
Palmitate-Triggered COX2/PGE2-Related Hyperinflammation in Dual-Stressed PdL Fibroblasts Is Mediated by Repressive H3K27 Trimethylation

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Stimulation with Palmitic Acid
2.3. Stimulation with P. gingivalis Lipopolysaccharides
2.4. UNC1999 Application
2.5. Application of Compressive Force
2.6. Expression Analysis with Quantitative PCR
2.7. Immunofluorescent Staining
2.8. THP1 Adhesion Assay
2.9. MTT Assay
2.10. Nuclear Extraction and Histone Methyltransferase Activity Assay
2.11. TUNEL Assay
2.12. ELISA
2.13. Chromatin-Immunoprecipitation
2.14. Microscopy and Image Analysis
2.15. Statistics
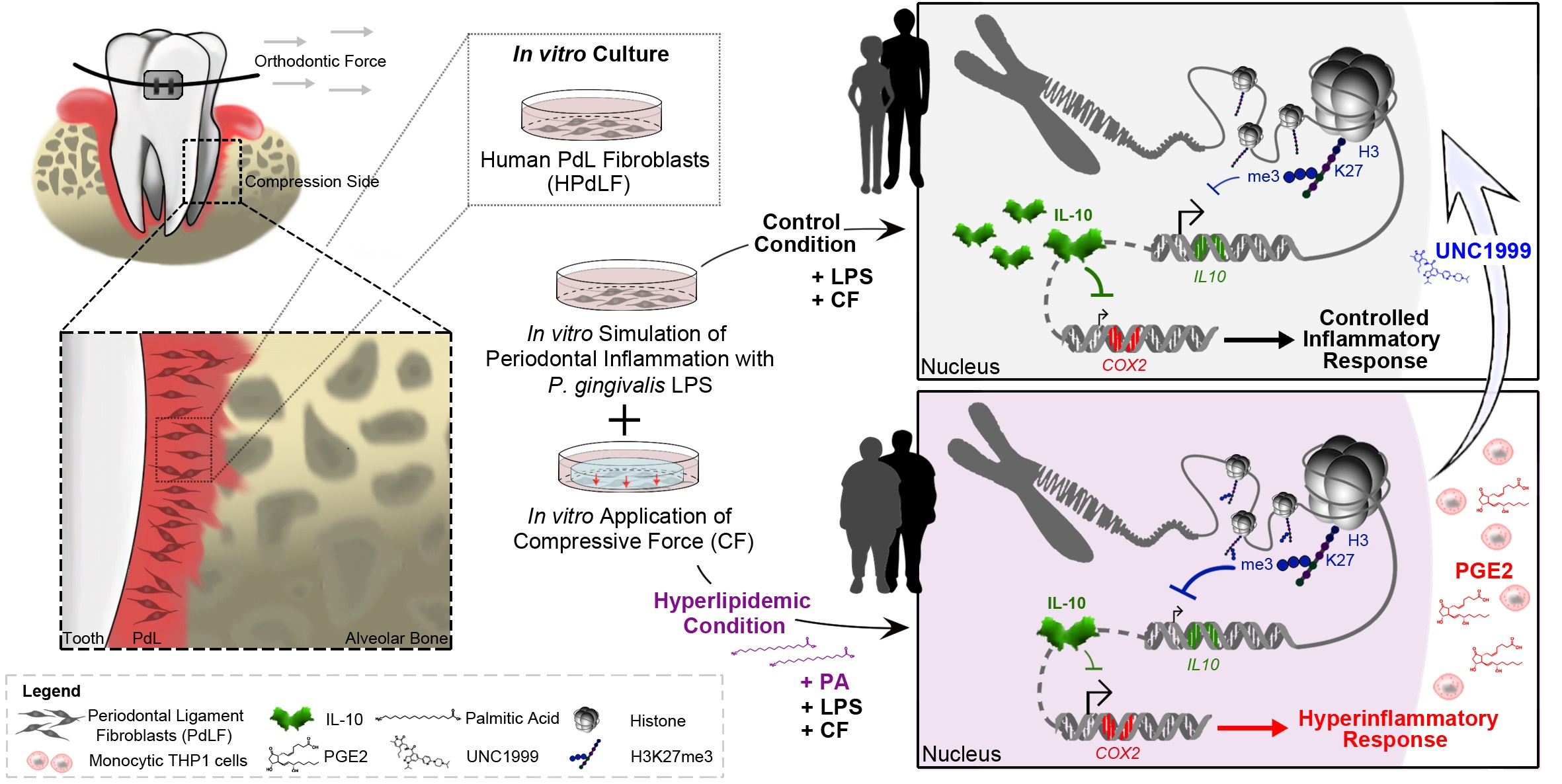
3. Results
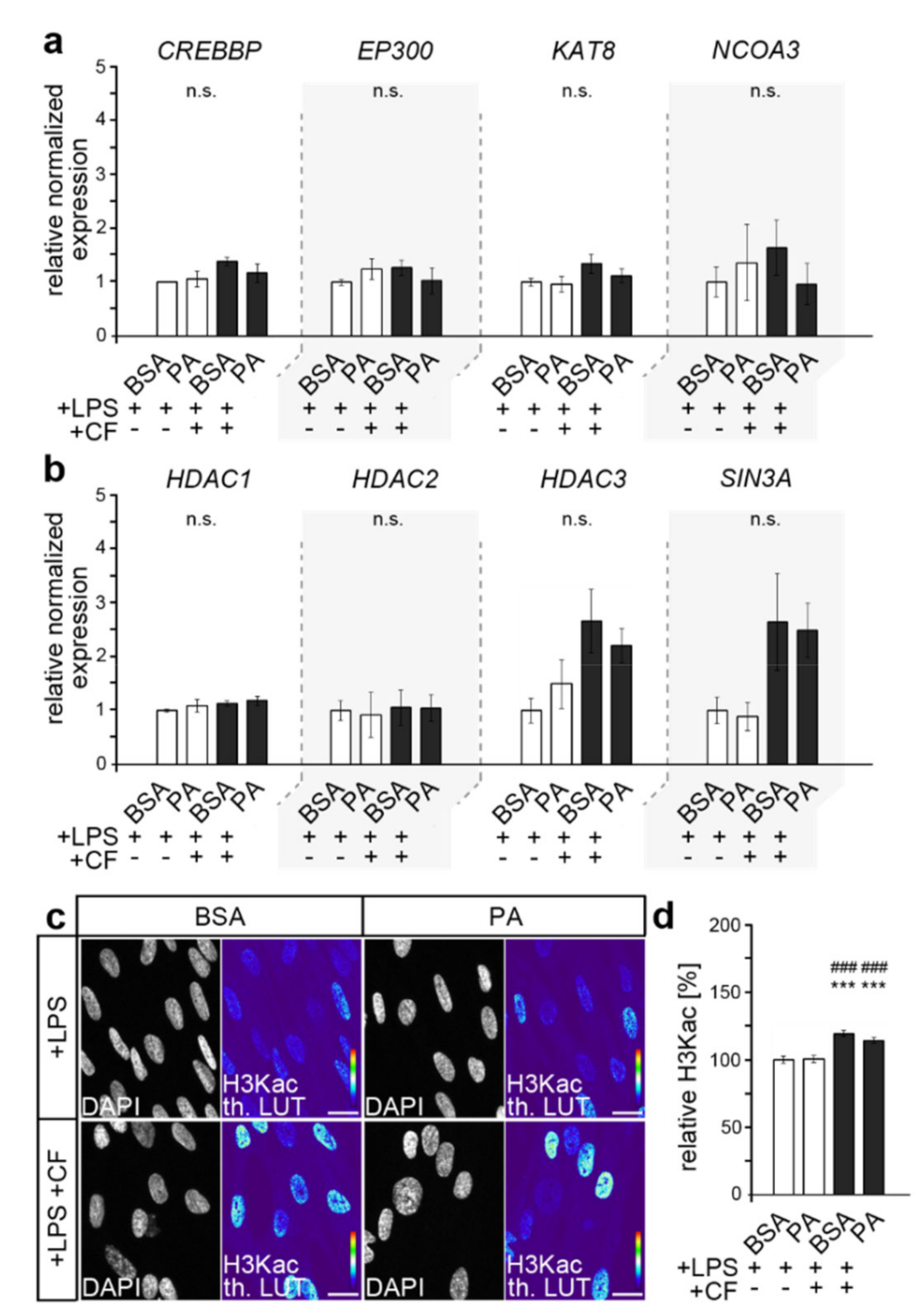
3.1. Force-Induced Increase in H3 Lysine Acetylation in Human Periodontal Fibroblasts Is Not Affected by Palmitic Acid
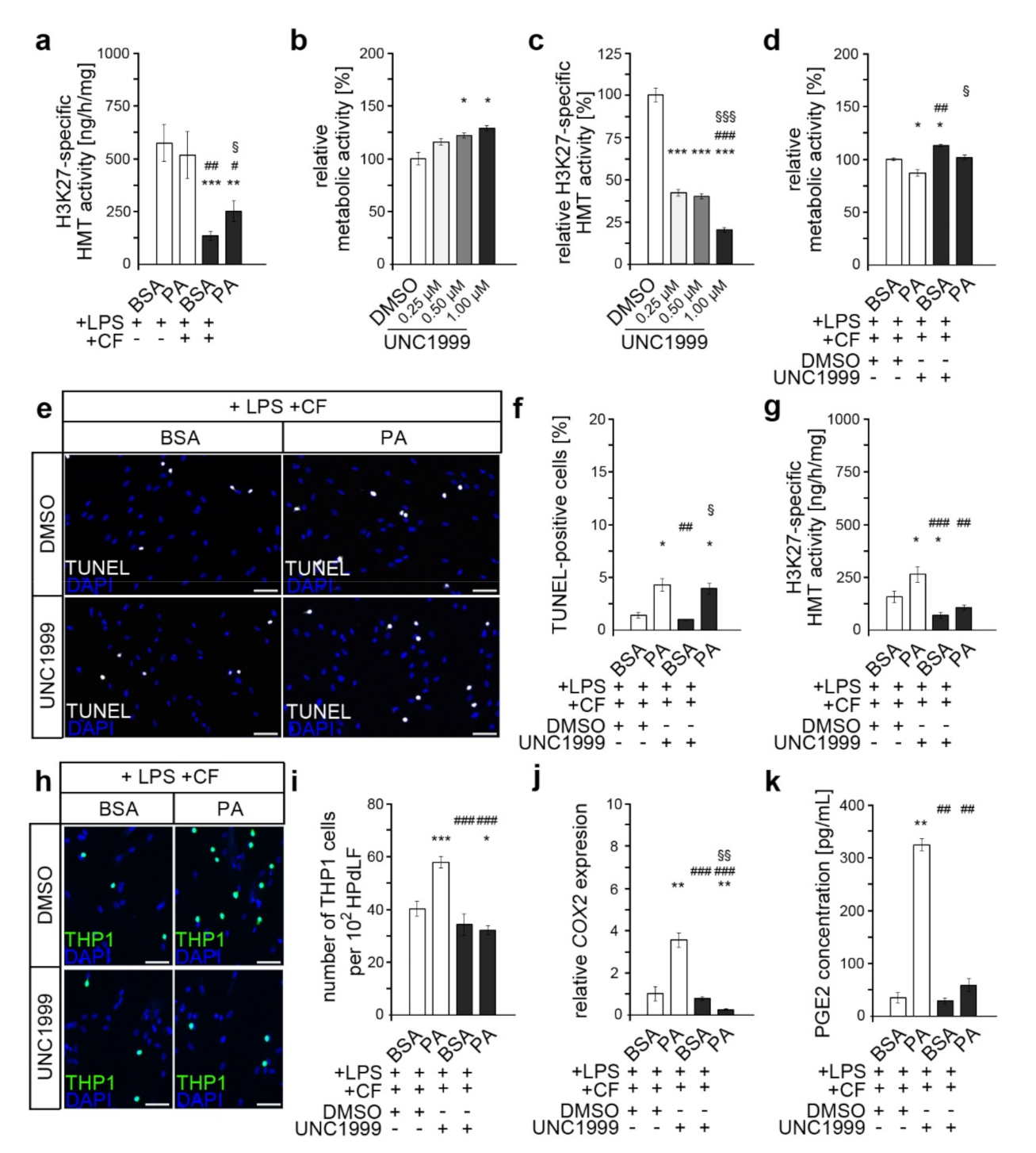
3.2. Palmitic Acid Impacts Force-Induced Reduction in H3K27me3 in LPS-Stimulated HPdLFs
3.3. Inhibition of PRC2 Enzymes EZH1 and EZH2 Abrogates PA-Induced Excessive Inflammation, Possibly via COX2/PGE2 Modulation
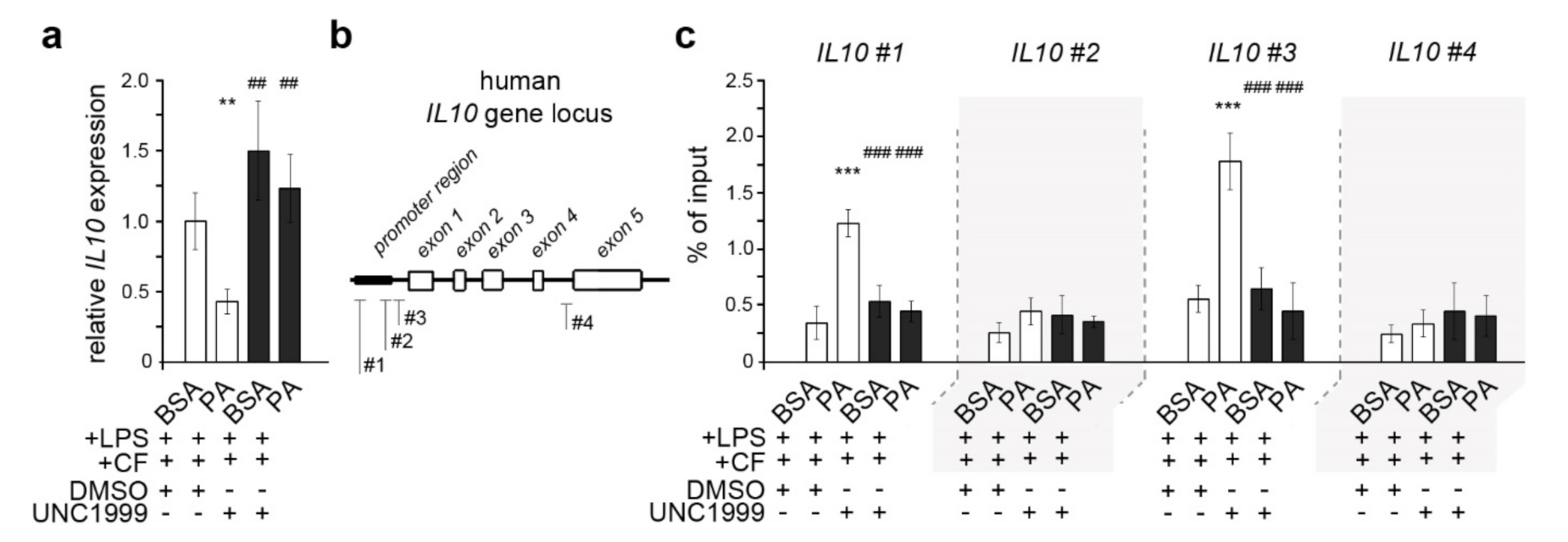
3.4. Palmitic Acid Causes Enhanced Trimethylation of H3K27 at Il10 Promoter-Associated Regions in Dual-Stimulated HPdLFs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Larsson, L. Current Concepts of Epigenetics and Its Role in Periodontitis. Curr. Oral Health Rep. 2017, 4, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Batchelor, P. Is periodontal disease a public health problem? Br. Dent. J. 2014, 217, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Offenbacher, S.; Barros, S.P.; Beck, J.D. Rethinking Periodontal Inflammation. J. Periodontol. 2008, 79, 1577–1584. [Google Scholar] [CrossRef]
- Bird, A. Perceptions of epigenetics. Nature 2007, 447, 396–398. [Google Scholar] [CrossRef]
- Jurdziński, K.T.; Potempa, J.; Grabiec, A.M. Epigenetic regulation of inflammation in periodontitis: Cellular mechanisms and therapeutic potential. Clin. Epigenetics 2020, 12, 186. [Google Scholar] [CrossRef]
- Rothbart, S.B.; Strahl, B.D. Interpreting the language of histone and DNA modifications. Biochim. Biophys. Acta 2014, 1839, 627–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer, E.L.; Shi, Y. Histone methylation: A dynamic mark in health, disease and inheritance. Nat. Rev. Genet. 2012, 13, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Niederman, R.; Buyle-Bodin, Y.; Lu, B.-Y.; Naleway, C.; Robinson, P.; Kent, R. The relationship of gingival crevicular fluid short chain carboxylic acid concentration to gingival inflammation. J. Clin. Periodontol. 1996, 23, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the toll-like receptor 4/nf-kappab pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, A.A.; Connaughton, R.M.; Lyons, C.; McMorrow, A.M.; Roche, H.M. Fatty acids and chronic low grade inflammation associated with obesity and the metabolic syndrome. Eur. J. Pharmacol. 2016, 785, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Rössner, S.; Walldius, G.; Björvell, H. Fatty acid composition in serum lipids and adipose tissue in severe obesity before and after six weeks of weight loss. Int. J. Obes. 1989, 13, 603–612. [Google Scholar]
