
Key Genes and Biochemical Networks in Various Brain Regions Affected in Alzheimer’s Disease

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Dataset Selection
2.3. Integrated Genomic Analysis
2.4. Pathway Analysis
2.5. Protein–Protein Interaction Analysis
2.6. Gene–miRNA Interaction Analysis
2.7. Gene-Transcription Factors Interaction Analysis
3. Results
3.1. Search Results and Characteristics of Selected Studies

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Datasets | Country | Number of AD/CTR | Age (years) AD/CTR | Postmortem Interval (h) AD/CTR | Brain Region (s) | Reference |
|---|---|---|---|---|---|---|
| GSE118553 | UK | 52/27 | 82.9 ± 8.7/70.6 ± 15.9 | 39.9 ± 21.3/ 37.1 ± 20.7 | Cerebellum/Entorhinal/Frontal/Temporal | [26] |
| GSE44768 | USA | 129/101 | - | - | Cerebellum | [27] |
| GSE48350 | USA | 15/39 | 85.7 ± 6.3/64.8 ± 9.5 | - | Entorhinal/Frontal/Hippocampus | [28] |
| GSE5281 | USA | 33/14 | 79.9 ± 6.9/79.8 ± 9.1 | 2.5/2.5 | Entorhinal/Frontal/Temporal/Hippocampus | [29,30] |
| GSE33000 | USA | 310/157 | 80.6 ± 9.0/63.5 ± 9.9 | 13.7 ± 7.4/22.4 ± 5.8 | Frontal | [31] |
| GSE44770 | USA | 129/101 | - | - | Frontal | [27] |
| GSE36980 | Japan | 26/62 | 83.0 ± 5.7 | - | Frontal/Temporal/Hippocampus | [32,33] |
| GSE122063 | USA | 12/11 | 80.9 ± 7.4/78.6 ± 8.5 | 8.0 ± 4.0/ 9.0 ± 3.0 | Frontal/Temporal | [34] |
| GSE132903 | USA | 97/98 | 85.02 ± 6.75/84.98 ± 6.90 | - | Temporal | [35] |
| GSE29378 | USA | 17/16 | 77.3 ± 9.1/81.7 ± 6.9 | 11.2 ± 6.3/10.8 ± 6.8 | Hippocampus | [36] |
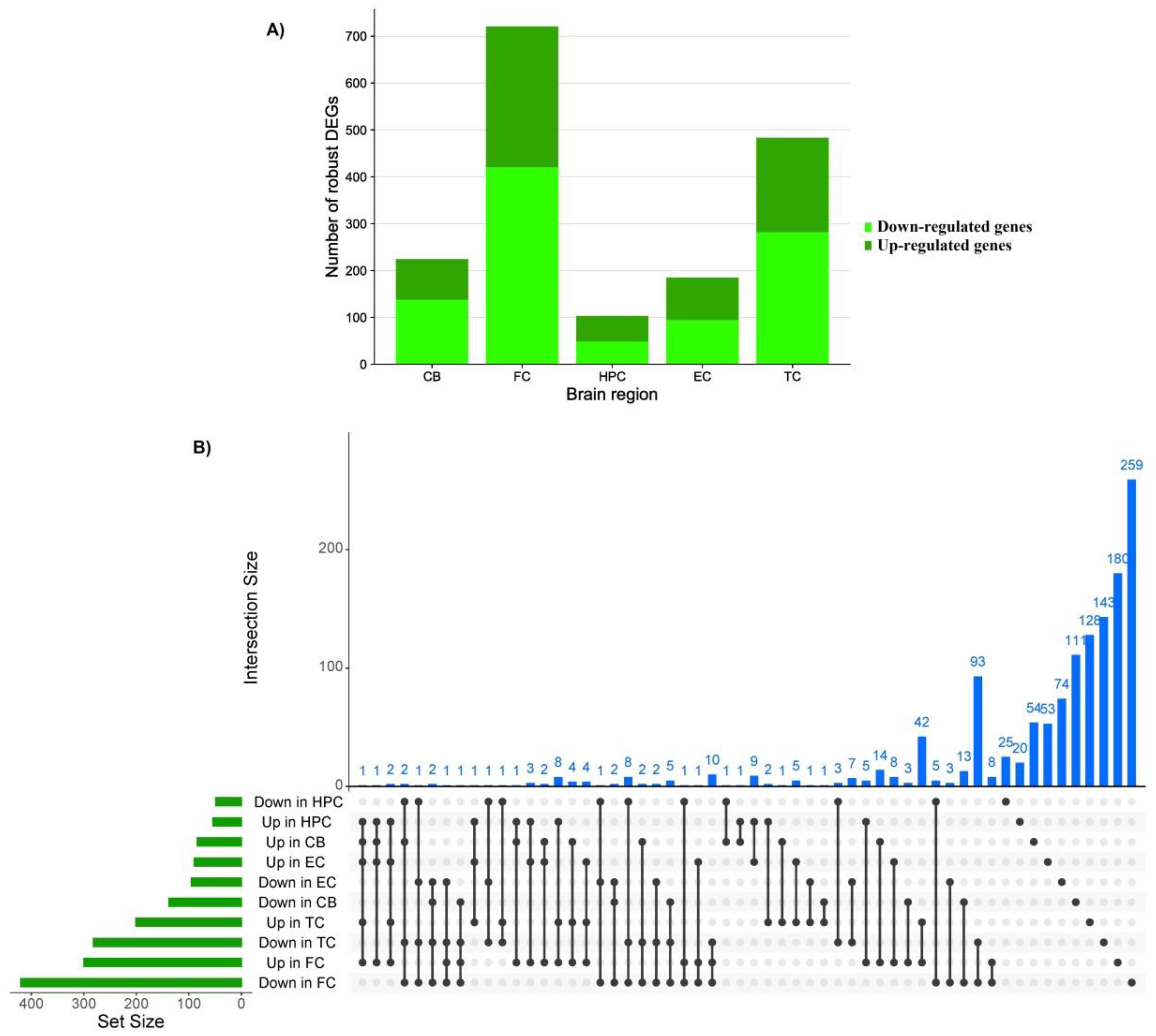
3.2. Robust Differentially Expressed Genes
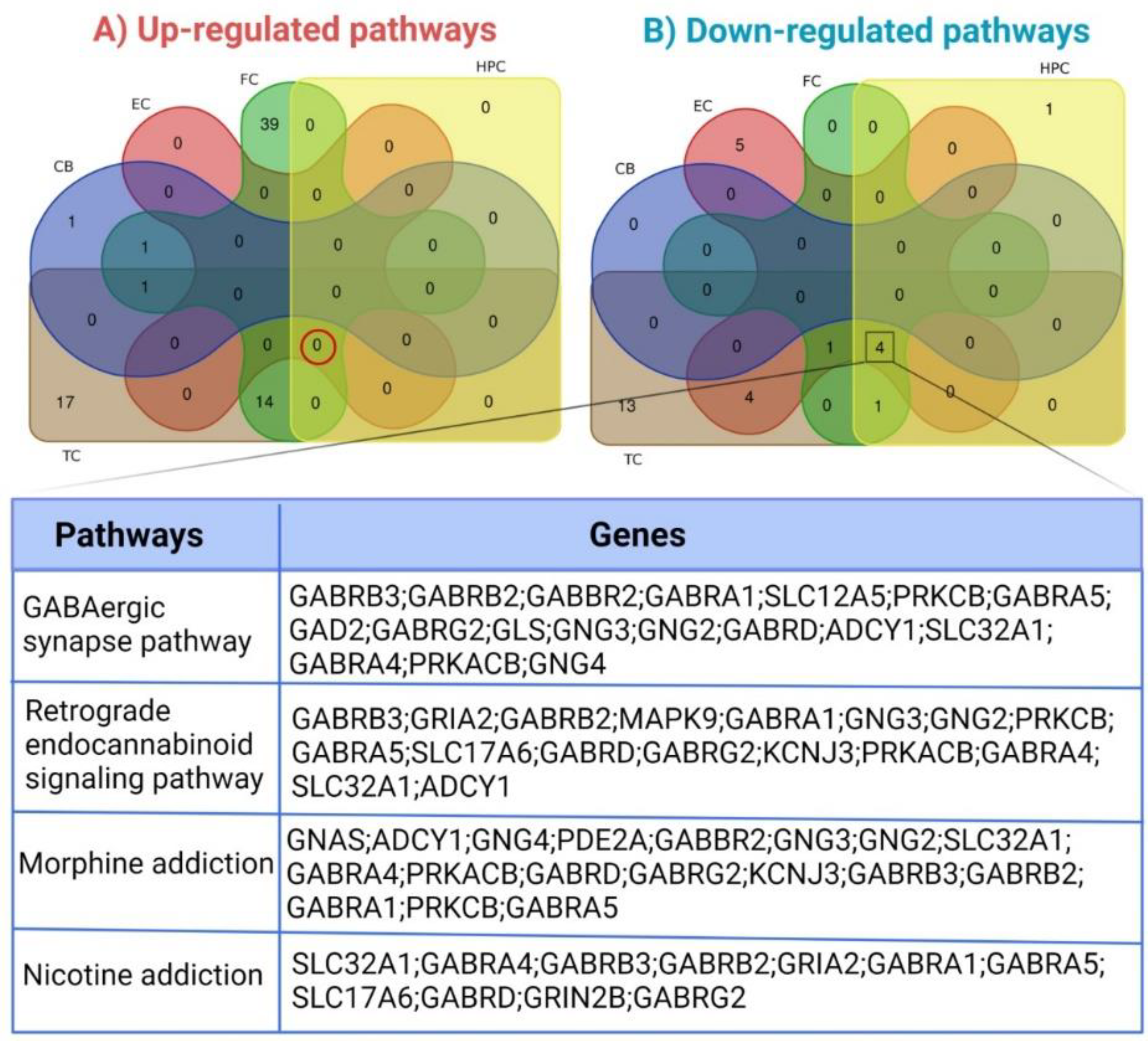
3.3. Pathways Enriched by Differentially Expressed Genes
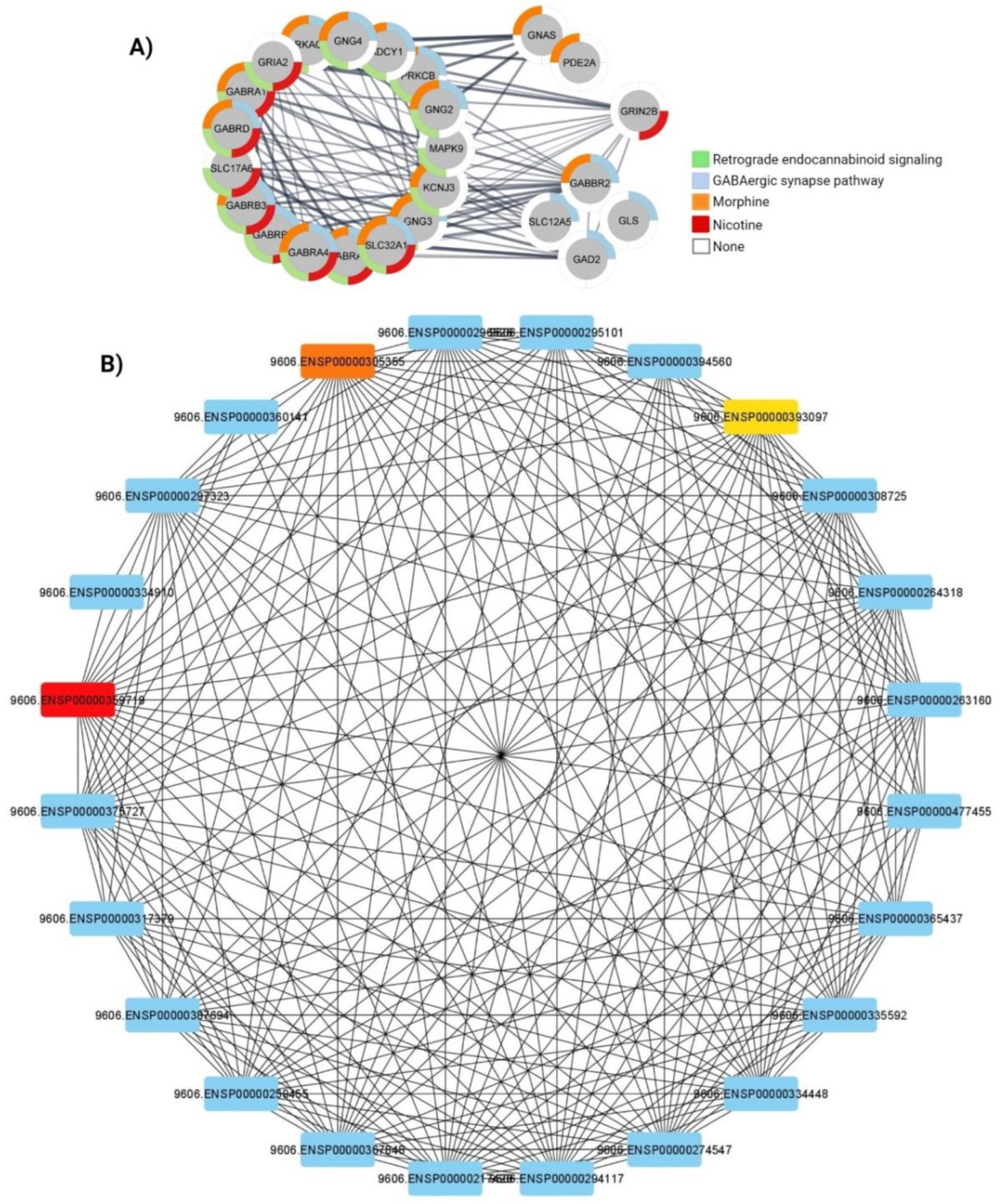
3.4. Protein-Protein Interaction Analysis
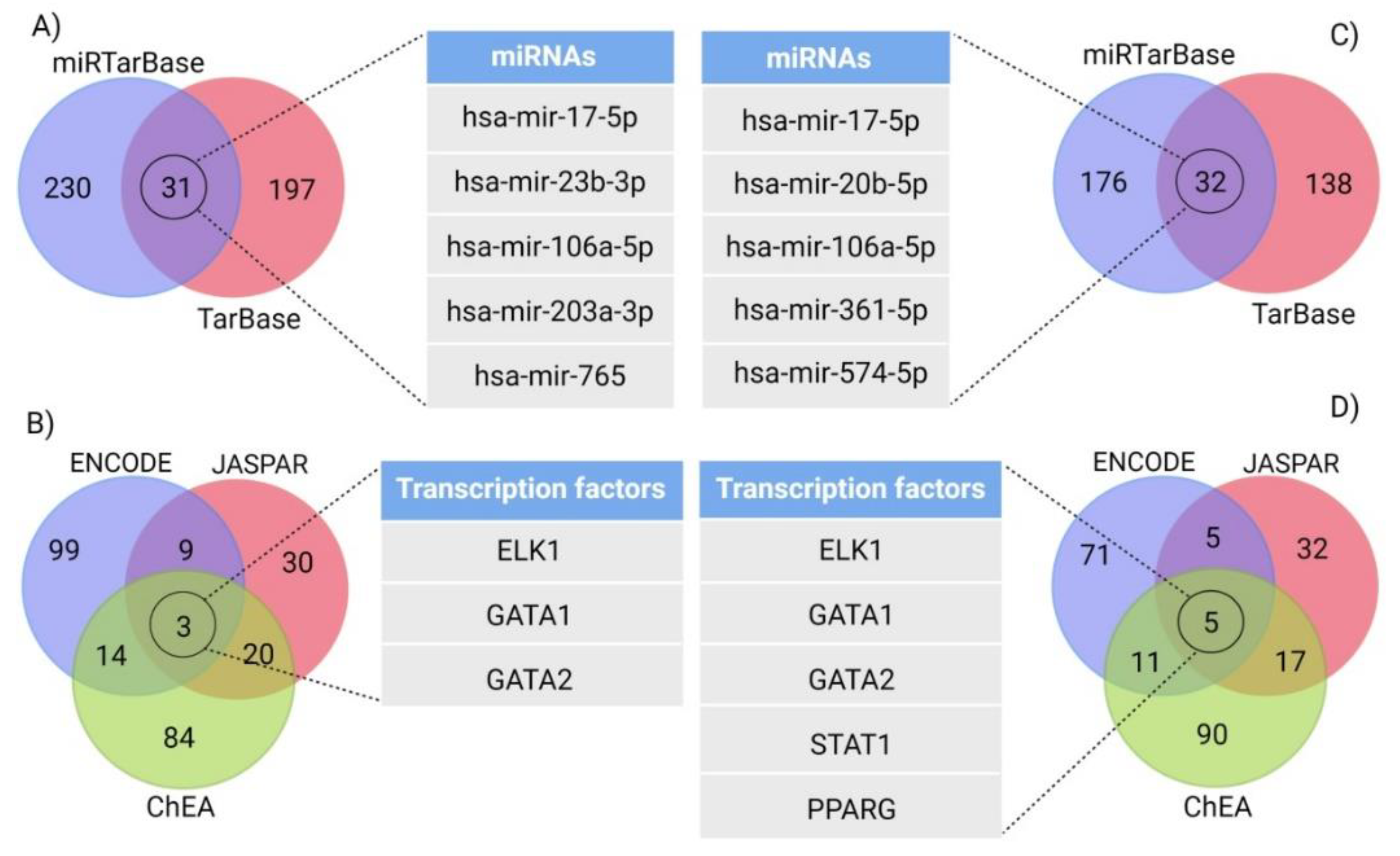
3.5. Gene–miRNA Interaction and Targeting miRNA Analysis
3.6. Gene–Transcription Factors Interaction Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Abyadeh, M.; Gupta, V.; Gupta, V.; Chitranshi, N.; Wu, Y.; Amirkhani, A.; Meyfour, A.; Sheriff, S.; Shen, T.; Dhiman, K. Comparative Analysis of Aducanumab, Zagotenemab and Pioglitazone as Targeted Treatment Strategies for Alzheimer’s Disease. Aging Dis. 2021, 12, 1964. [Google Scholar] [CrossRef] [PubMed]
- Abyadeh, M.; Djafarian, K.; Heydarinejad, F.; Alizadeh, S.; Shab-Bidar, S. Association between apolipoprotein E gene polymorphism and Alzheimer’s disease in an Iranian population: A meta-analysis. J. Mol. Neurosci. 2019, 69, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Abyadeh, M.; Gupta, V.; Chitranshi, N.; Gupta, V.; Wu, Y.; Saks, D.; Wander Wall, R.; Fitzhenry, M.J.; Basavarajappa, D.; You, Y. Mitochondrial dysfunction in Alzheimer’s disease-a proteomics perspective. Expert Rev. Proteom. 2021, 18, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Long, J.M.; Holtzman, D.M. Alzheimer disease: An update on pathobiology and treatment strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef]
