
Revealing the Roles of MOAP1 in Diseases: A Review


Department of Neurobiology, School of Basic Medical Sciences, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, China
*
Author to whom correspondence should be addressed.
Cells 2022, 11(5), 889; https://0-doi-org.brum.beds.ac.uk/10.3390/cells11050889
Submission received: 9 December 2021
/
Revised: 28 February 2022
/
Accepted: 1 March 2022
/
Published: 4 March 2022
(This article belongs to the Collection Rejuvenating, Geroprotective and Cytoprotective Activities of Natural and Synthetic Compounds: Proofs of Concept, Associated Mechanisms and Applications)
Abstract
:Modulator of apoptosis protein1 (MOAP1), also known as MAP1 and PNMA4, belongs to the PNMA gene family consisting of at least 15 genes located on different chromosomes. MOAP1 interacts with the BAX protein, one of the most important apoptosis regulators. Due to its critical role in a few of disease-associated pathways, MOAP1 is associated with many diseases such as cancers and neurological diseases. In this study, we introduced MOAP1 and its biological functions and reviewed the associations between MOAP1 and a few diseases including cancers, neurological diseases, and other diseases such as inflammation and heart diseases. We also explained possible biological mechanisms underlying the associations between MOAP1 and these diseases, and discussed a few future directions regarding MOAP1, especially its potential roles in neurodegenerative disorders. In summary, MOAP1 plays a critical role in the development and progression of cancers and neurological diseases by regulating a few genes related to cellular apoptosis such as BAX and RASSF1A and interacting with disease-associated miRNAs, including miR-25 and miR1228.
1. Introduction
The modulator of apoptosis protein1 (MOAP1), also known as MAP1 and PNMA4, belongs to the PNMA gene family, which consists of at least 15 genes on different chromosomes [1]. Specifically, PNMA1 and PNMA4 are located in human chromosome 14; PNMA8A, PNMA8B, PNMA8C, and CCDC8 (a paralog of PNMA6F) are localized to chromosome 19; PNMA2 belongs to chromosome 8; and PNMA3, PNMA5, PNMA6A, PNMA6B, PNMA6E, PNMA6F, PNMA7A, and PNMA7B are located in X chromosome [1]. It is known that the PNMA family is highly expressed in the brain, testis, and heart with MOAP1, a resident protein in the mitochondria [1]. There are multiple acronyms existing for MOAP1 (e.g., MAP1, PNMA4, etc.), and we unified them to MOAP1 from now on to avoid any confusion.
Among the PNMA genes, MOAP1 is probably the most important because it not only plays a central role in mitochondria and death receptor-mediated apoptosis [2], but also is associated with many diseases like cancer [3]. The specific roles of MOAP1 in cellular apoptosis and various diseases are determined by its protein structure. Specifically, in the BH3L domain of MOAP1, a unique protein sequence or structure (UPD) between the NCD and CCD domains was identified. The protein sequence of MOAP1 also contains a domain rich in lysine and arginine residues, named KRs [1].
First identified by the yeast-two-hybridization assay, MOAP1 was a BAX-associated protein that binds to BAX only during apoptosis induction [2]. It plays a vital role in mitochondrial-dependent apoptosis by influencing and maintaining BAX activation. The down-regulation of MOAP1 leads to the instability and polymerization of BAX on the mitochondrial membrane so that cytochrome C is not released into the cytoplasm. The knockdown of this protein inhibited the apoptotic stimulation induced by TNFα and TRAIL. Therefore, abolishing this protein will cause cells to be insensitive to death signals mediated by the external and internal pathways of apoptosis [4].
In addition, the complement system is a tightly regulated network of proteins involved in defense against pathogens, inflammatory processes, and the coordination of innate and adaptive immune responses. Hertz et al. mentioned MOAP1 as a pattern-recognition molecule (PRM) related to inhibitors of the complement system’s lectin pathway. The C4B binding protein (C4BP) regulates both classical and lectin pathways and MOAP1/C4BP chimeras have unique complement inhibition properties, providing a new therapeutic approach for upstream and central complement activation [4]. MOAP1 also plays a role in liver Fas signaling by promoting MTCH2-mediated tBid recruitment to mitochondria [5].
Besides BAX, MOAP1 also attenuates cell apoptosis by collaborating with other molecules. For example, it is known that RASSF1A and MOAP1 intimately interact to form the RASSF1A/MOAP1 molecular pathway in cancer. One potential function of this pathway is to link death receptors with BAX conformational change to promote tumor suppression [6]. MOAP1 also interacts with RASSF6 to attenuate RASSF6-induced cell apoptosis; however, the attenuation was not observed in a p53-negative background [7,8,9].
Moreover, MOAP1 is regulated by ubiquitin protein degradation [10]. Ubiquitin proteasome system is an indispensable process during the transformation from normal cells to cancer cells. There are a few studies showing that a new regulation factor, Trim39, can stabilize MOAP1 [11,12]. Moreover, UBR5 enhances MOAP1 ubiquitination. Matsuura et al. found that UBR5 ubiquitin ligase down-regulates MOAP1 in ovarian cancer, which contributes to its role in ovarian cancer resistance to cisplatin [13]. Since the in-vitro MOAP1 expression level is low in tumor cells, MOAP1 ubiquitin can be reduced by apoptosis stimulating processes [13].
Finally, there are a few studies suggesting a unique role of MOAP1 in promoting autophagy signal transduction. Autophagy goes through three different stages by phagocytotic molecules and organelles, including initiation of recruitment, nucleation, and expansion of phagocytic membrane, followed by lysosomal degradation. P62 is an autophagy receptor. The BAX-binding protein MOAP1 regulates the p62-KEAP1-NRF2 signaling pathway by disrupting the p62 corpuscles. MOAP1 is recruited to p62 corpuscles by the induction of formation stimulated by cellular stress and reducing their levels independently of the autophagy pathway [14]. Interestingly, MOAP1 was found to be an LC3-binding protein, which plays a unique role in promoting autophagy by interacting with LC3 to promote efficient shutdown of autophagy during starvation [15].