- Sokolova, M.; Vinge, L.E.; Alfsnes, K.; Olsen, M.B.; Eide, L.; Kaasbøll, O.J.; Attramadal, H.; Torp, M.-K.; Fosshaug, L.E.; Rashidi, A.; et al. Palmitate promotes inflammatory responses and cellular senescence in cardiac fibroblasts. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Pillon, N.J.; Azizi, P.M.; Li, Y.E.; Liu, J.; Wang, C.; Chan, K.L.; Hopperton, K.E.; Bazinet, R.P.; Heit, B.; Bilan, P.J.; et al. Palmitate-induced inflammatory pathways in human adipose microvascular endothelial cells promote monocyte adhesion and impair insulin transcytosis. Am. J. Physiol. Metab. 2015, 309, E35–E44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemecz, M.; Constantin, A.; Dumitrescu, M.; Alexandru, N.; Filippi, A.; Tanko, G.; Georgescu, A. The Distinct Effects of Palmitic and Oleic Acid on Pancreatic Beta Cell Function: The Elucidation of Associated Mechanisms and Effector Molecules. Front. Pharmacol. 2019, 9, 1554. [Google Scholar] [CrossRef] [Green Version]
- Gillet, C.; Spruyt, D.; Rigutto, S.; Valle, A.D.; Berlier, J.; Louis, C.; Debier, C.; Gaspard, N.; Malaisse, W.J.; Gangji, V.; et al. Oleate Abrogates Palmitate-Induced Lipotoxicity and Proinflammatory Response in Human Bone Marrow-Derived Mesenchymal Stem Cells and Osteoblastic Cells. Endocrinology 2015, 156, 4081–4093. [Google Scholar] [CrossRef] [Green Version]
- De Souza, C.O.; Valenzuela, C.A.; Baker, E.J.; Miles, E.A.; Neto, J.C.R.; Calder, P.C. Palmitoleic Acid has Stronger Anti-Inflammatory Potential in Human Endothelial Cells Compared to Oleic and Palmitic Acids. Mol. Nutr. Food Res. 2018, 62, e1800322. [Google Scholar] [CrossRef]
- Carrillo, C.; Cavia Mdel, M.; Alonso-Torre, S. Role of oleic acid in immune system; mechanism of action: A review. Nutr. Hosp. 2012, 27, 978–990. [Google Scholar] [PubMed] [Green Version]
- de Lima-Salgado, T.M.; Alba-Loureiro, T.C.; Nascimento, C.S.D.; Nunes, M.T.; Curi, R. Molecular Mechanisms by Which Saturated Fatty Acids Modulate TNF-α Expression in Mouse Macrophage Lineage. Cell Biophys. 2010, 59, 89–97. [Google Scholar] [CrossRef]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Agents Actions 2019, 68, 915–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.R.; Zhang, J.A.; Zhang, Q.; Permatasari, F.; Xu, Y.; Wu, D.; Yin, Z.Q.; Luo, D. Palmitic acid induces production of proinflammatory cytokines interleukin-6, interleukin-1beta, and tumor necrosis factor-alpha via a nf-kappab-dependent mechanism in hacat keratinocytes. Mediat. Inflamm. 2013, 2013, 530429. [Google Scholar] [CrossRef] [Green Version]
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; LaRocca, A.V.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M.; et al. Fatty Acids in Membranes as Homeostatic, Metabolic and Nutritional Biomarkers: Recent Advancements in Analytics and Diagnostics. Diagnostics 2016, 7, 1. [Google Scholar] [CrossRef]
- Amar, S.; Zhou, Q.; Shaik-Dasthagirisaheb, Y.; Leeman, S. Diet-induced obesity in mice causes changes in immune responses and bone loss manifested by bacterial challenge. Proc. Natl. Acad. Sci. USA 2007, 104, 20466–20471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, M.; Ohara, N. Molecular mechanisms of Porphyromonas gingivalis -host cell interaction on periodontal diseases. Jpn. Dent. Sci. Rev. 2017, 53, 134–140. [Google Scholar] [CrossRef]
- Jönsson, D.; Nebel, D.; Bratthall, G.; Nilsson, B.-O. The human periodontal ligament cell: A fibroblast-like cell acting as an immune cell. J. Periodontal Res. 2010, 46, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Proff, P.; Reicheneder, C.; Faltermeier, A.; Kubein-Meesenburg, D.; Römer, P. Effects of mechanical and bacterial stressors on cytokine and growth-factor expression in periodontal ligament cells. J. Orofac. Orthop. 2014, 75, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Fukasawa, S. Is Inflammation a Friend or Foe for Orthodontic Treatment?: Inflammation in Orthodontically Induced Inflammatory Root Resorption and Accelerating Tooth Movement. Int. J. Mol. Sci. 2021, 22, 2388. [Google Scholar] [CrossRef]