- Chitranshi, N.; Kumar, A.; Sheriff, S.; Gupta, V.; Godinez, A.; Saks, D.; Sarkar, S.; Shen, T.; Mirzaei, M.; Basavarajappa, D. Identification of novel cathepsin B inhibitors with implications in Alzheimer’s disease: Computational refining and biochemical evaluation. Cells 2021, 10, 1946. [Google Scholar] [CrossRef]
- Mirzaei, M.; Deng, L.; Gupta, V.B.; Graham, S.; Gupta, V. Complement pathway in Alzheimer’s pathology and retinal neurodegenerative disorders–the road ahead. Neural Regen. Res. 2020, 15, 257. [Google Scholar] [CrossRef]
- Joseph, C.; Mangani, A.S.; Gupta, V.; Chitranshi, N.; Shen, T.; Dheer, Y.; Kb, D.; Mirzaei, M.; You, Y.; Graham, S.L. Cell cycle deficits in neurodegenerative disorders: Uncovering molecular mechanisms to drive innovative therapeutic development. Aging Dis. 2020, 11, 946. [Google Scholar] [CrossRef]
- Vosoughi, A.; Sadigh-Eteghad, S.; Ghorbani, M.; Shahmorad, S.; Farhoudi, M.; Rafi, M.A.; Omidi, Y. Mathematical models to shed light on amyloid-beta and tau protein dependent pathologies in Alzheimer’s disease. Neuroscience 2020, 424, 45–57. [Google Scholar] [CrossRef]
- Hampel, H.; Nisticò, R.; Seyfried, N.T.; Levey, A.I.; Modeste, E.; Lemercier, P.; Baldacci, F.; Toschi, N.; Garaci, F.; Perry, G. Omics Sciences for Systems Biology in Alzheimer’s Disease: State-of-the-Art of the Evidence. Ageing Res. Rev. 2021, 69, 101346. [Google Scholar] [CrossRef]
- Deng, L.; Gupta, V.K.; Wu, Y.; Pushpitha, K.; Chitranshi, N.; Gupta, V.B.; Fitzhenry, M.J.; Moghaddam, M.Z.; Karl, T.; Salekdeh, G.H. Mouse model of Alzheimer’s disease demonstrates differential effects of early disease pathology on various brain regions. Proteomics 2021, 21, 2000213. [Google Scholar] [CrossRef]
- Deng, L.; Haynes, P.A.; Wu, Y.; Amirkhani, A.; Kamath, K.S.; Wu, J.X.; Pushpitha, K.; Gupta, V.; Graham, S.; Gupta, V.K. Amyloid-beta peptide neurotoxicity in human neuronal cells is associated with modulation of insulin-like growth factor transport, lysosomal machinery and extracellular matrix receptor interactions. Neural Regen. Res. 2020, 15, 2131. [Google Scholar] [PubMed]
- Hosseinian, S.; Arefian, E.; Rakhsh-Khorshid, H.; Eivani, M.; Rezayof, A.; Pezeshk, H.; Marashi, S.-A. A meta-analysis of gene expression data highlights synaptic dysfunction in the hippocampus of brains with Alzheimer’s disease. Sci. Rep. 2020, 10, 8384. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, H. Integrated Bioinformatics Analysis to Identify Alternative Therapeutic Targets for Alzheimer’s Disease: Insights from a Synaptic Machinery Perspective. J. Mol. Neurosci. 2021, 72, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Long, J.; He, T.; Belshaw, R.; Scott, J. Integrated genomic approaches identify major pathways and upstream regulators in late onset Alzheimer’s disease. Sci. Rep. 2015, 5, 12393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noori, A.; Mezlini, A.M.; Hyman, B.T.; Serrano-Pozo, A.; Das, S. Systematic review and meta-analysis of human transcriptomics reveals neuroinflammation, deficient energy metabolism, and proteostasis failure across neurodegeneration. Neurobiol. Dis. 2021, 149, 105225. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; Ding, F.; Zhao, Y. Integrated identification of key genes and pathways in Alzheimer’s disease via comprehensive bioinformatical analyses. Hereditas 2019, 156, 25. [Google Scholar] [CrossRef]
