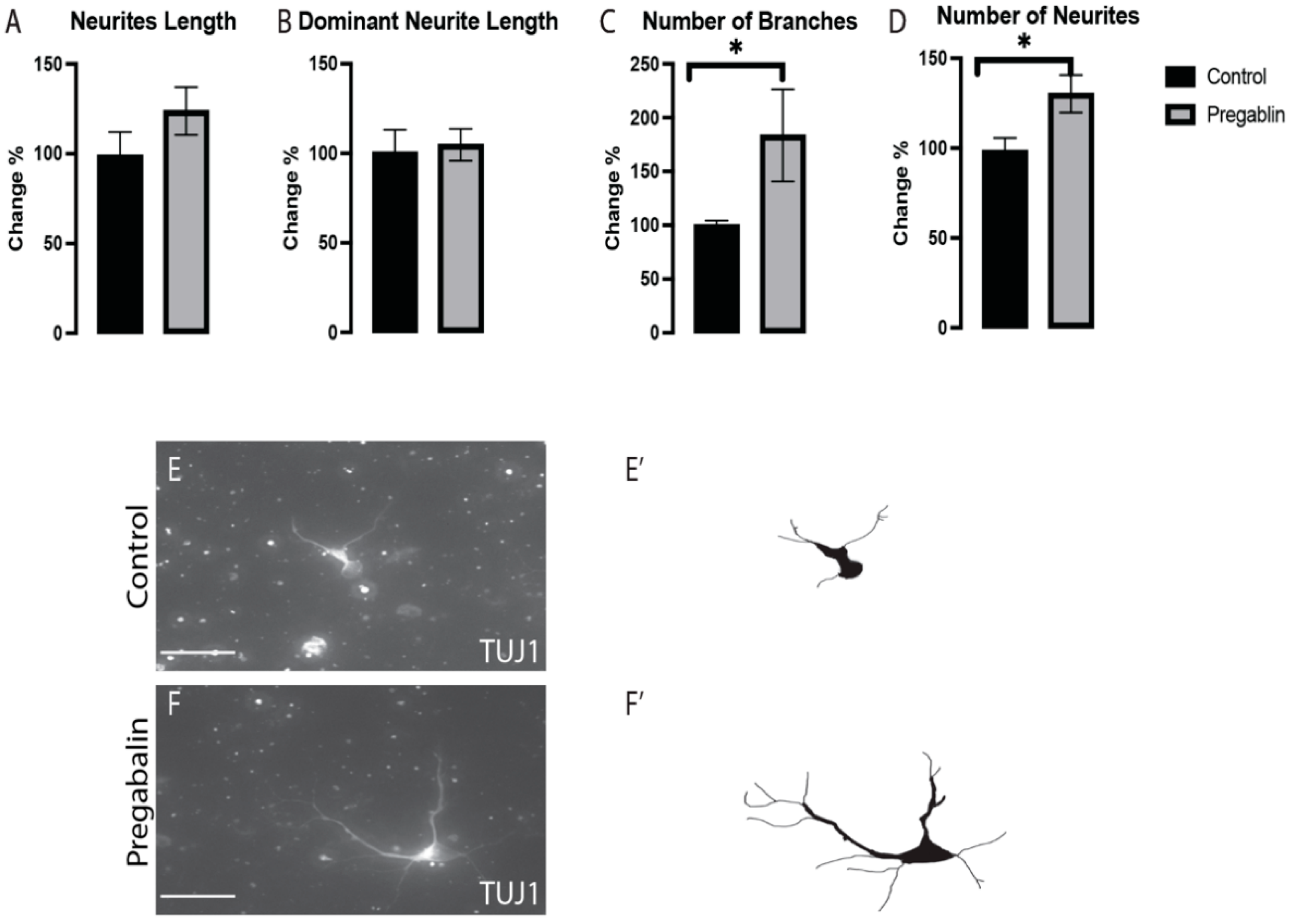
3.3. Pregabalin Affected the Morphogenesis of vm Non-DA Neurons
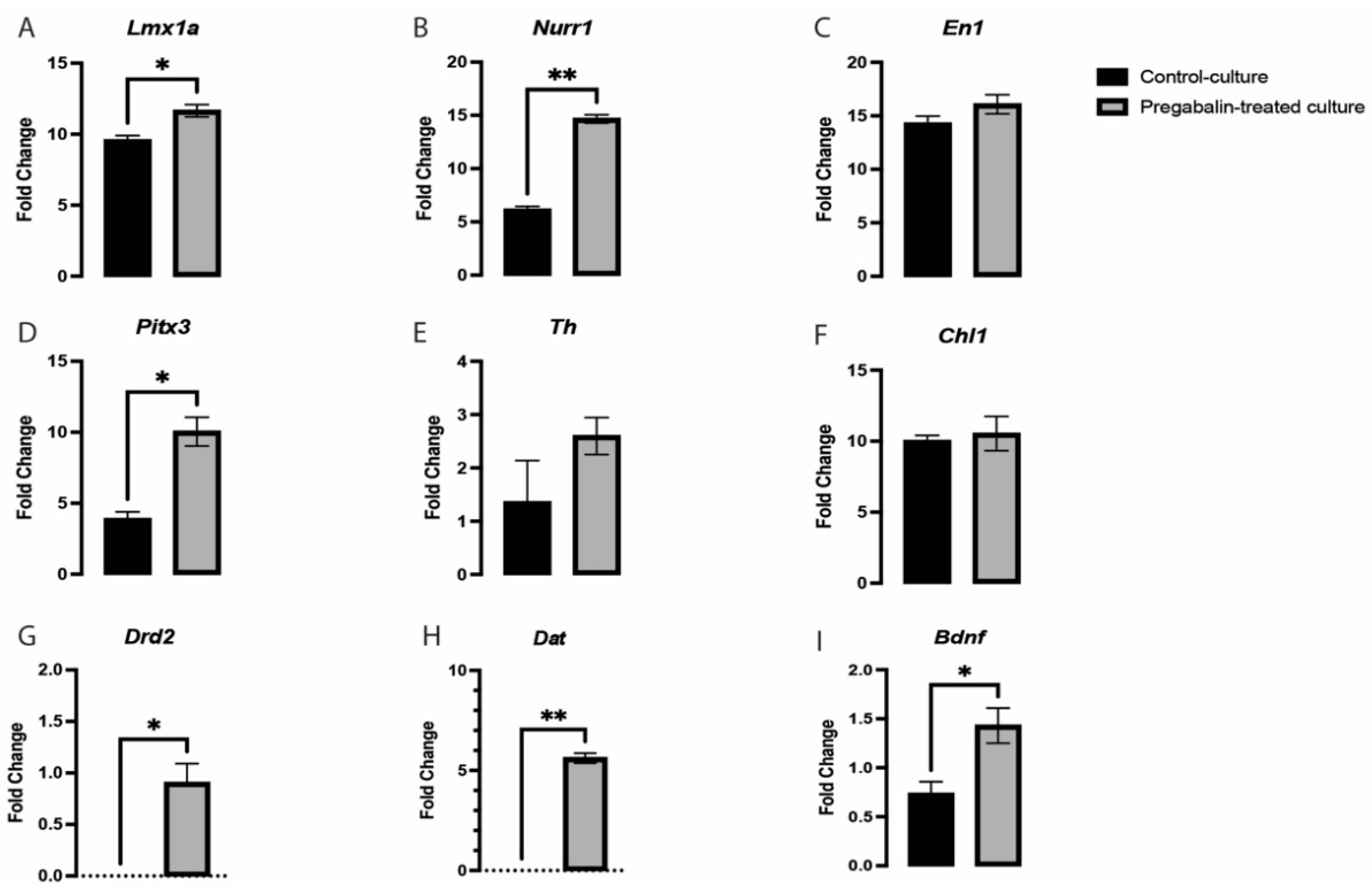
3.4. Pregabalin Induced the Upregulation of Key Dopaminergic-Related Genes in vmDA Neurons
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tassone, D.M.; Boyce, E.; Guyer, J.; Nuzum, D. Pregabalin: A novel γ-aminobutyric acid analogue in the treatment of neuropathic pain, partial-onset seizures, and anxiety disorders. Clin. Ther. 2007, 29, 26–48. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Singh, N.; Singh Jaggi, A. Send Orders for Reprints to [email protected] Pregabalin in Neuropathic Pain: Evidences and Possible Mechanisms. Curr. Neuropharmacol. 2014, 12, 44–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoptaw, S.J.; Kao, U.; Heinzerling, K.; Ling, W. Treatment for amphetamine withdrawal. Cochrane Database Syst. Rev. 2009, 2009, CD003021. [Google Scholar] [CrossRef] [PubMed]
- Shibahara, H.; Okubo, K.; Takeshita, N.; Nishimura, D. Medical treatment including pregabalin and radiation therapy provided remarkable relief for neuropathic pain by brachial plexus invasion in a patient with esophageal cancer. Gan Kagaku Ryoho 2012, 39, 277–280. [Google Scholar]
- Freynhagen, R.; Strojek, K.; Griesing, T.; Whalen, E.; Balkenohl, M. Efficacy of pregabalin in neuropathic pain evaluated in a 12-week, randomised, double-blind, multicentre, placebo-controlled trial of flexible- and fixed-dose regimens. Pain 2005, 115, 254–263. [Google Scholar] [CrossRef]
- Onakpoya, I.J.; Thomas, E.T.; Lee, J.; Goldacre, B.; Heneghan, C.J. Benefits and harms of pregabalin in the management of neuropathic pain: A rapid review and meta-analysis of randomised clinical trials. BMJ Open 2019, 9, e023600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, C.W.; Brett, A.S. Gabapentin and Pregabalin for Pain—Is Increased Prescribing a Cause for Concern? N. Engl. J. Med. 2017, 377, 411–414. [Google Scholar] [CrossRef]
- Spence, D. Bad medicine: Gabapentin and pregabalin. BMJ 2013, 347, f6747. [Google Scholar] [CrossRef] [PubMed]