- Weltman, B.; Vig, K.W.; Fields, H.W.; Shanker, S.; Kaizar, E.E. Root resorption associated with orthodontic tooth movement: A systematic review. Am. J. Orthod. Dentofac. Orthop. 2010, 137, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Wishney, M. Potential risks of orthodontic therapy: A critical review and conceptual framework. Aust. Dent. J. 2017, 62, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Kirschneck, C.; Fanghänel, J.; Wahlmann, U.; Wolf, M.; Roldán, J.C.; Proff, P. Interactive effects of periodontitis and orthodontic tooth movement on dental root resorption, tooth movement velocity and alveolar bone loss in a rat model. Ann. Anat. Anat. Anz. 2017, 210, 32–43. [Google Scholar] [CrossRef]
- Jayachandran, T.; Srinivasan, B.; Padmanabhan, S. Salivary leptin levels in normal weight and overweight individuals and their correlation with orthodontic tooth movement. Angle Orthod. 2017, 87, 739–744. [Google Scholar] [CrossRef] [Green Version]
- Michelogiannakis, D.; Rossouw, P.E.; Khan, J.; Akram, Z.; Menenakos, E.; Javed, F. Influence of increased body mass index on orthodontic tooth movement and related parameters in children and adolescents: A systematic review of longitudinal controlled clinical studies. J. Orthod. 2019, 46, 323–334. [Google Scholar] [CrossRef]
- Saloom, H.; Papageorgiou, S.; Carpenter, G.; Cobourne, M. Impact of Obesity on Orthodontic Tooth Movement in Adolescents: A Prospective Clinical Cohort Study. J. Dent. Res. 2017, 96, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Consolaro, A. Obesity and orthodontic treatment: Is there any direct relationship? Dental. Press. J. Orthod. 2017, 22, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Lopez, O.; Milagro, F.I.; Riezu-Boj, J.I.; Martinez, J.A. Epigenetic signatures underlying inflammation: An interplay of nutrition, physical activity, metabolic diseases, and environmental factors for personalized nutrition. Agents Actions 2020, 70, 29–49. [Google Scholar] [CrossRef]
- McDonnell, E.; Crown, S.B.; Fox, D.B.; Kitir, B.; Ilkayeva, O.R.; Olsen, C.A.; Grimsrud, P.A.; Hirschey, M.D. Lipids Reprogram Metabolism to Become a Major Carbon Source for Histone Acetylation. Cell Rep. 2016, 17, 1463–1472. [Google Scholar] [CrossRef] [Green Version]
- Pascual, G.; Domínguez, D.; Elosúa-Bayes, M.; Beckedorff, F.; Laudanna, C.; Bigas, C.; Douillet, D.; Greco, C.; Symeonidi, A.; Hernández, I.; et al. Dietary palmitic acid promotes a prometastatic memory via Schwann cells. Nature 2021, 599, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pamulapati, H.; Tikoo, K. Fatty acid induced metabolic memory involves alterations in renal histone H3K36me2 and H3K27me3. Mol. Cell. Endocrinol. 2016, 422, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Heerboth, S.; Lapinska, K.; Snyder, N.; Leary, M.; Rollinson, S.; Sarkar, S. Use of Epigenetic Drugs in Disease: An Overview. Genet. Epigenetics 2014, 6, 12270. [Google Scholar] [CrossRef]
- Symmank, J.; Appel, S.; Bastian, J.; Knaup, I.; Marciniak, J.; Hennig, C.-L.; Döding, A.; Schulze-Späte, U.; Jacobs, C.; Wolf, M. Hyperlipidemic Conditions Impact Force-Induced Inflammatory Response of Human Periodontal Ligament Fibroblasts Concomitantly Challenged with P. gingivalis-LPS. Int. J. Mol. Sci. 2021, 22, 6069. [Google Scholar] [CrossRef] [PubMed]