- Guennewig, B.; Lim, J.; Marshall, L.; McCorkindale, A.N.; Paasila, P.J.; Patrick, E.; Kril, J.J.; Halliday, G.M.; Cooper, A.A.; Sutherland, G.T. Defining early changes in Alzheimer’s disease from RNA sequencing of brain regions differentially affected by pathology. Sci. Rep. 2021, 11, 4865. [Google Scholar] [CrossRef]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M. Gene set knowledge discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Deng, L.; Pushpitha, K.; Joseph, C.; Gupta, V.; Rajput, R.; Chitranshi, N.; Dheer, Y.; Amirkhani, A.; Kamath, K.; Pascovici, D. Amyloid β induces early changes in the ribosomal machinery, cytoskeletal organization and oxidative phosphorylation in retinal photoreceptor cells. Front. Mol. Neurosci. 2019, 12, 24. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [Green Version]
- Karagkouni, D.; Paraskevopoulou, M.D.; Chatzopoulos, S.; Vlachos, I.S.; Tastsoglou, S.; Kanellos, I.; Papadimitriou, D.; Kavakiotis, I.; Maniou, S.; Skoufos, G. DIANA-TarBase v8: A decade-long collection of experimentally supported miRNA–gene interactions. Nucleic Acids Res. 2018, 46, D239–D245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, C.-H.; Shrestha, S.; Yang, C.-D.; Chang, N.-W.; Lin, Y.-L.; Liao, K.-W.; Huang, W.-C.; Sun, T.-H.; Tu, S.-J.; Lee, W.-H. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef] [PubMed]
- Consortium, E.P. A user’s guide to the encyclopedia of DNA elements (ENCODE). PLoS Biol. 2011, 9, e1001046. [Google Scholar]
- Lachmann, A.; Xu, H.; Krishnan, J.; Berger, S.I.; Mazloom, A.R.; Ma’ayan, A. ChEA: Transcription factor regulation inferred from integrating genome-wide ChIP-X experiments. Bioinformatics 2010, 26, 2438–2444. [Google Scholar] [CrossRef]
- Fornes, O.; Castro-Mondragon, J.A.; Khan, A.; Van der Lee, R.; Zhang, X.; Richmond, P.A.; Modi, B.P.; Correard, S.; Gheorghe, M.; Baranašić, D. JASPAR 2020: Update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2020, 48, D87–D92. [Google Scholar] [CrossRef]
- Patel, H.; Hodges, A.K.; Curtis, C.; Lee, S.H.; Troakes, C.; Dobson, R.J.; Newhouse, S.J. Transcriptomic analysis of probable asymptomatic and symptomatic alzheimer brains. Brain Behav. Immun. 2019, 80, 644–656. [Google Scholar] [CrossRef]
- Zhang, B.; Gaiteri, C.; Bodea, L.-G.; Wang, Z.; McElwee, J.; Podtelezhnikov, A.A.; Zhang, C.; Xie, T.; Tran, L.; Dobrin, R. Integrated systems approach identifies genetic nodes and networks in late-onset Alzheimer’s disease. Cell 2013, 153, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Berchtold, N.C.; Coleman, P.D.; Cribbs, D.H.; Rogers, J.; Gillen, D.L.; Cotman, C.W. Synaptic genes are extensively downregulated across multiple brain regions in normal human aging and Alzheimer’s disease. Neurobiol. Aging 2013, 34, 1653–1661. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.S.; Dunckley, T.; Beach, T.G.; Grover, A.; Mastroeni, D.; Ramsey, K.; Caselli, R.J.; Kukull, W.A.; McKeel, D.; Morris, J.C. Altered neuronal gene expression in brain regions differentially affected by Alzheimer’s disease: A reference data set. Physiol. Genom. 2008, 33, 240–256. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.S.; Dunckley, T.; Beach, T.G.; Grover, A.; Mastroeni, D.; Walker, D.G.; Caselli, R.J.; Kukull, W.A.; McKeel, D.; Morris, J.C. Gene expression profiles in anatomically and functionally distinct regions of the normal aged human brain. Physiol. Genom. 2007, 28, 311–322. [Google Scholar] [CrossRef]
- Narayanan, M.; Huynh, J.L.; Wang, K.; Yang, X.; Yoo, S.; McElwee, J.; Zhang, B.; Zhang, C.; Lamb, J.R.; Xie, T. Common dysregulation network in the human prefrontal cortex underlies two neurodegenerative diseases. Mol. Syst. Biol. 2014, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Ninomiya, T.; Kubo, M.; Hirakawa, Y.; Doi, Y.; Hata, J.; Iwaki, T.; Kanba, S.; Kiyohara, Y. Apolipoprotein genotype for prediction of Alzheimer’s disease in older Japanese: The Hisayama Study. J. Am. Geriatr. Soc. 2011, 59, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Hokama, M.; Oka, S.; Leon, J.; Ninomiya, T.; Honda, H.; Sasaki, K.; Iwaki, T.; Ohara, T.; Sasaki, T.; LaFerla, F.M. Altered expression of diabetes-related genes in Alzheimer’s disease brains: The Hisayama study. Cereb. Cortex 2014, 24, 2476–2488. [Google Scholar] [CrossRef] [PubMed]
- McKay, E.C.; Beck, J.S.; Khoo, S.K.; Dykema, K.J.; Cottingham, S.L.; Winn, M.E.; Paulson, H.L.; Lieberman, A.P.; Counts, S.E. Peri-infarct upregulation of the oxytocin receptor in vascular dementia. J. Neuropathol. Exp. Neurol. 2019, 78, 436–452. [Google Scholar] [CrossRef] [PubMed]
- Piras, I.S.; Krate, J.; Delvaux, E.; Nolz, J.; Mastroeni, D.F.; Persico, A.M.; Jepsen, W.M.; Beach, T.G.; Huentelman, M.J.; Coleman, P.D. Transcriptome changes in the Alzheimer’s disease middle temporal gyrus: Importance of RNA metabolism and mitochondria-associated membrane genes. J. Alzheimer’s Dis. 2019, 70, 691–713. [Google Scholar] [CrossRef]
- Miller, J.A.; Woltjer, R.L.; Goodenbour, J.M.; Horvath, S.; Geschwind, D.H. Genes and pathways underlying regional and cell type changes in Alzheimer’s disease. Genome Med. 2013, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Kolde, R.; Laur, S.; Adler, P.; Vilo, J. Robust rank aggregation for gene list integration and meta-analysis. Bioinformatics 2012, 28, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhao, M.; Han, Y.; Zhang, H. GABAergic inhibitory interneuron deficits in Alzheimer’s disease: Implications for treatment. Front. Neurosci. 2020, 14, 660. [Google Scholar] [CrossRef]
- Mizukami, K.; Ikonomovic, M.D.; Grayson, D.R.; Sheffield, R.; Armstrong, D.M. Immunohistochemical study of GABAA receptor α1 subunit in the hippocampal formation of aged brains with Alzheimer-related neuropathologic changes. Brain Res. 1998, 799, 148–155. [Google Scholar] [CrossRef]
- Rissman, R.; Mishizen-Eberz, A.; Carter, T.; Wolfe, B.; De Blas, A.; Miralles, C.; Ikonomovic, M.; Armstrong, D. Biochemical analysis of GABAA receptor subunits α1, α5, β1, β2 in the hippocampus of patients with Alzheimer’s disease neuropathology. Neuroscience 2003, 120, 695–704. [Google Scholar] [CrossRef]