- Patorno, E.; Bateman, B.T.; Huybrechts, K.F.; MacDonald, S.C.; Cohen, J.M.; Desai, R.J.; Panchaud, A.; Mogun, H.; Pennell, P.B.; Hernandez-Diaz, S. Pregabalin use early in pregnancy and the risk of major congenital malformations. Neurology 2017, 88, 2020–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Althobaiti, Y.S.; Almutairi, F.M.; Alshehri, F.S.; Altowairqi, E.; Marghalani, A.M.; Alghorabi, A.A.; Alsanie, W.F.; Gaber, A.; Alsaab, H.O.; Almalki, A.H.; et al. Involvement of the dopaminergic system in the reward-related behavior of pregabalin. Sci. Rep. 2021, 11, 10577. [Google Scholar] [CrossRef]
- Alsanie, W.F.; Bahri, O.A.; Habeeballah, H.H.; Alhomrani, M.; Almehmadi, M.; Alsharif, K.; Felemban, E.M.; Althobaiti, Y.S.; Almalki, A.H.; Alsaab, H.O.; et al. Generating homogenous cortical preplate and deep-layer neurons using a combination of 2D and 3D differentiation cultures. Sci. Rep. 2020, 10, 6272. [Google Scholar] [CrossRef]
- Alsanie, W.F.; Penna, V.; Schachner, M.; Thompson, L.; Parish, C.L. Homophilic binding of the neural cell adhesion molecule CHL1 regulates development of ventral midbrain dopaminergic pathways. Sci. Rep. 2017, 7, 9368. [Google Scholar] [CrossRef] [PubMed]
- Taha, S.H.N.; Zaghloul, H.S.; Ali, A.A.E.R.; Gaballah, I.F.; Rashed, L.A.; Aboulhoda, B.E. The neurotoxic effect of long-term use of high-dose Pregabalin and the role of alpha tocopherol in amelioration: Implication of MAPK signaling with oxidative stress and apoptosis. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 2020, 393, 1635–1648. [Google Scholar] [CrossRef]
- Elgazzar, F.M.; Elseady, W.S.; Hafez, A.S. Neurotoxic effects of pregabalin dependence on the brain frontal cortex in adult male albino rats. NeuroToxicology 2021, 83, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.M.E. Cognitive impairment and pregabalin dependence. Egypt. J. Psychiatry 2020, 41, 14. [Google Scholar] [CrossRef]
- Burbach, J.P.H.; Smits, S.; Smidt, M.P. Transcription Factors in the Development of Midbrain Dopamine Neurons. Ann. N. Y. Acad. Sci. 2006, 991, 61–68. [Google Scholar] [CrossRef]
- Heuvel, D.M.V.D.; Pasterkamp, R.J. Getting connected in the dopamine system. Prog. Neurobiol. 2008, 85, 75–93. [Google Scholar] [CrossRef]