- Greenhough, A.; Smartt, H.J.; Moore, A.E.; Roberts, H.R.; Williams, A.C.; Paraskeva, C.; Kaidi, A. The cox-2/pge2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 2009, 30, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Kirschneck, C.; Batschkus, S.; Proff, P.; Kostler, J.; Spanier, G.; Schroder, A. Valid gene expression normalization by rt-qpcr in studies on hpdl fibroblasts with focus on orthodontic tooth movement and periodontitis. Sci. Rep. 2017, 7, 14751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symmank, J.; Chorus, M.; Appel, S.; Marciniak, J.; Knaup, I.; Bastian, A.; Hennig, C.-L.; Döding, A.; Schulze-Späte, U.; Jacobs, C.; et al. Distinguish fatty acids impact survival, differentiation and cellular function of periodontal ligament fibroblasts. Sci. Rep. 2020, 10, 15706. [Google Scholar] [CrossRef] [PubMed]
- Stemmler, A.; Symmank, J.; Steinmetz, J.; von Brandenstein, K.; Hennig, C.L.; Jacobs, C. Gdf15 supports the inflammatory response of pdl fibroblasts stimulated by P. gingivalis lps and concurrent compression. Int. J. Mol. Sci. 2021, 22, 3608. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Symmank, J.; Bayer, C.; Reichard, J.; Pensold, D.; Zimmer-Bensch, G. Neuronal Lhx1 expression is regulated by DNMT1-dependent modulation of histone marks. Epigenetics 2020, 15, 1259–1274. [Google Scholar] [CrossRef]
- Haring, M.; Offermann, S.; Danker, T.; Horst, I.; Peterhansel, C.; Stam, M. Chromatin immunoprecipitation: Optimization, quantitative analysis and data normalization. Plant. Methods 2007, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Symmank, J.; Bayer, C.; Schmidt, C.; Hahn, A.; Pensold, D.; Zimmer-Bensch, G. DNMT1 modulates interneuron morphology by regulating Pak6 expression through crosstalk with histone modifications. Epigenetics 2018, 13, 536–556. [Google Scholar] [CrossRef] [Green Version]
- Schuldt, L.; von Brandenstein, K.; Jacobs, C.; Symmank, J. Oleic acid-related anti-inflammatory effects in force-stressed pdl fibroblasts are mediated by h3 lysine acetylation associated with altered il10 expression. BioRxiv 2022. [Google Scholar] [CrossRef]
- Huynh, N.C.; Everts, V.; Pavasant, P.; Ampornaramveth, R.S. Inhibition of Histone Deacetylases Enhances the Osteogenic Differentiation of Human Periodontal Ligament Cells. J. Cell. Biochem. 2015, 117, 1384–1395. [Google Scholar] [CrossRef]
- Rodrigues, C.P.; Chatterjee, A.; Wiese, M.; Stehle, T.; Szymanski, W.; Shvedunova, M.; Akhtar, A. Histone H4 lysine 16 acetylation controls central carbon metabolism and diet-induced obesity in mice. Nat. Commun. 2021, 12, 6212. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Zhang, Z.; Li, Y.; Li, Q.; Geng, F.; Liu, J.; Pan, Y. Analysis of differentially expressed genes in oral epithelial cells infected with Fusobacterium nucleatum for revealing genes associated with oral cancer. J. Cell. Mol. Med. 2020, 25, 892–904. [Google Scholar] [CrossRef]
- Meaney, S. Epigenetic regulation of cholesterol homeostasis. Front. Genet. 2014, 5, 311. [Google Scholar] [CrossRef] [Green Version]
- Gry, M.; Rimini, R.; Strömberg, S.; Asplund, A.; Pontén, F.; Uhlén, M.; Nilsson, P. Correlations between RNA and protein expression profiles in 23 human cell lines. BMC Genom. 2009, 10, 365. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Sun, X.; Tang, Y.; Qu, Y.; Zhou, Y.; Zhang, Y. EZH2 reduction is an essential mechanoresponse for the maintenance of super-enhancer polarization against compressive stress in human periodontal ligament stem cells. Cell Death Dis. 2020, 11, 757. [Google Scholar] [CrossRef] [PubMed]