- Iwakiri, M.; Mizukami, K.; Ikonomovic, M.D.; Ishikawa, M.; Abrahamson, E.E.; DeKosky, S.T.; Asada, T. An immunohistochemical study of GABAA receptor gamma subunits in Alzheimer’s disease hippocampus: Relationship to neurofibrillary tangle progression. Neuropathology 2009, 29, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizukami, K.; Grayson, D.R.; Ikonomovic, M.D.; Sheffield, R.; Armstrong, D.M. GABAA receptor β2 and β3 subunits mRNA in the hippocampal formation of aged human brain with Alzheimer-related neuropathology. Mol. Brain Res. 1998, 56, 268–272. [Google Scholar] [CrossRef]
- Li, Y.; Sun, H.; Chen, Z.; Xu, H.; Bu, G.; Zheng, H. Implications of GABAergic neurotransmission in Alzheimer’s disease. Front. Aging Neurosci. 2016, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.Q.; Yao, J.J.; Liu, D.D.; Ma, Q.; Mei, Y.A. Aβ40 modulates GABAA receptor α6 subunit expression and rat cerebellar granule neuron maturation through the ERK/mTOR pathway. J. Neurochem. 2014, 128, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Frere, S.; Slutsky, I. Alzheimer’s disease: From firing instability to homeostasis network collapse. Neuron 2018, 97, 32–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, L.M.; Djukic, B.; Arnold, C.; Gillespie, A.K.; Yoon, S.Y.; Wang, M.M.; Zhang, O.; Knoferle, J.; Rubenstein, J.L.; Alvarez-Buylla, A. Inhibitory interneuron progenitor transplantation restores normal learning and memory in ApoE4 knock-in mice without or with Aβ accumulation. J. Neurosci. 2014, 34, 9506–9515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, Z.; Tan, L.; Wang, Y.; Lu, C.; Chen, R.; Zhang, S.; Gao, Y.; Liu, Y.; Yin, Y. Correcting miR92a-vGAT-mediated GABAergic dysfunctions rescues human tau-induced anxiety in mice. Mol. Ther. 2017, 25, 140–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Li, H.-L.; Tian, N.; Liu, F.; Wang, L.; Yin, Y.; Yue, L.; Ma, L.; Wan, Y.; Wang, J.-Z. Interneuron accumulation of phosphorylated tau impairs adult hippocampal neurogenesis by suppressing GABAergic transmission. Cell Stem Cell 2020, 26, 331–345.e336. [Google Scholar] [CrossRef]
- Abyadeh, M.; Gupta, V.; Paulo, J.A.; Gupta, V.; Chitranshi, N.; Godinez, A.; Saks, D.; Hasan, M.; Amirkhani, A.; McKay, M. A proteomic view of cellular and molecular effects of cannabis. Biomolecules 2021, 11, 1411. [Google Scholar] [CrossRef]
- Lu, H.-C.; Mackie, K. An introduction to the endogenous cannabinoid system. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Cristino, L.; Bisogno, T.; Di Marzo, V. Cannabinoids and the expanded endocannabinoid system in neurological disorders. Nat. Rev. Neurol. 2020, 16, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Ohno-Shosaku, T.; Maejima, T.; Kano, M. Endogenous cannabinoids mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron 2001, 29, 729–738. [Google Scholar] [CrossRef]