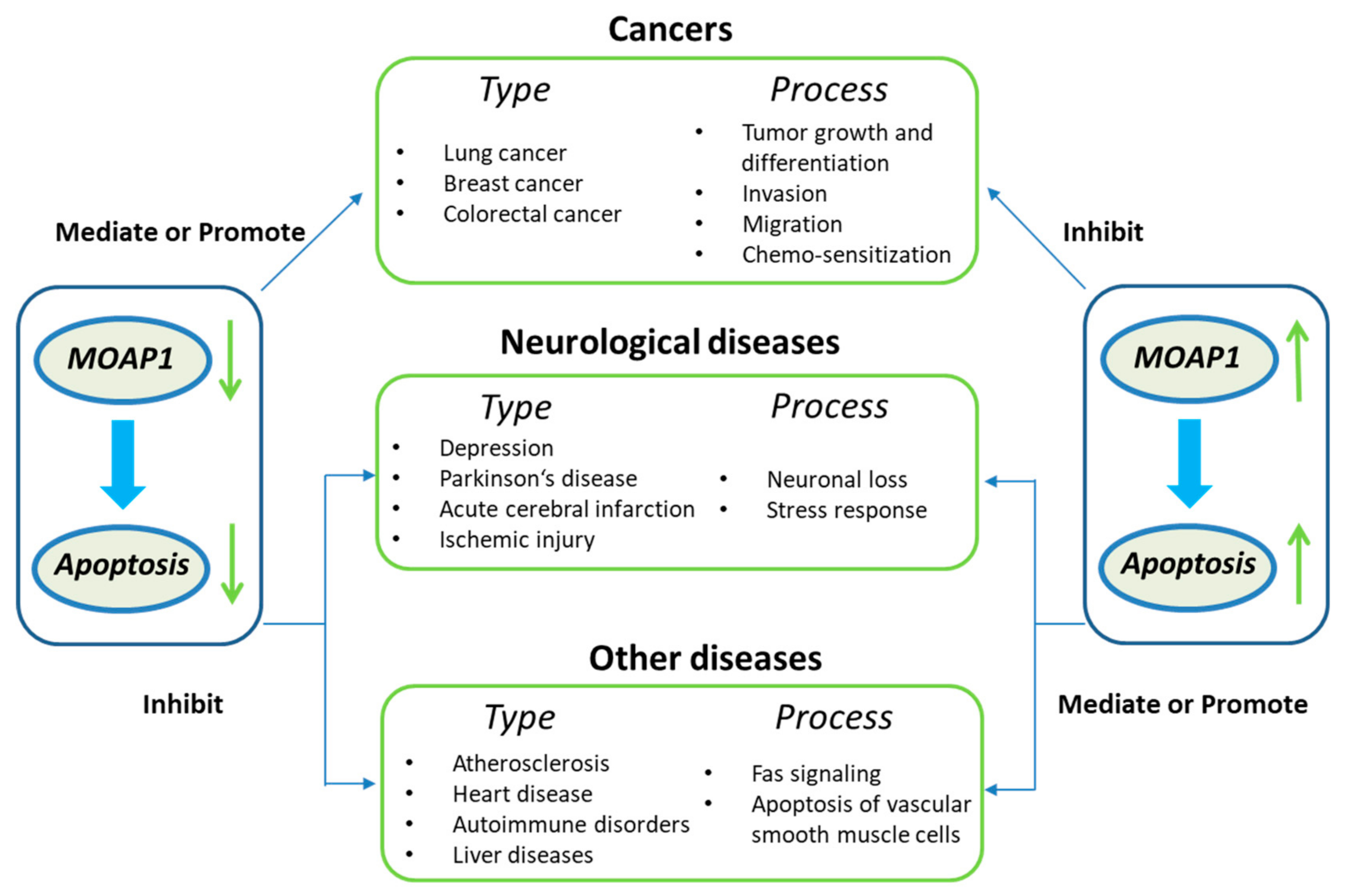
In summary, MOAP1 can activate the apoptosis signaling pathway, promoting its role in many diseases, such as cancers and neurological diseases (Figure 1). For example, MOAP1 is involved in the progression, invasion, metastasis, and chemo-sensitization of various cancers, including lung cancer, colorectal cancer, and breast cancer [16,17,18]. In the nervous system, MOAP1 can induce neuronal loss, actuate cerebral infarction, depressive behavior, and stress response [19,20]. Finally, Fas, also known as Cd95 and APO1, is involved in diseases like atherosclerosis, heart disease, autoimmune disorders, liver disease, and myocardial infarction [21,22,23,24,25]. Since there is evidence that MOAP1 and Fas signaling are highly associated [5], MOAP1 may also play a role in these diseases. In the following, we will review the molecules mediating the association between MOAP1 and the diseases mentioned above and their acting mechanisms.
2. Roles of MOAP1 in Cancers
With the increasing of human lifespan, cancer has become one of the top three diseases threatening human lives [26]. The hallmarks of cancer, including evading apoptosis, self-sufficiency in growth signals, insensitivity to anti-growth signals, tissue invasion and metastasis, limitless replicative potential, and sustained angiogenesis; the five processes are mutually interacted [27]. Previous studies have suggested that MOAP1 is highly associated with the apoptosis pathway [28], one of the most critical pathways and hallmarks in cancer. Thus, the roles of MOAP1 in various cancers and related biological mechanisms have been widely studied [16,29], among which lung cancer, colorectal cancer (CRC), and breast cancer are probably the most well studied.
2.1. MOAP1 in Lung Cancer
Lung cancer is the most prevalent cancer in China and the third most common cancer in the United States (https://www.cdc.gov/cancer/lung/ accessed on 9 December 2021), accounting for approximately 13% of new cancer incidences worldwide and causing 1.4 million deaths each year [30]. Lung cancer consists of small cell lung cancer (SCLC) and non-small cell lung cancer (NSCLC), which can be further divided into squamous-cell carcinoma, adenocarcinoma, large-cell carcinoma, and so on.
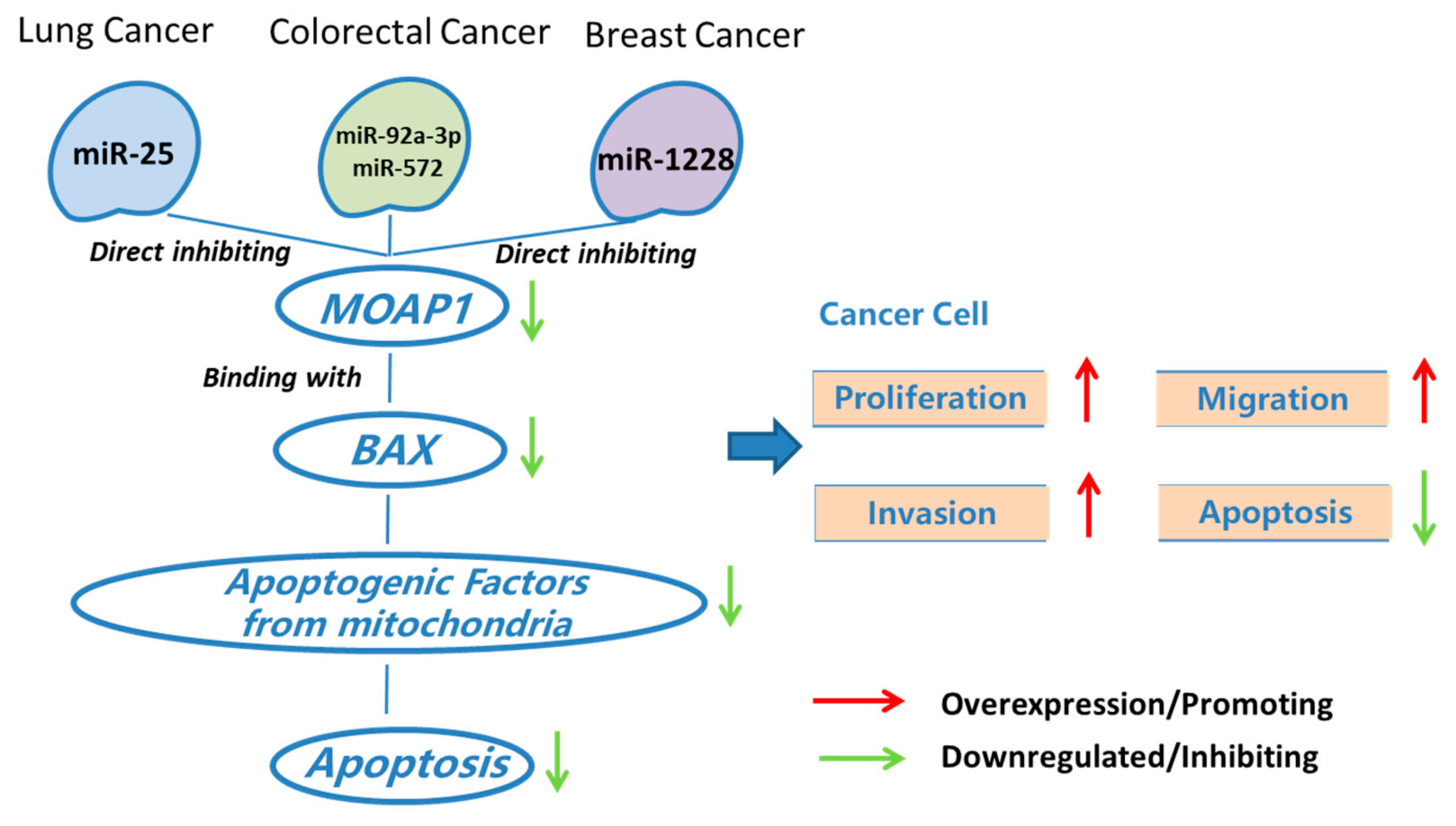
The proteins in the BCL2 family can regulate the intrinsic apoptotic pathway [31]. BCL2 family proteins consist of pro-apoptotic proteins (like BAX, BAK, and BAD) and anti-apoptotic proteins (like BCL2, BCLW, and MCL1). Liu et al. observed abnormal expression of BCL2 protein in NSCLC and showed that BCL2 and MCL1 amplification is related to drug resistance [32]. Chan et al. found that MOAP1 can directly or indirectly interact with BCL2 family proteins, which promotes its role in lung cancer [15] (Figure 2).