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Li, G. Post-translational modifications of PRC2: Signals directing its activity. Epigenetics Chromatin 2020, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Konze, K.D.; Ma, A.; Li, F.; Barsyte-Lovejoy, D.; Parton, T.; MacNevin°, C.J.; Liu, F.; Gao, C.; Huang, X.-P.; Kuznetsova, E.; et al. An Orally Bioavailable Chemical Probe of the Lysine Methyltransferases EZH2 and EZH1. ACS Chem. Biol. 2013, 8, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Grinshtein, N.; Rioseco, C.C.; Marcellus, R.; Uehling, D.; Aman, A.; Lun, X.; Muto, O.; Podmore, L.; Lever, J.; Shen, Y.; et al. Small molecule epigenetic screen identifies novel EZH2 and HDAC inhibitors that target glioblastoma brain tumor-initiating cells. Oncotarget 2016, 7, 59360–59376. [Google Scholar] [CrossRef] [PubMed]
- Katona, B.; Liu, Y.; Ma, A.; Jin, J.; Hua, X. EZH2 inhibition enhances the efficacy of an EGFR inhibitor in suppressing colon cancer cells. Cancer Biol. Ther. 2014, 15, 1677–1687. [Google Scholar] [CrossRef] [Green Version]
- Rankovic, M.J.; Docheva, D.; Wichelhaus, A.; Baumert, U. Effect of static compressive force on in vitro cultured PDL fibroblasts: Monitoring of viability and gene expression over 6 days. Clin. Oral Investig. 2019, 24, 2497–2511. [Google Scholar] [CrossRef]
- Brockhaus, J.; Craveiro, R.; Azraq, I.; Niederau, C.; Schröder, S.; Weiskirchen, R.; Jankowski, J.; Wolf, M. In Vitro Compression Model for Orthodontic Tooth Movement Modulates Human Periodontal Ligament Fibroblast Proliferation, Apoptosis and Cell Cycle. Biomolecules 2021, 11, 932. [Google Scholar] [CrossRef] [PubMed]
- Panpradit, N.; Nilmoje, T.; Kasetsuwan, J.; Sangkhamanee, S.S.; Surarit, R. Effect of Nicotine and Porphyromonas gingivalis on the Differentiation Properties of Periodontal Ligament Fibroblasts. Eur. J. Dent. 2021, 15, 727–732. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.J.; Zhang, J.; Lauricella, D.M.; Moore, S. IL-10 Is a Central Regulator of Cyclooxygenase-2 Expression and Prostaglandin Production. J. Immunol. 2001, 166, 2674–2680. [Google Scholar] [CrossRef] [Green Version]
- Jung, B.C.; Kang, S. Epigenetic regulation of inflammatory factors in adipose tissue. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 159019. [Google Scholar] [CrossRef]
- Le, H.Q.; Ghatak, S.; Yeung, C.-Y.C.; Tellkamp, F.; Günschmann, C.; Dieterich, C.; Yeroslaviz, A.; Habermann, B.; Pombo, A.; Niessen, C.M.; et al. Mechanical regulation of transcription controls Polycomb-mediated gene silencing during lineage commitment. Nat. Cell Biol. 2016, 18, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Thangavelu, S.R.; Merciaro, I.; D’Orazio, M.; Bramanti, P.; Mazzon, E.; Trubiani, O. Porphyromonas gingivalis lipopolysaccharide stimulation in human periodontal ligament stem cells: Role of epigenetic modifications to the inflammation. Eur. J. Histochem. 2017, 61, 2826. [Google Scholar] [CrossRef] [Green Version]
- Schwenk, R.W.; Holloway, G.P.; Luiken, J.J.; Bonen, A.; Glatz, J.F. Fatty acid transport across the cell membrane: Regulation by fatty acid transporters. Prostaglandins Leukot. Essent. Fat. Acids 2010, 82, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Kamp, F.; Hamilton, J. How fatty acids of different chain length enter and leave cells by free diffusion. Prostaglandins Leukot. Essent. Fat. Acids 2006, 75, 149–159. [Google Scholar] [CrossRef]