- Wilson, R.I.; Nicoll, R.A. Endogenous cannabinoids mediate retrograde signalling at hippocampal synapses. Nature 2001, 410, 588–592. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Kumar, U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.-Y.; Chen, C. Endocannabinoids in synaptic plasticity and neuroprotection. Neuroscientist 2015, 21, 152–168. [Google Scholar] [CrossRef] [Green Version]
- Mulder, J.; Zilberter, M.; Pasquare, S.J.; Alpar, A.; Schulte, G.; Ferreira, S.G.; Köfalvi, A.; Martín-Moreno, A.M.; Keimpema, E.; Tanila, H. Molecular reorganization of endocannabinoid signalling in Alzheimer’s disease. Brain 2011, 134, 1041–1060. [Google Scholar] [CrossRef]
- Bottero, V.; Potashkin, J.A. Meta-analysis of gene expression changes in the blood of patients with mild cognitive impairment and Alzheimer’s disease dementia. Int. J. Mol. Sci. 2019, 20, 5403. [Google Scholar] [CrossRef]
- Ge, X.-Y.; Cui, K.; Liu, L.; Qin, Y.; Cui, J.; Han, H.-J.; Luo, Y.-H.; Yu, H.-M. Screening and predicting progression from high-risk mild cognitive impairment to Alzheimer’s disease. Sci. Rep. 2021, 11, 17558. [Google Scholar] [CrossRef]
- Estfanous, S.; Daily, K.P.; Eltobgy, M.; Deems, N.P.; Anne, M.N.; Krause, K.; Badr, A.; Hamilton, K.; Carafice, C.; Hegazi, A. Elevated Expression of MiR-17 in Microglia of Alzheimer’s Disease Patients Abrogates Autophagy-Mediated Amyloid-β Degradation. Front. Immunol. 2021, 12, 2839. [Google Scholar] [CrossRef]
- Yılmaz, Ş.G.; Erdal, M.E.; Özge, A.A.; Sungur, M.A. Can peripheral MicroRNA expression data serve as epigenomic (upstream) biomarkers of Alzheimer’s disease? Omics A J. Integr. Biol. 2016, 20, 456–461. [Google Scholar] [CrossRef]
- Ma, J.; Wang, W.; Azhati, B.; Wang, Y.; Tusong, H. miR-106a-5p Functions as a Tumor Suppressor by Targeting VEGFA in Renal Cell Carcinoma. Dis. Markers 2020, 2020, 8837941. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, E.R.; Dumitrescu, L.; Moore, A.M.; Cambronero, F.E.; De Jager, P.L.; Koran, M.E.I.; Petyuk, V.A.; Robinson, R.A.; Goyal, S.; Schneider, J.A. Brain expression of the vascular endothelial growth factor gene family in cognitive aging and alzheimer’s disease. Mol. Psychiatry 2021, 26, 888–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Li, X.; Zuo, Y.; Xu, Q.; Liu, L.; Wu, H.; Chen, L.; Zhang, Y.; Liu, Y.; Li, Y. miR-373-3p inhibits epithelial–mesenchymal transition via regulation of TGFβR2 in choriocarcinoma. J. Obstet. Gynaecol. Res. 2021, 47, 2417–2432. [Google Scholar] [CrossRef] [PubMed]
- Fessel, J. Ineffective levels of transforming growth factors and their receptor account for old age being a risk factor for Alzheimer’s disease. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2019, 5, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Callahan, L.M.; Sul, J.-Y.; Kim, T.K.; Barrett, L.; Kim, M.; Powers, J.M.; Federoff, H.; Eberwine, J. A neurotoxic phosphoform of Elk-1 associates with inclusions from multiple neurodegenerative diseases. PLoS ONE 2010, 5, e9002. [Google Scholar] [CrossRef]
- Galan, B.; Caboche, J. Elk-1 a transcription factor with multiple facets in the brain. Front. Neurosci. 2011, 5, 35. [Google Scholar]