On the other way, the role of miRNAs in regulating the apoptosis pathway in lung cancer has been widely studied. For example, some miRNAs are critical in lung cancer treatment by targeting TP53-dependent cell apoptosis [33]. Previous studies also suggested that miRNAs regulating lung cancer cell apoptosis interact with many important cancer pathways, including PI3K/Akt, PTEN, GSK-3b, NF-kB, BCL2, BAX, P53, and mTOR [34]. However, the biological mechanisms underlying lung cancer-associated miRNAs are unknown. MOAP1 is one of the genes to mediate lung cancer-related miRNAs and lung cancer. That is, a few miRNAs can regulate MOAP1 and BAX, which may contribute to the apoptosis of lung cancer cells. For example, miR-25 is a critical biomarker for non-small-cell lung cancer. Wu et al. found that this miRNA down-regulates the expression of MOAP1, which promotes cell proliferation and inhibits caspase-dependent apoptosis in NSCLC cells (Figure 2). Thus, miR-25 antagomir can promote MOAP1 expression and thus inhibit lung cancer growth in a mouse xenograft model [16]. We refer to synthetic antagonists as silencing agents of miRNAs by the term antagomir.
2.2. MOAP1 in Colorectal Cancer
Colorectal cancer (CRC), also known as bowel cancer, is among the top three most common cancers in the world. This cancer is caused by uncontrolled cell growth in the colon or rectum; the tumors in the two body parts are genetically the same and thus are called uniformly as colorectal cancer (https://0-www-nature-com.brum.beds.ac.uk/subjects/colorectal-cancer accessed on 9 December 2021). One major issue in CRC and probably all cancers is recurrence and metastasis, which eventually leads to the death of most cancer patients. That is, the cancer cell re-grows in the same location after surgical resection or move to another body location through blood flow and proliferation [35].
In recent years, much effort has been placed on revealing the biological mechanisms behind the progression, recurrence, and metastasis of CRC. Among many pathways related to CRC, p53-dependent apoptosis is probably the most studied. BAX and BCL2 are two important regulators of p53-dependent apoptosis and thus are critical in many processes of CRC. For example, Katkoori et al. suggested that the expression level of BAX in a CRC patient can be used to predict his/her prognosis [36]. Low BAX expression was considered a negative prognostic factor for CRC patients with liver metastasis in an early study [37]. The BAX/BCL2 ratio was also considered one of the prognostic markers for tumor location in CRC [38]. Besides the prognostic value, BAX and BCL2 are also important for CRC at molecular levels. Pryczynicz et al. found that BAX protein could reduce CRC invasion [38]. Jansson and Sun found that the expression of BAX was significantly higher in metastatic CRC than in primary CRC [39]. In addition, BAX is also involved in other inhibition mechanisms associated with CRC, such as the PGC1α pro-apoptotic effect [40], regulating autophagy-related genes [41], and mediating CRC with microsatellite instability [42]. Similarly, BCL2 is a suppressor of p53-dependent apoptosis in CRC [43] and is found to regulate the invasion and metastasis of CRC cells [44].
It is known that BAX could be regulated by MOAP1 in CRC [3,6], which promotes the roles of MOAP1 in various processes of CRC (Figure 2). Specifically, the function of BAX and MOAP1 might be regulated by a few miRNAs. For instance, miR-92a-3p and miR-572 are two known colorectal cancer-associated miRNAs. High miR-92a-3p expression contributes to chemotherapy resistance and metastasis of CRC [17]. A recent study suggested that these functions of miR-92a-3p were achieved by inhibiting FBXW7 and MOAP1 directly to activate the Wnt/β-catenin pathway and inhibit cell apoptosis, respectively [17] (Figure 2). In addition, the up-regulation of miR-572 can inhibit MOAP1 expression, which promotes the migration, invasion, and growth of CRC cells [29] (Figure 2). As a result, miR-572 antagomir might be a potential therapeutic means for treating colorectal cancer.
2.3. MOAP1 in Breast Cancer
Breast cancer is the second most common woman cancer in the United States (https://www.cdc.gov/cancer/breast/statistics/ accessed on 9 December 2021). There are five main molecular subtypes of breast cancer: Luminal A, Luminal B, Triple-negative/basal-like, HER2-enriched, and Normal-like. HER2 is probably the most important gene for breast cancer, with HER2-positive and HER2-negative patients accounting for about 25–30% and 70–75% of all breast cancer incidences, respectively [45,46,47]. Even after trastuzumab adjuvant treatment, a large portion of breast cancer develops recurrence and metastasis, especially for the HER2-positive subtype.
The role of BAX apoptosis signaling in breast cancer has been widely studied. BAX and BCL2 are known prognostic biomarkers for breast cancer. BCL2, also known as BCL2 Apoptosis Regulator and B-cell lymphoma-2, is a gene capable of suppressing apoptosis. Specifically, the integral outer mitochondrial membrane protein encoded by BCL2 can block the apoptotic death of many cells like lymphocytes, making it a prognostic biomarker for cancers. Sjostrom et al. identified the predictive value of BAX and BCL2 in chemotherapy responses in advanced breast cancer [48]. Azimian et al. found that the ratio between the expressions of BAX and BCL2 (BAX/BCL2) could be used to predict the effect of breast cancer radiotherapy in an individual patient [49]. In addition, Binder et al. found that the differential BAX/BCL2 expression pattern is important for cancer cell proliferation and thus is a predictive marker for cancer malignant progression [50]. Linjawi et al. identified that BCL2 expression is significantly correlated with hormonal receptors in breast cancer patients. Sehrawat suggested that BAX activation can mediate the selective death of human breast cancer cells and might provide some direction for breast cancer treatment. Finally, a recent study suggested that the tricistronic expression of MOAP1, BAX, and RASSF1A enhances chemo-sensitization in breast cancer cell lines [51].