- Dimri, M.; Bommi, P.; Sahasrabuddhe, A.; Khandekar, J.D.; Dimri, G.P. Dietary omega-3 polyunsaturated fatty acids suppress expression of EZH2 in breast cancer cells. Carcinogenesis 2009, 31, 489–495. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Shahir, A.-M.; Sha, J.; Feng, Z.; Eapen, B.; Nithianantham, S.; Das, B.; Karn, J.; Weinberg, A.; Bissada, N.F.; et al. Short-Chain Fatty Acids from Periodontal Pathogens Suppress Histone Deacetylases, EZH2, and SUV39H1 To Promote Kaposi’s Sarcoma-Associated Herpesvirus Replication. J. Virol. 2014, 88, 4466–4479. [Google Scholar] [CrossRef] [Green Version]
- Ramazi, S.; Zahiri, J. Posttranslational modifications in proteins: Resources, tools and prediction methods. Database 2021, 2021, baab012. [Google Scholar] [CrossRef]
- Davis, F.M.; Dendekker, A.; Joshi, A.D.; Wolf, S.J.; Audu, C.; Melvin, W.J.; Mangum, K.; O Riordan, M.; Kunkel, S.L.; A Gallagher, K. Palmitate-TLR4 signaling regulates the histone demethylase, JMJD3, in macrophages and impairs diabetic wound healing. Eur. J. Immunol. 2020, 50, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Arisaka, M.; Yamashiro, Y. Fatty acid and prostaglandin metabolism in children with diabetes mellitus. II—The effect of evening primrose oil supplementation on serum fatty acid and plasma prostaglandin levels. Prostaglandins Leukot. Essent. Fat. Acids 1991, 43, 197–201. [Google Scholar] [CrossRef]
- Coll, T.; Palomer, X.; Blanco-Vaca, F.; Escolà-Gil, J.C.; Sánchez, R.M.; Laguna, J.C.; Vázquez-Carrera, M. Cyclooxygenase 2 Inhibition Exacerbates Palmitate-Induced Inflammation and Insulin Resistance in Skeletal Muscle Cells. Endocrinology 2010, 151, 537–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.K.; White, E.S.; Wettlaufer, S.H.; Grifka, H.; Hogaboam, C.M.; Thannickal, V.J.; Horowitz, J.C.; Peters-Golden, M. Prostaglandin E2 induces fibroblast apoptosis by modulating multiple survival pathways. FASEB J. 2009, 23, 4317–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifi, M.; Eslami, B.; Saffar, A.S. The effect of prostaglandin E2 and calcium gluconate on orthodontic tooth movement and root resorption in rats. Eur. J. Orthod. 2003, 25, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Saito, M.; Saito, S.; Ngan, P.W.; Shanfeld, J.; Davidovitch, Z. Interleukin 1 beta and prostaglandin E are involved in the response of periodontal cells to mechanical stress in vivo and in vitro. Am. J. Orthod. Dentofac. Orthop. 1991, 99, 226–240. [Google Scholar] [CrossRef]
- Hageman, J.; McCulloch, K.; Hunt, C.; Cobb, M.; Quade, B.; Crussi, F.; Pachman, L. Oleic acid lung injury increases plasma prostaglandin levels. Prostaglandins Leukot. Essent. Fat. Acids 1989, 35, 157–164. [Google Scholar] [CrossRef]
- Hollebeeck, S.; Raas, T.; Piront, N.; Schneider, Y.-J.; Toussaint, O.; Larondelle, Y.; During, A. Dimethyl sulfoxide (DMSO) attenuates the inflammatory response in the in vitro intestinal Caco-2 cell model. Toxicol. Lett. 2011, 206, 268–275. [Google Scholar] [CrossRef]
- Coward, W.R.; Feghali-Bostwick, C.A.; Jenkins, G.; Knox, A.J.; Pang, L. A central role for G9a and EZH2 in the epigenetic silencing of cyclooxygenase-2 in idiopathic pulmonary fibrosis. FASEB J. 2014, 28, 3183–3196. [Google Scholar] [CrossRef]