- Parish, C.L.; Thompson, L.H. Modulating Wnt signaling to improve cell replacement therapy for Parkinson′s disease. J. Mol. Cell Biol. 2013, 6, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doucet-Beaupré, H.; Gilbert, C.; Profes, M.S.; Chabrat, A.; Pacelli, C.; Giguère, N.; Rioux, V.; Charest, J.; Deng, Q.; Laguna, A.; et al. Lmx1a and Lmx1b regulate mitochondrial functions and survival of adult midbrain dopaminergic neurons. Proc. Natl. Acad. Sci. USA 2016, 113, E4387–E4396. [Google Scholar] [CrossRef] [Green Version]
- Hobert, O.; Westphal, H. Functions of LIM-homeobox genes. Trends Genet. 2000, 16, 75–83. [Google Scholar] [CrossRef]
- Caiazzo, M.; Dell’Anno, M.T.; Dvoretskova, E.; Lazarevic, D.; Taverna, S.; Leo, D.; Sotnikova, T.D.; Menegon, A.; Roncaglia, P.; Colciago, G.; et al. Direct generation of functional dopaminergic neurons from mouse and human fibroblasts. Nature 2011, 476, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Bergman, O.; Håkansson, A.; Westberg, L.; Belin, A.C.; Sydow, O.; Olson, L.; Holmberg, B.; Fratiglioni, L.; Bäckman, L.; Eriksson, E.; et al. Do polymorphisms in transcription factors LMX1A and LMX1B influence the risk for Parkinson’s disease? J. Neural Transm. 2009, 116, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Doucet-Beaupré, H.; Lévesque, M. The role of developmental transcription factors in adult midbrain dopaminergic neurons. OA Neurosci. 2013, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Hoekstra, E.J.; Von Oerthel, L.; Van Der Heide, L.P.; Kouwenhoven, W.M.; Veenvliet, J.V.; Wever, I.; Jin, Y.-R.; Yoon, J.K.; Van Der Linden, A.J.A.; Holstege, F.C.P.; et al. Lmx1a Encodes a Rostral Set of Mesodiencephalic Dopaminergic Neurons Marked by the Wnt/B-Catenin Signaling Activator R-spondin 2. PLoS ONE 2013, 8, e74049. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.M.; Michelhaugh, S.K.; Bouhamdan, M.; Schmidt, C.J.; Bannon, M.J. The transcription factor NURR1 exerts concentration-dependent effects on target genes mediating distinct biological processes. Front. Neurosci. 2011, 5, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadkhodaei, B.; Alvarsson, A.; Schintu, N.; Ramsköld, D.; Volakakis, N.; Joodmardi, E.; Yoshitake, T.; Kehr, J.; Decressac, M.; Björklund, A.; et al. Transcription factor Nurr1 maintains fiber integrity and nuclear-encoded mitochondrial gene expression in dopamine neurons. Proc. Natl. Acad. Sci. USA 2013, 110, 2360–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torretta, S.; Rampino, A.; Basso, M.; Pergola, G.; Di Carlo, P.; Shin, J.H.; Kleinman, J.E.; Hyde, T.M.; Weinberger, D.R.; Masellis, R.; et al. NURR1 and ERR1 Modulate the Expression of Genes of a DRD2 Coexpression Network Enriched for Schizophrenia Risk. J. Neurosci. 