As a result, MOAP1, its upstream genes, and miRNAs play critical roles in the progression, invasion, and migration of breast cancer. For example, miR-1228 is a miRNA responsible for breast cancer invasion and migration. Lin et al. found that miR-1228 could be attenuated by increasing the expression of MOAP1 (Figure 2), suggesting the potential of MOAP1 in breast cancer studies and therapies.
Together, the downregulation of MOAP1 could reduce the effect of BAX, which may suppress p53-dependent cancer cell apoptosis. This process places an important role in bridging various cancers and their associated miRNAs. Overexpression of these miRNAs usually inhibits the expression of MOAP1, thus promoting cancer cell proliferation, migration, and invasion.
2.4. MOAP1 in Cancer Drug Resistance and Chemo-Sensitization
MOAP1 was involved in cancer drug resistance and chemo-sensitization (Figure 3). For example, Matsuura et al. found that UBR5 down-regulates proapoptotic MOAP1 in ovarian cancer, which results in cisplatin resistance [13] (Figure 3). In cancer cells, tricistronic expression of MOAP1, BAX, and RASSF1A (MBR) expression will induce cell death and thus increase chemo-sensitization. Recently, Lee et al. found that this process requires the involvement of the BH3L domain of MOAP1 [51]. As a member of the PNMA family, MOAP1 also interacts with other members in this family to promote chemo-sensitization. In MCF-7 cells, overexpression of MOAP1 and PNMA1 will promote chemo-sensitization; however, the chemo-sensitization will be greatly inhibited when co-expressing with PNMA2, indicating that PNMA2 might be an antagonist of MOAP1 and PNMA1 induced chemo-sensitization [52]. In addition, the co-expression of PNMA5 and MOAP1 greatly enhances the chemo-sensitivity of MCF-7 cells to Etoposide treatment [52].
3. Roles of MOAP1 in Neurological Diseases
Previous studies suggested that PNMA2, PNMA3, and MOAP1 mRNAs are abundantly expressed in mouse brains. In the neocortex, PNMA2, PNMA3, and MOAP1 mRNAs are expressed throughout all layers, although the expression levels of the genes differ slightly among the layers [19]. It is known that neurological diseases, especially neuron-degenerative diseases such as Alzheimer’s disease and Parkinson’s disease, are highly associated with mitochondrial-dependent neuron cell apoptosis [53,54]. In addition, MOAP1 is abundantly expressed in other brain tissues [20,55]. MOAP1 is speculated to be associated with neurological diseases, and there are a few studies in this area.
3.1. MOAP1 and Cerebral Ischemia
Stroke remains one of the top diseases for human disability and death worldwide. This is largely due to the narrow time window for recognition and the administration of outcome-modifying treatment [56]. Acute stroke can result in permanent damage to the human brain and even cognitive death, making it critical to unveil the mechanism behind its formation and treatments.
A few early studies suggested that upregulation of BAX proteins in neuronal cells is a risk factor for cerebral ischemia [57,58]. BAX ablation might be a feasible direction to control cerebral ischemia [59], which makes MOAP1 critical in cerebral ischemia since it can regulate BAX and BCL2 in neurons. Studies have shown that the MOAP1−/− primary cortical neurons and MOAP1þ/þ primary neurons are resistant against oxygen and glucose deprivation (OGD) treatment; MOAP1−/− primary cortical neurons are stronger. In the mouse transient middle cerebral artery occlusion (tMCAO) model, cerebral ischemia causes the MOAP1/BAX association, activating the MOAP1-dependent apoptosis cascade. In the study, the authors found that 24 h after tMCAO, MOAP1−/− mice showed less neuron loss and smaller infarct volume than MOAP1+/+ mice. Similarly, MOAP1−/− mice also have better integrity in neurological functions, which was proved in their rotarod test [20]. Together, these studies suggested that MOAP1 plays an important role in regulating apoptosis in cerebral ischemic injury.
3.2. MOAP1 and Depression
Stress response is closely related to one of the most important mental diseases, namely depression. Depression, also called major depressive disorder, is a serious mental illness that negatively affects people’s the feelings and actions.
Depression and BAX apoptosis signaling are highly correlated. For example, the signal cascade composed of BAD, BAX, and Caspase 3 is sufficient for synaptic depression [60]. In addition, the upregulation of BAX and downregulation of BCL2 were observed in the olfactory bulb of a rat depression model [61]. Due to its regulatory role in BAX apoptosis, MOAP1 plays an important role in regulating stress response by the Dorsal raphe nucleus (DRN). Forced swimming test is one of the famous techniques in wet lab to develop depression model in mice [62]. Compared with age-matched wild-type mice, young MOAP1−/− mice in the forced swimming test showed depressive behavior with increased stationary time, which was eliminated by acute fluoxetine treatment. In young wild-type (WT) control mice, repeated forced swimming stress resulted in upregulation of tryptophan hydroxylase 2 (TPH2) and down-regulation of brain-derived neurotrophic factor (BDNF) in the dorsal renal nucleus (DRN). In contrast, TPH2 was not upregulated in aging WT mice. Interestingly, this stress response was not present in both young and old MOAP1−/− mice [55].
3.3. MOAP1 and Parkinson’s Disease
Parkinson’s disease (PD) becomes the second most common neurodegenerative disease in the world after Alzheimer’s disease, affecting nearly five million people worldwide, and this number is expected to double by 2030 [63]. In Parkinson’s disease, the main pathological features are connected with progressive neurodegeneration of dopaminergic neurons and typical motor characteristics in the substantia nigra striatum (SNC), which locates in the dense region of the black substantia [64]. It results from genetic, environmental, and aging factors and currently has no good treatment. Apoptosis and autophagy play a vital function in the pathogenesis of PD. Therefore, adjusting their balance is a potential therapeutic strategy [63]. Previous studies have suggested that the BCL2 protein family is a therapeutic biomarker in Parkinson’s disease [53,65,66,67]. Specifically, proteins in the BCL2 family, such as BAX, can mediate the apoptosis of dopaminergic neurons in the Parkinson’s disease. For example, Vila et al. found that the ablation of BAX protein can prevent neurodegeneration in Parkinson’s disease [68]. By targeting BAX, some miRNAs like MicroRNA-216a also inhibit neuronal cell apoptosis in Parkinson’s disease [69]. Thus, proteins in the BCL2 family might be a promising target in treating Parkinson’s disease.
It is well known that MOAP1 directly binds to BAX, which suggests that MOAP1 may play a role in Parkinson’s disease. Capurro et al. used external laser capture micro-anatomical data to verify the differentially expressed genes identified by PSEA (population-specific expression analysis) in the dense part of substantia nigra neurons. In this study, the expression of the MOAP1 gene was found to be reduced in cortical neurons of Parkinson’s disease (PD) [70]. In Table 1, we summarized the genes involved in the connection between MOAP1 and neuron-related traits and diseases. Besides BAX, MOAP1 also interacts with other molecules like PNMA2. For example, in immunoprecipitation studies, PNMA2 was closely related to PNMA1 and MOAP1 and functionally antagonized the pro-apoptotic of MOAP1 and PNMA1 [71].