- Coward, W.R.; Watts, K.; Feghali-Bostwick, C.A.; Knox, A.; Pang, L. Defective Histone Acetylation Is Responsible for the Diminished Expression of Cyclooxygenase 2 in Idiopathic Pulmonary Fibrosis. Mol. Cell. Biol. 2009, 29, 4325–4339. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, M.; Hori, M.; Fujikawa, D.; Ohsugi, T.; Honma, D.; Adachi, N.; Katano, H.; Hishima, T.; Kobayashi, S.; Nakano, K.; et al. Targeting Excessive EZH1 and EZH2 Activities for Abnormal Histone Methylation and Transcription Network in Malignant Lymphomas. Cell Rep. 2019, 29, 2321–2337.e7. [Google Scholar] [CrossRef]
- Bradley, R.L.; Fisher, F.M.; Maratos-Flier, E. Dietary Fatty Acids Differentially Regulate Production of TNF-α and IL-10 by Murine 3T3-L1 Adipocytes. Obesity 2008, 16, 938–944. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kinkel, S.; Maksimovic, J.; Bandala-Sanchez, E.; Tanzer, M.C.; Naselli, G.; Zhang, J.G.; Zhan, Y.; Lew, A.M.; Silke, J.; et al. The polycomb repressive complex 2 governs life and death of peripheral T cells. Blood 2014, 124, 737–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobar, T.M.; Kanellopoulou, C.; Kugler, D.; Kilaru, G.; Nguyen, C.; Nagarajan, V.; Bhairavabhotla, R.K.; Northrup, D.; Zahr, R.; Burr, P.; et al. miR-155 Activates Cytokine Gene Expression in Th17 Cells by Regulating the DNA-Binding Protein Jarid2 to Relieve Polycomb-Mediated Repression. Immunity 2014, 40, 865–879. [Google Scholar] [CrossRef] [Green Version]
- Pande, V. Understanding the Complexity of Epigenetic Target Space. J. Med. Chem. 2016, 59, 1299–1307. [Google Scholar] [CrossRef]
- Bierman, E.L.; Dole, V.P.; Roberts, T.N. An Abnormality of Nonesterified Fatty Acid Metabolism in Diabetes Mellitus. Diabetes 1957, 6, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Taskinen, M.-R.; Bogardus, C.; Kennedy, A.; Howard, B.V. Multiple disturbances of free fatty acid metabolism in noninsulin-dependent diabetes. Effect of oral hypoglycemic therapy. J. Clin. Investig. 1985, 76, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Soares, E.A.; Nakagaki, W.; Garcia, J.A.D.; Camilli, J.A. Effect of hyperlipidemia on femoral biomechanics and morphology in low-density lipoprotein receptor gene knockout mice. J. Bone Miner. Metab. 2012, 30, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Kissebah, A.H.; Alfarsi, S.; Adams, P.W.; Wynn, V. Role of insulin resistance in adipose tissue and liver in the pathogenesis of endogenous hypertriglyceridaemia in man. Diabetologia 1976, 12, 563–571. [Google Scholar] [CrossRef]
- Cistola, D.; Small, D.M. Fatty acid distribution in systems modeling the normal and diabetic human circulation. A 13C nuclear magnetic resonance study. J. Clin. Investig. 1991, 87, 1431–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, M.; Tan, L.; Huang, S.; Zhao, L.; Tang, T.; Liu, J.; Zhao, Z. Analysis of time-course gene expression profiles of a periodontal ligament tissue model under compression. Arch. Oral Biol. 2013, 58, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, H.; Chiba, M.; Shimizu, Y.; Mitani, H. Periodontal ligament cells under mechanical stress induce osteoclastogenesis by receptor activator of nuclear factor kappab ligand up-regulation via prostaglandin e2 synthesis. J. Bone Miner Res. 2002, 17, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Cantley, M.D.; Bartold, P.M.; Marino, V.; Fairlie, D.P.; Le, G.T.; Lucke, A.J.; Haynes, D.R. Histone deacetylase inhibitors and periodontal bone loss. J. Periodontal Res. 2011, 46, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Pang, X.; Li, Z.; Chen, Z.; Wang, Y. Immunomodulation in the Treatment of Periodontitis: Progress and Perspectives. Front. Immunol. 2021, 12, 781378. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Symbol | NCBI Gene ID | Primer Sequence | Length |