2019, 40, 932–941. [Google Scholar] [CrossRef]
- Hegarty, S.V.; Sullivan, A.M.; O’Keeffe, G.W. Midbrain dopaminergic neurons: A review of the molecular circuitry that regulates their development. Dev. Biol. 2013, 379, 123–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.H.; Lim, S.H.; Shim, C.R.; Park, J.; Song, W.H.; Kwon, M.C.; Lee, J.H.; Park, J.S.; Choi, H.-G. Development of a Novel Controlled-Release Tablet of Pregabalin: Formulation Variation and Pharmacokinetics in Dogs and Humans. Drug Des. Dev. Ther. 2020, 14, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Jutzeler, C.R.; Cragg, J.J.; Warner, F.M.; Archibald, J.; Thomas, C.P.; Elliott, S.; Kramer, J.L. Journal Club: Pregnancy outcome following maternal exposure to pregabalin may call for concern. Neurology 2016, 88, e5–e7. [Google Scholar] [CrossRef] [Green Version]
- Wettermark, B.; Brandt, L.; Kieler, H.; Bodén, R. Pregabalin is increasingly prescribed for neuropathic pain, generalised anxiety disorder and epilepsy but many patients discontinue treatment. Int. J. Clin. Pract. 2014, 68, 104–110. [Google Scholar] [CrossRef]
- Derry, S.; Bell, R.F.; Straube, S.; Wiffen, P.J.; Aldington, D.; Moore, R.A. Pregabalin for neuropathic pain in adults. Cochrane Database Syst. Rev. 2019, 1, CD007076. [Google Scholar] [CrossRef]
- Ben-Menachem, E. Pregabalin Pharmacology and Its Relevance to Clinical Practice. Epilepsia 2004, 45, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ling, K.H.; Tan, J.J.; Lu, C.B. Development and Differentiation of Midbrain Dopaminergic Neuron: From Bench to Bedside. Cells 2020, 9, 1489. [Google Scholar] [CrossRef] [PubMed]
- Sałat, K.; Librowski, T.; Nawiesniak, B.; Gluch-Lutwin, M. Evaluation of analgesic, antioxidant, cytotoxic and metabolic effects of pregabalin for the use in neuropathic pain. Neurol. Res. 2013, 35, 948–958. [Google Scholar] [CrossRef]