Besides cancers and neuronal diseases, MOAP1 was also related to a few other diseases. For example, MOAP1 was targeted by miR-25 to block the apoptosis of vascular smooth muscle cells, which plays a critical role in inflammation and heart disease [21]. MOAP1 is also commonly identified differential genes of coronary artery disease, ischemic cardiomyopathy, and myocardial infarction [73,74]. In addition, it is known that Fas, also known as Cd95 and Apo1, is implicated in a few diseases like liver diseases, autoimmune disorders, and fulminant hepatitis [22,23,24,25]. Tan et al. found that MOAP1 is involved in the Fas signaling for apoptosis in the liver [5] and thus is associated with liver diseases. In summary, MOAP1 might be implicated in many diseases and its acting mechanisms are yet to be discovered.
4. Conclusions and Future Perspectives on MOAP1
In this study, we reviewed the roles and mechanisms of MOAP1 in a few biological processes including cell apoptosis, innate and adaptive immunity, ubiquitin protein degradation, and autophagy. We also reviewed the molecules mediating the association between MOAP1 and a few diseases like cancers and neurological diseases.
As an important gene in the PNMA gene family, MOAP1 can bind with the BAX protein to activate mitochondria and death receptor-mediated cell apoptosis, which will inhibit the progression and metastasis of a few cancers like lung cancer, breast cancer, and colorectal cancer and may mediate neurological diseases. In addition, a few miRNAs can directly interact with MOAP1 in a few cancers. For example, miR-25 can directly inhibit MOAP1 in lung cancer; miR-92a-3p and miR-572 can inhibit MOAP1 in colorectal cancer, and miR-1228 can inhibit MOAP1 in breast cancer. These may explain the functions of these miRNAs in cancer etiology. Moreover, MOAP1 also interacts with other genes to perform specific functions. For example, MOAP1 and RASSF1A collaborate to form the RASSF1A/MOAP1 pathway, which is important for cell apoptosis and chemosensitivity. MOAP1 also interacts with RASSF6 and MBR to promote cell apoptosis and is co-expressed with PNMA5 and PNMA1 to enhance chemosensitivity.
Interestingly, the role of MOAP1 in cancers is relatively well studied in the literature; however, its roles in other diseases are yet to be revealed. Other diseases driven by mitochondrial-dependent apoptosis might also have a close relationship with MOAP1 and need to be further studied. Moreover, it is worthy of mentioning that there are many other biomarkers associated with MOAP1, such as long non-coding RNA (lncRNA), DNA methylation, and histone modification, regulating the expression of a gene. For example, the regulating roles of lncRNA in cancer have been widely studied recently [75], and a few lncRNAs such as SNHG1 also play important roles in neuronal cells [76,77]. The co-expression between lncRNA SNHG5 and MOAP1 in thyroid cancer has been reported; however, the underlying mechanism is unclear [78]. In the future, studies on the relationship between MOAP1 and other regulation molecules should be explored.
Author Contributions
Y.S.: writing, review & editing, visualization. W.W.: review & editing. X.M.: conceptualization, visualization, funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (grant numbers 81671066 and 81974162).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pang, S.W.; Lahiri, C.; Poh, C.L.; Tan, K.O. PNMA family: Protein interaction network and cell signalling pathways implicated in cancer and apoptosis. Cell Signal. 2018, 45, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Tan, K.; Chan, S.L.; Yee, K.S.Y.; Bévort, M.; Ang, K.; Yu, V. MAP-1, a Novel Proapoptotic Protein Containing a BH3-like Motif That Associates with Bax through Its Bcl-2 Homology Domains. J. Biol. Chem. 2001, 276, 2802–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, M.D.; Dallol, A.; Eckfeld, K.; Allen, N.P.; Donninger, H.; Hesson, L.B.; Calvisi, D.; Latif, F.; Clark, G.J. The RASSF1A tumor suppressor activates Bax via MOAP-1. J. Biol. Chem 2006, 281, 4557–4563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertz, C.E.; Bayarri-Olmos, R.; Kirketerp-Moller, N.; van Putten, S.; Pilely, K.; Skjoedt, M.O.; Garred, P. Chimeric Proteins Containing MAP-1 and Functional Domains of C4b-Binding Protein Reveal Strong Complement Inhibitory Capacities. Front. Immunol. 2018, 9, 1945. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.T.; Zhou, Q.L.; Su, Y.C.; Fu, N.Y.; Chang, H.C.; Tao, R.N.; Sukumaran, S.K.; Baksh, S.; Tan, Y.J.; Sabapathy, K.; et al. MOAP-1 Mediates Fas-Induced Apoptosis in Liver by Facilitating tBid Recruitment to Mitochondria. Cell Rep. 2016, 16, 174–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baksh, S.; Tommasi, S.; Fenton, S.; Yu, V.C.; Martins, L.M.; Pfeifer, G.P.; Latif, F.; Downward, J.; Neel, B.G. The tumor suppressor RASSF1A and MAP-1 link death receptor signaling to Bax conformational change and cell death. Mol. Cell 2005, 18, 637–650. [Google Scholar] [CrossRef]
- Ikeda, M.; Kawata, A.; Nishikawa, M.; Tateishi, Y.; Yamaguchi, M.; Nakagawa, K.; Hirabayashi, S.; Bao, Y.; Hidaka, S.; Hirata, Y.; et al. Hippo pathway-dependent and -independent roles of RASSF6. Sci. Signal. 2009, 2, ra59. [Google Scholar] [CrossRef]
- Allen, N.P.; Donninger, H.; Vos, M.D.; Eckfeld, K.; Hesson, L.; Gordon, L.; Birrer, M.J.; Latif, F.; Clark, G.J. RASSF6 is a novel member of the RASSF family of tumor suppressors. Oncogene 2007, 26, 6203–6211. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, H.; Kudo, T.; Maimaiti, S.; Ikeda, M.; Maruyama, J.; Nakagawa, K.; Hata, Y. The RASSF6 tumor suppressor protein regulates apoptosis and the cell cycle via MDM2 protein and p53 protein. J. Biol. Chem. 2013, 288, 30320–30329. [Google Scholar] [CrossRef] [Green Version]