|---|---|---|---|---|
| CREB-binding protein | CREBBP | 1387 | fw: CCAAGACCTGCGATTTCCAC rev: TTTTGATGTCCCAGAAGCGG | 100 bp |
| Embryonic ectoderm development | EED | 8726 | fw: TGCGATGGTTAGGCGATTTG rev: CCAAATGTCACACTGGCTGT | 158 bp |
| E1A-binding protein p300 | EP300 | 2033 | fw: TGACCAAGGGAGACAGCAAA rev: GAGGCGGATCACAAAGAAGAC | 182 bp |
| Enhancer of zeste 1 PRC2 subunit | EZH1 | 2145 | fw: CGAGAATGTGACCCTGACCT rev: TTATGAAGGTGCCCCATCCG | 154 bp |
| Enhancer of zeste 2 PRC2 subunit | EZH2 | 2146 | fw: ACAGTTCGTGCCCTTGTGTG rev: CACTCTCGGACAGCCAGGTA | 148 bp |
| Histone deacetylase 1 | HDAC1 | 3065 | fw: AGCTCCACATCAGTCCTTCCA rev: TTCGTCCTCATCGCCACTCT | 170 bp |
| Histone deacetylase 2 | HDAC2 | 3066 | fw: ACTGATGCTTGGAGGAGGTG rev: CTGGAGTGTTCTGGTTTGTCA | 185 bp |
| Histone deacetylase 3 | HDAC3 | 8841 | fw: GCTGGGTGGTGGTGGTTATA rev: TTCTGATTCTCGATGCGGGT | 174 bp |
| Interleukin 10 | IL10 | 3586 | fw: AGCCATGAGTGAGTTTGACA rev: AGAGCCCCAGATCCGATTTT | 141 bp |
| Lysine acetyltransferase 8 | KAT8 | 84148 | fw: GCAAGATCACTCGCAACCAA rev: AGTCTTCGGGGAATGGTGAG | 195 bp |
| Nuclear receptor coactivator 3 | NCOA3 | 8202 | fw: GGCTCTATTCCCACATTGCC rev: CCCAGTTGGTTAGATGCTGC | 158 bp |
| Prostaglandin endoperoxide synthase 2 | PTGS2 (COX2) | 5743 | fw: GATGATTGCCCGACTCCCTT rev: GGCCCTCGCTTATGATCTGT | 185 bp |
| Ribosomal protein L22 | RPL22 | 6146 | fw: TGATTGCACCCACCCTGTAG rev: GGTTCCCAGCTTTTCCGTTC | 98 bp |
| SIN3 transcription regulator family member A | SIN3A | 25942 | fw: GAGCAGCAGGAAAAGGAAGG rev: TGTAGACGCTTGCTTACACG | 200 bp |
| SUZ12 polycomb repressive complex 2 subunit | SUZ12 | 23512 | fw: CGGACCAGTTAAGAGAACACC rev TTGTGGACGGAGAGGTAAGC | 181 bp |
| TATA box-binding protein | TBP | 6908 | fw: CGGCTGTTTAACTTCGCTTCC rev: TGGGTTATCTTCACACGCCAAG | 86 bp |
| Location | Label | Primer Sequence | Length |
|---|---|---|---|
| Promoter region | #1 | fw: TGAAGAAGTCCTGATGTCAC rev: TTACCTATCCCTACTTCCCC | 187 bp |
| Promoter region | #2 | fw: AGCACTACCTGACTAGCATA rev: AGAGACTGGCTTCCTACAG | 192 bp |
| Promoter region | #3 | fw: GGGGACCCAATTATTTCTCA rev: TGGGCTACCTCTCTTAGAAT | 188 bp |
| Non-promoter region | #4 | fw: GCTTAGAGCGTTTCCAGACC rev: CTCCCCACTGTAGACATCCA | 131 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuldt, L.; Reimann, M.; von Brandenstein, K.; Steinmetz, J.; Döding, A.; Schulze-Späte, U.; Jacobs, C.; Symmank, J. Palmitate-Triggered COX2/PGE2-Related Hyperinflammation in Dual-Stressed PdL Fibroblasts Is Mediated by Repressive H3K27 Trimethylation. Cells 2022, 11, 955. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060955
Schuldt L, Reimann M, von Brandenstein K, Steinmetz J, Döding A, Schulze-Späte U, Jacobs C, Symmank J. Palmitate-Triggered COX2/PGE2-Related Hyperinflammation in Dual-Stressed PdL Fibroblasts Is Mediated by Repressive H3K27 Trimethylation. Cells. 2022; 11(6):955. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060955
Chicago/Turabian StyleSchuldt, Lisa, Michael Reimann, Katrin von Brandenstein, Julia Steinmetz, Annika Döding, Ulrike Schulze-Späte, Collin Jacobs, and Judit Symmank. 2022. "Palmitate-Triggered COX2/PGE2-Related Hyperinflammation in Dual-Stressed PdL Fibroblasts Is Mediated by Repressive H3K27 Trimethylation" Cells 11, no. 6: 955. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060955