- Tong, L.; Balazs, R.; Thornton, P.L.; Cotman, C.W. β-amyloid peptide at sublethal concentrations downregulates brain-derived neurotrophic factor functions in cultured cortical neurons. J. Neurosci. 2004, 24, 6799–6809. [Google Scholar] [CrossRef] [Green Version]
- Pastorcic, M.; Das, H.K. Ets transcription factors ER81 and Elk1 regulate the transcription of the human presenilin 1 gene promoter. Mol. Brain Res. 2003, 113, 57–66. [Google Scholar] [CrossRef]
- Chu, J.; Wisniewski, T.; Praticò, D. GATA 1-mediated transcriptional regulation of the γ-secretase activating protein increases A β formation in D own syndrome. Ann. Neurol. 2016, 79, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Roy, J.; Mallick, B. Altered gene expression in late-onset Alzheimer’s disease due to SNPs within 3′ UTR microRNA response elements. Genomics 2017, 109, 177–185. [Google Scholar] [CrossRef]
- Tam, K.T.; Chan, P.K.; Zhang, W.; Law, P.P.; Tian, Z.; Fung Chan, G.C.; Philipsen, S.; Festenstein, R.; Tan-Un, K.C. Identification of a novel distal regulatory element of the human Neuroglobin gene by the chromosome conformation capture approach. Nucleic Acids Res. 2017, 45, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vidania, S.; Palomares-Perez, I.; Frank-García, A.; Saito, T.; Saido, T.C.; Draffin, J.; Szaruga, M.; Chávez-Gutierrez, L.; Calero, M.; Medina, M. Prodromal Alzheimer’s Disease: Constitutive Upregulation of Neuroglobin Prevents the Initiation of Alzheimer’s Pathology. Front. Neurosci. 2020, 14, 1001. [Google Scholar] [CrossRef] [PubMed]
- Ciccone, L.; Nencetti, S.; Socci, S.; Orlandini, E. Neuroglobin and neuroprotection: The role of natural and synthetic compounds in neuroglobin pharmacological induction. Neural Regen. Res. 2021, 16, 2353. [Google Scholar] [PubMed]
- Bottero, V.; Powers, D.; Yalamanchi, A.; Quinn, J.P.; Potashkin, J.A. Key Disease Mechanisms Linked to Alzheimer’s Disease in the Entorhinal Cortex. Int. J. Mol. Sci. 2021, 22, 3915. [Google Scholar] [CrossRef]




| Brain Region | Pathway | Alteration | p-Value | Adjusted p-Value | Odd Ratio | Combined Score | Genes |
|---|---|---|---|---|---|---|---|
| Cerebellum | Mineral absorption | Up-regulated | 0.000005926 | 0.0007467 | 22.29 | 268.27 | MT2A, MT1A, MT1M, ATP2B4, MT1G |
| IL-17 signaling pathway | Up-regulated | 0.00005312 | 0.003347 | 13.75 | 135.34 | NFKBIA, CEBPB, CXCL1, S100A9, S100A8 | |
| NF-kappa B signaling pathway | Up-regulated | 0.01023 | 0.3936 | 7.09 | 32.49 | NFKBIA, GADD45A, CXCL1 | |
| Frontal cortex | Neuroactive ligand-receptor interaction | Down-regulated | 0.000001048 | 0.0002148 | 3.50 | 48.19 | GABBR2, GABRA1, CHRM3, EDN3, NPY5R, GABRA5, GABRA4, GRIK1, HTR2A, MCHR1, GABRG2, MCHR2, ADCYAP1, CORT, CCKBR, GLRB, SST, NMU, CRH, TAC3, TAC1, VIP, GABRD |
| GABAergic synapse | Down-regulated | 0.000002641 | 0.0002252 | 6.71 | 86.16 | GABBR2, GABRA1, GNG3, GNG2, SLC32A1, GABRA5, GABRA4, GAD2, PRKACB, GABRD, GABRG2 | |
| Complement and coagulation cascades | Up-regulated | 8.435 × 10−15 | 1.839 × 10−12 | 17.28 | 560.01 | C1QB, C1QA, C1S, CFH, C1R, C5AR1, CFI, F13A1, SERPINA5, C4B, C4A, C7, CFHR1, C3AR1, VSIG4, CFB, C1QC | |