- Van de Vijver, M.J.; He, Y.D.; Veer, L.J.v.; Dai, H.; Hart, A.A.M.; Voskuil, D.W.; Schreiber, G.J.; Peterse, J.L.; Roberts, C.; Marton, M.J.; et al. A Gene-Expression Signature as a Predictor of Survival in Breast Cancer. N. Engl. J. Med. 2002, 347, 1999–2009. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Fu, N.Y.; Sukumaran, S.K.; Wan, K.F.; Wan, Q.; Yu, V.C. TRIM39 is a MOAP-1-binding protein that stabilizes MOAP-1 through inhibition of its poly-ubiquitination process. Exp. Cell Res. 2009, 315, 1313–1325. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.J.; Zhang, L.; Tang, W.; Chen, C.; Yang, C.S.; Kornbluth, S. The Trim39 ubiquitin ligase inhibits APC/CCdh1-mediated degradation of the Bax activator MOAP-1. J. Cell Biol. 2012, 197, 361–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, K.; Huang, N.J.; Cocce, K.; Zhang, L.; Kornbluth, S. Downregulation of the proapoptotic protein MOAP-1 by the UBR5 ubiquitin ligase and its role in ovarian cancer resistance to cisplatin. Oncogene 2017, 36, 1698–1706. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.T.; Chang, H.C.; Zhou, Q.; Yu, C.; Fu, N.Y.; Sabapathy, K.; Yu, V.C. MOAP-1-mediated dissociation of p62/SQSTM1 bodies releases Keap1 and suppresses Nrf2 signaling. EMBO Rep. 2021, 22, e50854. [Google Scholar] [CrossRef]
- Chang, H.C.; Tao, R.N.; Tan, C.T.; Wu, Y.J.; Bay, B.H.; Yu, V.C. The BAX-binding protein MOAP1 associates with LC3 and promotes closure of the phagophore. Autophagy 2021, 17, 3725–3739. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Chen, W.; Kong, D.; Li, X.; Lu, H.; Liu, S.; Wang, J.; Du, L.; Kong, Q.; Huang, X.; et al. miR-25 targets the modulator of apoptosis 1 gene in lung cancer. Carcinogenesis 2015, 36, 925–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.L.; Wang, W.; Lan, X.L.; Zeng, Z.C.; Liang, Y.S.; Yan, Y.R.; Song, F.Y.; Wang, F.F.; Zhu, X.H.; Liao, W.J.; et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol. Cancer 2019, 18, 91. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Liu, D.; Liang, H.; Xue, L.; Su, C.; Liu, M. MiR-1228 promotes breast cancer cell growth and metastasis through targeting SCAI protein. Int. J. Clin. Exp. Pathol. 2015, 8, 6646–6655. [Google Scholar]
- Takaji, M.; Komatsu, Y.; Watakabe, A.; Hashikawa, T.; Yamamori, T. Paraneoplastic antigen-like 5 gene (PNMA5) is preferentially expressed in the association areas in a primate specific manner. Cereb Cortex 2009, 19, 2865–2879. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.J.; Zhao, H.; Hayakawa, K.; Chai, C.; Tan, C.T.; Huang, J.; Tao, R.; Hamanaka, G.; Arumugam, T.V.; Lo, E.H.; et al. Modulator of apoptosis-1 is a potential therapeutic target in acute ischemic injury. J. Cereb. Blood Flow Metab. 2019, 39, 2406–2418. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, G.; Wei, T.; Yang, Z.; Tan, W.; Mo, Z.; Liu, J.; Li, D.; Wei, Y.; Zhang, L.; et al. MicroRNA-25 Protects Smooth Muscle Cells against Corticosterone-Induced Apoptosis. Oxid Med. Cell Longev. 2019, 2019, 2691514. [Google Scholar] [CrossRef] [PubMed]
- Guicciardi, M.E.; GoRes., G.J. Apoptosis: A mechanism of acute and chronic liver injury. Gut 2005, 54, 1024–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, M.E.; Budd, R.C.; Desbarats, J.; Hedrick, S.M.; Hueber, A.O.; Newell, M.K.; Owen, L.B.; Pope, R.M.; Tschopp, J.; Wajant, H.; et al. The CD95 receptor: Apoptosis revisited. Cell 2007, 129, 447–450. [Google Scholar] [CrossRef] [PubMed]
- Strasser, A.; Jost, P.J.; Nagata, S. The many roles of FAS receptor signaling in the immune system. Immunity 2009, 30, 180–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryo, K.; Kamogawa, Y.; Ikeda, I.; Yamauchi, K.; Yonehara, S.; Nagata, S.; Hayashi, N. Significance of Fas antigen-mediated apoptosis in human fulminant hepatic failure. Am. J. Gastroenterol. 2000, 95, 2047–2055. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Iwasa, H.; Jiang, X.; Hata, Y. RASSF6; the Putative Tumor Suppressor of the RASSF Family. Cancers 2015, 7, 2415–2426. [Google Scholar] [CrossRef]
- Wang, N.; He, X.; Zhou, R.; Jia, G.; Qiao, Q. STAT3 induces colorectal carcinoma progression through a novel miR-572-MOAP-1 pathway. Onco Targets Ther. 2018, 11, 3475–3484. [Google Scholar] [CrossRef] [Green Version]
- Travis, W.D.; Brambilla, E.; Noguchi, M.; Nicholson, A.G.; Geisinger, K.R.; Yatabe, Y.; Beer, D.G.; Powell, C.A.; Riely, G.J.; Van Schil, P.E.; et al. International association for the study of lung cancer/american thoracic society/european respiratory society international multidisciplinary classification of lung adenocarcinoma. J. Thorac. Oncol. 2011, 6, 244–285. [Google Scholar] [CrossRef] [Green Version]
- Van Delft, M.F.; Huang, D.C. How the Bcl-2 family of proteins interact to regulate apoptosis. Cell Res. 2006, 16, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Pei, F.; Yang, F.; Li, L.; Amin, A.D.; Liu, S.; Buchan, J.R.; Cho, W.C. Role of Autophagy and Apoptosis in Non-Small-Cell Lung Cancer. Int. J. Mol. Sci. 2017, 18, 367. [Google Scholar] [CrossRef] [PubMed]
- Othman, N.; Nagoor, N.H. The role of microRNAs in the regulation of apoptosis in lung cancer and its application in cancer treatment. Biomed. Res. Int. 2014, 2014, 318030. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Aghabalazade, A.; Shoorei, H.; Majidpoor, J.; Taheri, M.; Mokhtari, M. The Impact of lncRNAs and miRNAs on Apoptosis in Lung Cancer. Front Oncol. 2021, 11, 714795. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; He, Y.; Wang, Y.; Li, X.; Young, J.; Ioannidis, J.; Dunlop, M.G.; Theodoratou, E. Risk factors and risk prediction models for colorectal cancer metastasis and recurrence: An umbrella review of systematic reviews and meta-analyses of observational studies. BMC Med. 2020, 18, 172. [Google Scholar] [CrossRef]
- Katkoori, V.; Suarez-Cuervo, C.; Shanmugam, C.; Jhala, N.; Callens, T.; Messiaen, L.; Posey, J.; Bumpers, H.; Meleth, S.; Grizzle, W.; et al. Bax expression is a candidate prognostic and predictive marker of colorectal cancer. J. Gastrointest. Oncol. 2010, 1, 76–89. [Google Scholar]
- Sturm, I.; Kohne, C.H.; Wolff, G.; Petrowsky, H.; Hillebrand, T.; Hauptmann, S.; Lorenz, M.; Dorken, B.; Daniel, P.T. Analysis of the p53/BAX pathway in colorectal cancer: Low BAX is a negative prognostic factor in patients with resected liver metastases. J. Clin. Oncol. 1999, 17, 1364–1374. [Google Scholar] [CrossRef]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 Ratio Considered as a Prognostic Marker with Age and Tumor Location in Colorectal Cancer? Iran. Biomed. J. 2015, 19, 69–75. [Google Scholar]