- Margulis, A.V.; Hernandez-Diaz, S.; McElrath, T.; Rothman, K.J.; Plana, E.; Almqvist, C.; D’Onofrio, B.M.; Oberg, A.S. Relation of in-utero exposure to antiepileptic drugs to pregnancy duration and size at birth. PLoS ONE 2019, 14, e0214180. [Google Scholar] [CrossRef] [Green Version]
- Andrade, C. Safety of Pregabalin in Pregnancy. J. Clin. Psychiatry 2018, 79, 18f12568. [Google Scholar] [CrossRef]
- Yin, M.; Liu, S.; Yin, Y.; Li, S.; Li, Z.; Wu, X.; Zhang, B.; Ang, S.-L.; Ding, Y.-Q.; Zhou, J. Ventral Mesencephalon-Enriched Genes That Regulate the Development of Dopaminergic Neurons In Vivo. J. Neurosci. 2009, 29, 5170–5182. [Google Scholar] [CrossRef]
- Volpicelli, F.; De Gregorio, R.; Pulcrano, S.; Perrone-Capano, C.; di Porzio, U.; Bellenchi, G.C. Direct Regulation of Pitx3 Expression by Nurr1 in Culture and in Developing Mouse Midbrain. PLoS ONE 2012, 7, e30661. [Google Scholar] [CrossRef] [Green Version]
- Gyllborg, D.; Ahmed, M.; Toledo, E.M.; Theofilopoulos, S.; Yang, S.; Ffrench-Constant, C.; Arenas, E. The Matricellular Protein R-Spondin 2 Promotes Midbrain Dopaminergic Neurogenesis and Differentiation. Stem Cell Rep. 2018, 11, 651–664. [Google Scholar] [CrossRef] [Green Version]
- Hyman, C.; Hofer, M.; Barde, Y.; Juhasz, M.; Yancopoulos, G.D.; Squinto, S.P.; Lindsay, R.M. BDNF is a neurotrophic factor for dopaminergic neurons of the substantia nigra. Nature 1991, 350, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.-Y.; Hong, S.; Jeong, J.-W.; Choi, S.; Kim, H.; Kim, J.; Kim, K.-S. Vesicular monoamine transporter 2 and dopamine transporter are molecular targets of Pitx3 in the ventral midbrain dopamine neurons. J. Neurochem. 2009, 111, 1202–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Choi, K.C.; Chang, M.S.; Kim, M.H.; Na, Y.-S.; Lee, J.E.; Jin, B.K.; Lee, B.-H.; Baik, J.-H. The Dopamine D2 Receptor Regulates the Development of Dopaminergic Neurons via Extracellular Signal-Regulated Kinase and Nurr1 Activation. J. Neurosci. 2006, 26, 4567–4576. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.J. Critical issues in BDNF Val66met genetic studies of neuropsychiatric disorders. Front. Mol. Neurosci. 2018, 11, 156. [Google Scholar] [CrossRef] [Green Version]