| Hippocampus | Bacterial invasion of epithelial cells | Down-regulated | 0.0003395 | 0.01724 | 13.26 | 105.91 | CDC42, ARPC1A, ARPC4, MET |
| Synaptic vesicle cycle | Down-regulated | 0.0003567 | 0.01724 | 13.08 | 103.83 | ATP6V1G2, SLC32A1, SLC17A6, SLC1A6 | |
| Morphine addiction | Down-regulated | 0.0006406 | 0.01724 | 11.12 | 81.74 | GABRA1, SLC32A1, GNG4, PDE2A | |
| Entorhinal cortex | Morphine addiction | Down-regulated | 0.00008088 | 0.009868 | 12.52 | 118.00 | GABRB2, GABRA1, GNAS, ADCY1, GABRD |
| Gap junction | Down-regulated | 0.0008877 | 0.02926 | 10.15 | 71.31 | GNAS, ADRB1, ADCY1, TUBB4A | |
| GABAergic synapse | Down-regulated | 0.0009261 | 0.02926 | 10.03 | 70.04 | GABRB2, GABRA1, ADCY1, GABRD | |
| Temporal cortex | GABAergic synapse | Down-regulated | 4.244 × 10−10 | 9.295 × 10−8 | 12.30 | 265.54 | GABRB3, GABRB2, GABBR2, GABRA1, SLC12A5, PRKCB, GABRA5, GAD2, GABRG2, GLS, GNG3, GNG2, GABRD |
| Nicotine addiction | Down-regulated | 4.089 × 10−9 | 4.478 × 10−7 | 20.63 | 398.46 | GABRB3, GABRB2, GRIA2, GABRA1, GABRA5, SLC17A6, GABRD, GRIN2B, GABRG2 | |
| Retrograde endocannabinoid signaling | Down-regulated | 0.000001505 | 0.00007431 | 6.30 | 84.53 | GABRB3, GRIA2, GABRB2, MAPK9, GABRA1, GNG3, GNG2, PRKCB, GABRA5, SLC17A6, GABRD, GABRG2 |
| Name | Pathway | Degree | Betweenness |
|---|---|---|---|
| miRNA | |||
| hsa-mir-17-5p | GABAergic synapse pathway | 2 | 887.4 |
| Retrograde endocannabinoid signaling | 3 | 1135.6 | |
| hsa-mir-106a-5p | GABAergic synapse pathway | 2 | 887.4 |
| Retrograde endocannabinoid signaling | 3 | 1135.96 | |
| hsa-mir-373-3p | GABAergic synapse pathway | 1 | 0 |
| Retrograde endocannabinoid signaling | 1 | 0 | |
| Transcription Factors | |||
| ELK1 | GABAergic synapse pathway | 2 | 7.35 |
| Retrograde endocannabinoid signaling | 2 | 8.13 | |
| GATA1 | GABAergic synapse pathway | 1 | 0 |
| Retrograde endocannabinoid signaling | 1 | 0 | |
| GATA2 | GABAergic synapse pathway | 10 | 245.61 |
| Retrograde endocannabinoid signaling | 10 | 261.51 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abyadeh, M.; Tofigh, N.; Hosseinian, S.; Hasan, M.; Amirkhani, A.; Fitzhenry, M.J.; Gupta, V.; Chitranshi, N.; Salekdeh, G.H.; Haynes, P.A.; et al. Key Genes and Biochemical Networks in Various Brain Regions Affected in Alzheimer’s Disease. Cells 2022, 11, 987. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060987
Abyadeh M, Tofigh N, Hosseinian S, Hasan M, Amirkhani A, Fitzhenry MJ, Gupta V, Chitranshi N, Salekdeh GH, Haynes PA, et al. Key Genes and Biochemical Networks in Various Brain Regions Affected in Alzheimer’s Disease. Cells. 2022; 11(6):987. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060987
Chicago/Turabian StyleAbyadeh, Morteza, Nahid Tofigh, Saeedeh Hosseinian, Mafruha Hasan, Ardeshir Amirkhani, Matthew J. Fitzhenry, Veer Gupta, Nitin Chitranshi, Ghasem H. Salekdeh, Paul A. Haynes, and et al. 2022. "Key Genes and Biochemical Networks in Various Brain Regions Affected in Alzheimer’s Disease" Cells 11, no. 6: 987. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11060987