- Jansson, A.; Sun, X. Bax expression decreases significantly from primary tumor to metastasis in colorectal cancer. J. Clin. Oncol. 2002, 20, 811–816. [Google Scholar] [CrossRef]
- D’Errico, I.; Sasso, G.; Salvatore, L.; Murzilli, S.; Martelli, N.; Cristofaro, M.; Latorre, D.; Villani, G.; Moschetta, A. Bax is necessary for PGC1α pro-apoptotic effect in colorectal cancer cells. Cell Cycle 2011, 10, 2937–2945. [Google Scholar] [CrossRef] [Green Version]
- Gil, J.; Ramsey, D.; Szmida, E.; Leszczynski, P.; Pawlowski, P.; Bebenek, M.; Sasiadek, M. The BAX gene as a candidate for negative autophagy-related genes regulator on mRNA levels in colorectal cancer. Med. Oncol. 2017, 34, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yashiro, M.; Hirakawa, K.; Boland, R. Mutations in TGFbeta-RII and BAXmediate tumor progression in the later stages of colorectal cancer with microsatellite instability. BMC Cancer 2010, 10, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, M.; Milner, J. Bcl-2 constitutively suppresses p53-dependent apoptosis in colorectal cancer cells. Genes Dev. 2003, 17, 832–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehler, B.C.; Scherr, A.L.; Lorenz, S.; Urbanik, T.; Kautz, N.; Elssner, C.; Welte, S.; Bermejo, J.L.; Jager, D.; Schulze-Bergkamen, H. Beyond cell death—Antiapoptotic Bcl-2 proteins regulate migration and invasion of colorectal cancer cells in vitro. PLoS ONE 2013, 8, e76446. [Google Scholar] [CrossRef] [PubMed]
- Perez, E.A.; Romond, E.H.; Suman, V.J.; Jeong, J.H.; Sledge, G.; Geyer, C.E., Jr.; Martino, S.; Rastogi, P.; Gralow, J.; Swain, S.M.; et al. Trastuzumab Plus Adjuvant Chemotherapy for Human Epidermal Growth Factor Receptor 2–Positive Breast Cancer: Planned JoInt. Analysis of Overall Survival from NSABP B-31 and NCCTG N9831. J. Clin. Oncol. 2014, 32, 3744. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Eiermann, W.; Robert, N.J.; Giermek, J.; Martin, M.; Jasiowka, M.; Mackey, J.R.; Chan, A.; Liu, M.C.; Pinter, T. Abstract S5–04: Ten year follow-up of BCIRG-006 comparing doxorubicin plus cyclophosphamide followed by docetaxel (AC→T) with doxorubicin plus cyclophosphamide followed by docetaxel and trastuzumab (AC→TH) with docetaxel, carboplatin and trastuzumab (TCH) in HER2+ early breast cancer. Cancer Res. 2016, 76, S5-04. [Google Scholar]
- Cameron, D.; Piccart-Gebhart, M.J.; D Gelber, R.; Procter, M.; Goldhirsch, A.; de Azambuja, E.; Castro Jr, G.; Untch, M.; Smith, I.; Gianni, L.; et al. 11 years’ follow-up of trastuzumab after adjuvant chemotherapy in HER2-positive early breast cancer: Final analysis of the HERceptin Adjuvant (HERA) trial. Lancet 2017, 389, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Sjostrom, J.; Blomqvist, C.; von Boguslawski, K.; Bengtsson, N.O.; Mjaaland, I.; Malmstrom, P.; Ostenstadt, B.; Wist, E.; Valvere, V.; Takayama, S.; et al. The predictive value of bcl-2, bax, bcl-xL.; bag-1, fas, and fasL for chemotherapy response in advanced breast cancer. Clin. Cancer Res. 2002, 8, 811–816. [Google Scholar]
- Azimian, H.; Dayyani, M.; Toossi, M.; Mahmoudi, M. Bax/Bcl-2 expression ratio in prediction of response to breast cancer radiotherapy. Iran. J. Basic Med. Sci. 2018, 21, 325–332. [Google Scholar]
- Binder, C.; Marx, D.; Binder, L.; Schauer, A.; Hiddemann, W. Expression of Bax in relation to Bcl-2 and other predictive parameters in breast cancer. Ann. Oncol. 1996, 7, 129–133. [Google Scholar] [CrossRef]
- Lee, Y.H.; Pang, S.W.; Revai Lechtich, E.; Shah, K.; Simon, S.E.; Ponnusamy, S.; Narayanan, R.; Poh, C.L.; Tan, K.O. Tricistronic expression of MOAP-1, Bax and RASSF1A in cancer cells enhances chemo-sensitization that requiRes. BH3L domain of MOAP-1. J. Cancer Res. Clin. Oncol. 2020, 146, 1751–1764. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Pang, S.W.; Poh, C.L.; Tan, K.O. Distinct functional domains of PNMA5 mediate protein-protein interaction, nuclear localization, and apoptosis signaling in human cancer cells. J. Cancer Res. Clin. Oncol. 2016, 142, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Honig, L.S.; Rosenberg, R.N. Apoptosis and neurologic disease. Am. J. Med 2000, 108, 317–330. [Google Scholar] [CrossRef]
- Cervio, E.; Volta, U.; Verri, M.; Boschi, F.; Pastoris, O.; Granito, A.; Barbara, G.; Parisi, C.; Felicani, C.; Tonini, M.; et al. Sera of patients with celiac disease and neurologic disorders evoke a mitochondrial-dependent apoptosis in vitro. Gastroenterology 2007, 133, 195–206. [Google Scholar] [CrossRef]
- Zhao, H.; Mohamed, N.E.; Chan, S.J.; Tan, C.T.; Tao, R.; Yu, V.C.; Wong, P.T. Absence of Stress Response in Dorsal Raphe Nucleus in Modulator of Apoptosis 1-Deficient Mice. Mol. Neurobiol. 2019, 56, 2185–2201. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, L.O.; Gellert, P.; White, M.; Araujo-SoaRes, V.; Ford, G.A.; Mackintosh, J.E.; Rodgers, H.; Sniehotta, F.F.; Thomson, R.G.; Dombrowski, S.U. The Recognition-Response Gap in Acute Stroke: Examining the Relationship between Stroke Recognition and Response in a General Population Survey. J. Stroke Cerebrovasc. Dis. 2020, 29, 104499. [Google Scholar] [CrossRef]
- Krajewski, S.; Mai, J.K.; Krajewska, M.; Sikorska, M.; Mossakowski, M.J.; Reed, J.C. Upregulation of bax protein levels in neurons following cerebral ischemia. J. Neurosci. 1995, 15, 6364–6376. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, R.L.; Nakayama, M.; Kawaguchi, K.; Jin, K.; Stetler, R.A.; Simon, R.P.; Graham, S.H. Expression of the apoptosis-effector gene, Bax, is up-regulated in vulnerable hippocampal CA1 neurons following global ischemia. J. Neurochem. 1996, 67, 64–71. [Google Scholar] [CrossRef]
- Hochhauser, E.; Kivity, S.; Offen, D.; Maulik, N.; Otani, H.; Barhum, Y.; Pannet, H.; Shneyvays, V.; Shainberg, A.; Goldshtaub, V.; et al. Bax ablation protects against myocardial ischemia-reperfusion injury in transgenic mice. Am. J. Physiol. Heart Circ. Physiol. 2003, 284, H2351–H2359. [Google Scholar] [CrossRef] [Green Version]