- Hansen, F.H.; Skjørringe, T.; Yasmeen, S.; Arends, N.V.; Sahai, M.; Erreger, K.; Andreassen, T.F.; Holy, M.; Hamilton, P.; Neergheen, V.; et al. Missense dopamine transporter mutations associate with adult parkinsonism and ADHD. J. Clin. Investig. 2014, 124, 3107–3120. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zhang, J. The association of DRD2 with insight problem solving. Front. Psychol. 2016, 7, 1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′ to 3′) |
|---|---|
| GAPDH | F-primer: TGAAGGTCGGAGTCAACGGA R-primer: CCAATTGATGACAAGCTTCCCG |
| Th | F-primer: TGAAGGAACGGACTGGCTTC R-primer: GAGTGCATAGGTGAGGAGGC |
| Nurr1 | F-primer: GACCAGGACCTGCTTTTTGA R-primer: ACCCCATTGCAAAAGATGAG |
| Lmx1a | F-primer: GAGACCACCTGCTTCTACCG R-primer: GCACGCATGACAAACTCATT |
| En1 | F-primer: TCACAGCAACCCCTAGTGTG R-primer: CGCTTGTCTTCCTTCTCGTT |
| Pitx3 | F-primer: CATGGAGTTTGGGCTGCTTG R-primer: CCTTCTCCGAGTCACTGTGC |
| Chl1 | F-primer: TGGAATTGCCATTATGTGGA R-primer: CACCTGCACGTATGACTGCT |
| Dat | F-primer: TTGCAGCTGGCACATCTATC R-primer: ATGCTGACCACGACCACATA |
| Drd2 | F-primer: CTCAACAACACAGACCAGAAT R-primer: GAACGAGACGATGGAGGA |
| Bdnf | F-primer: ACTATGGTTATTTCATACTTCGGTT R-primer: CCATTCACGCTCTCCAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsanie, W.F.; Alhomrani, M.; Gaber, A.; Habeeballah, H.; Alkhatabi, H.A.; Felimban, R.I.; Abdelrahman, S.; Hauser, C.A.E.; Chaudhary, A.G.; Alamri, A.S.; et al. The Effects of Prenatal Exposure to Pregabalin on the Development of Ventral Midbrain Dopaminergic Neurons. Cells 2022, 11, 852. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11050852
Alsanie WF, Alhomrani M, Gaber A, Habeeballah H, Alkhatabi HA, Felimban RI, Abdelrahman S, Hauser CAE, Chaudhary AG, Alamri AS, et al. The Effects of Prenatal Exposure to Pregabalin on the Development of Ventral Midbrain Dopaminergic Neurons. Cells. 2022; 11(5):852. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11050852
Chicago/Turabian StyleAlsanie, Walaa F., Majid Alhomrani, Ahmed Gaber, Hamza Habeeballah, Heba A. Alkhatabi, Raed I. Felimban, Sherin Abdelrahman, Charlotte A. E. Hauser, Adeel G. Chaudhary, Abdulhakeem S. Alamri, and et al. 2022. "The Effects of Prenatal Exposure to Pregabalin on the Development of Ventral Midbrain Dopaminergic Neurons" Cells 11, no. 5: 852. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11050852