- Jiao, S.; Li, Z. Nonapoptotic function of BAD and BAX in long-term depression of synaptic transmission. Neuron 2011, 70, 758–772. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Liu, X.; Zhang, R.; Cheng, K.; Mu, J.; Fang, L.; Xie, P. Increased apoptosis and different regulation of pro-apoptosis protein bax and anti-apoptosis protein bcl-2 in the olfactory bulb of a rat model of depression. Neurosci. Lett 2011, 504, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Nikoui, V.; Naveed, A.; Mumtaz, F.; Zaman, H.; Haider, A.; Aman, W.; Wahab, A.; Khan, S.N.; Ullah, N.; et al. Antidepressant-like effect of ethanol in mice forced swimming test is mediated via inhibition of NMDA/nitric oxide/cGMP signaling pathway. Alcohol 2021, 92, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, J.; Yao-Yao, B.I.; Chen, Z.; Zheng, Q.H.; Duan, H.F. Automatic Recognition of Breast Gland Based on Two-step Clustering and Random Forest. Comput. Sci. 2018, 45, 247–252. [Google Scholar]
- Foffani, G.; Obeso, J.A. A Cortical Pathogenic Theory of Parkinson’s Disease. Neuron 2018, 99, 1116–1128. [Google Scholar] [CrossRef] [Green Version]
- Aouacheria, A.; Baghdiguian, S.; Lamb, H.M.; Huska, J.D.; Pineda, F.J.; Hardwick, J.M. Connecting mitochondrial dynamics and life-or-death events via Bcl-2 family proteins. Neurochem. Int. 2017, 109, 141–161. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Yang, K.; Wang, R.; Liu, G.; Shu, Z.; Wang, N.; Zhang, R.; Yu, J.; Chen, J.; Li, X.; Zhou, X. HerGePred: Heterogeneous Network Embedding Representation for Disease Gene Prediction. IEEE J. Biomed. Health Inform. 2019, 23, 1805–1815. [Google Scholar] [CrossRef]
- Vila, M.; Jackson-Lewis, V.; Vukosavic, S.; Djaldetti, R.; Liberatore, G.; Offen, D.; Korsmeyer, S.J.; Przedborski, S. Bax ablation prevents dopaminergic neurodegeneration in the 1-methyl- 4-phenyl-1,2,3,6-tetrahydropyridine mouse model of Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2001, 98, 2837–2842. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Zhang, M.; Wei, M.; Wang, A.; Deng, Y.; Cao, H. MicroRNA-216a inhibits neuronal apoptosis in a cellular Parkinson’s disease model by targeting Bax. Metab. Brain Dis. 2020, 35, 627–635. [Google Scholar] [CrossRef]
- Capurro, A.; Bodea, L.G.; Schaefer, P.; Luthi-Carter, R.; Perreau, V.M. Computational deconvolution of genome wide expression data from Parkinson’s and Huntington’s disease brain tissues using population-specific expression analysis. Front Neurosci. 2014, 8, 441. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.H.; Pang, S.W.; Tan, K.O. PNMA2 mediates heterodimeric interactions and antagonizes chemo-sensitizing activities mediated by members of PNMA family. Biochem. Biophys. Res. Commun. 2016, 473, 224–229. [Google Scholar] [CrossRef]
- Walther, D.J.; Peter, J.; Bashammakh, S.; Hörtnagl, H.; Voits, M.; Fink, H.; Bader, M. Synthesis of Serotonin by a Second Tryptophan Hydroxylase Isoform. Science 2003, 299, 76. [Google Scholar] [CrossRef] [PubMed]
- Li, G.M.; Zhang, C.L.; Rui, R.P.; Sun, B.; Guo, W. Bioinformatics analysis of common differential genes of coronary artery disease and ischemic cardiomyopathy. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3553–3569. [Google Scholar] [PubMed]
- Wang, Q.; Liu, B.; Wang, Y.; Bai, B.; Yu, T.; Chu, X.M. The biomarkers of key miRNAs and target genes associated with acute myocardial infarction. PeerJ 2020, 8, e9129. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.C.; Ni, J.J.; Cui, W.Y.; Wang, B.Y.; Zhuo, W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am. J. Cancer Res. 2019, 9, 1354–1366. [Google Scholar] [PubMed]
- Wang, H.; Lu, B.; Chen, J. Knockdown of lncRNA SNHG1 attenuated Abeta25-35-inudced neuronal injury via regulating KREMEN1 by acting as a ceRNA of miR-137 in neuronal cells. Biochem. Biophys. Res. Commun. 2019, 518, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Simchovitz, A.; Hanan, M.; Yayon, N.; Lee, S.; Bennett, E.R.; Greenberg, D.S.; Kadener, S.; Soreq, H. A lncRNA survey finds increases in neuroprotective LINC-PINT in Parkinson’s disease substantia nigra. Aging Cell 2020, 19, e13115. [Google Scholar] [CrossRef] [Green Version]
- Murugan, A.K.; Munirajan, A.K.; Alzahrani, A.S. Long noncoding RNAs: Emerging players in thyroid cancer pathogenesis. Endocr. Relat. Cancer 2018, 25, R59–R82. [Google Scholar] [CrossRef]
Figure 1.
The common functions of MOAP1 and its related diseases.

Figure 2.
MOAP1 acts as a modulator on cancer-associated miRNAs and cancers.

Figure 3.
MOAP1 interacts with other genes to perform cancer-related functions and the literature supporting the interactions and functions.
Figure 3.
MOAP1 interacts with other genes to perform cancer-related functions and the literature supporting the interactions and functions.


{kind=link}
{kind=link}
{kind=link}
Table 1.
The molecules involved in the action of MOAP1 in neuron-related traits or diseases and their functions.
Table 1.
The molecules involved in the action of MOAP1 in neuron-related traits or diseases and their functions.
| Gene Symbol | Neuron-Related Traits or Diseases | Function |
|---|---|---|
| PNMA2 | Abundant in brain | Antagonize the signal transduction of apoptotic cells both MOAP1 and PNMA1 [71] |
| PNMA3 | Abundant in brain | Gene Ontology (GO) annotations related to this gene include nucleic acid binding (https://www.genecards.org/ accessed on 9 December 2021) |
| BAX | Ischemic injury | Binding with a novel BAX-related protein (MOAP1) mediates apoptosis [2] |
| TPH2 | Depression | Catalytic biosynthesis of serotonin [72] |
| BDNF | Depression | Bind with its homologous receptor promotes neuronal survival in the adult brain (https://www.genecards.org accessed on 9 December 2021) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Su, Y.; Wang, W.; Meng, X. Revealing the Roles of MOAP1 in Diseases: A Review. Cells 2022, 11, 889. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11050889
AMA Style
Su Y, Wang W, Meng X. Revealing the Roles of MOAP1 in Diseases: A Review. Cells. 2022; 11(5):889. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11050889
Chicago/Turabian StyleSu, Yanfang, Weixia Wang, and Xianfang Meng. 2022. "Revealing the Roles of MOAP1 in Diseases: A Review" Cells 11, no. 5: 889. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11050889
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.