
The Emerging Role of Innate Lymphoid Cells (ILCs) and Alarmins in Celiac Disease: An Update on Pathophysiological Insights, Potential Use as Disease Biomarkers, and Therapeutic Implications
 ,
,  , , ,
, , ,
Abstract
:1. Introduction
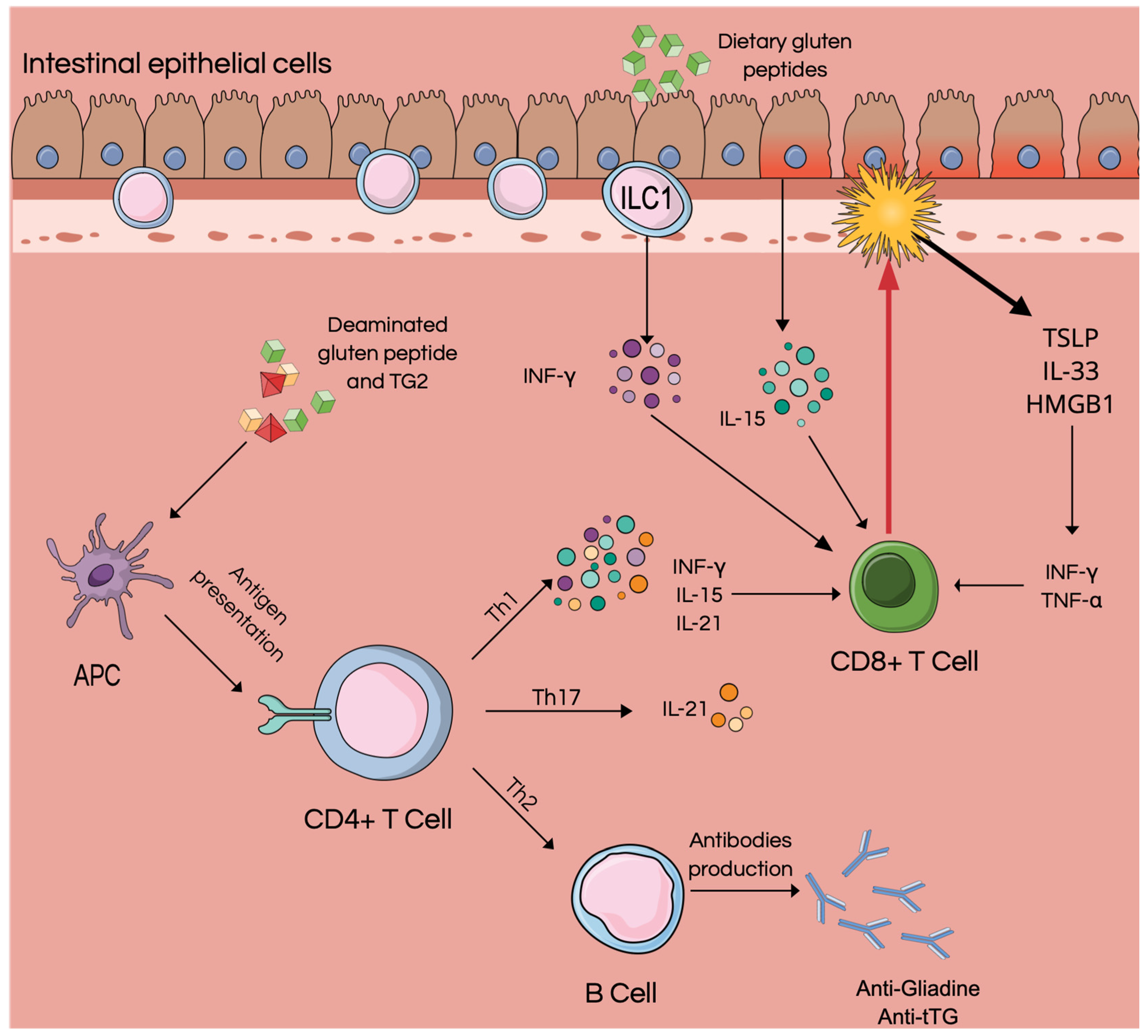
2. Intestinal Epithelial Cells
3. ILC
4. Role of Alarmins in Celiac Disease
4.1. Materials and Methods
4.2. Results
4.2.1. Nuclear Alarmins: Thymic Stromal Lymphopoietin (TSLP), IL-33, and HMBG1
4.2.2. Granule-Derived Alarmins: Defensins (α, β)
4.2.3. Cytoplasmic Alarmins: Calprotectin
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ludvigsson, J.F.; Leffler, D.A.; Bai, J.C.; Biagi, F.; Fasano, A.; Green, P.H.R.; Hadjivassiliou, M.; Kaukinen, K.; Kelly, C.P.; Leonard, J.N.; et al. The Oslo Definitions for Coeliac Disease and Related Terms. Gut 2013, 62, 43–52. [Google Scholar] [CrossRef]
- Aboulaghras, S.; Piancatelli, D.; Taghzouti, K.; Balahbib, A.; Alshahrani, M.M.; Al Awadh, A.A.; Goh, K.W.; Ming, L.C.; Bouyahya, A.; Oumhani, K. Meta-Analysis and Systematic Review of HLA DQ2/DQ8 in Adults with Celiac Disease. Int. J. Mol. Sci. 2023, 24, 1188. [Google Scholar] [CrossRef] [PubMed]
- Trynka, G.; Hunt, K.A.; Bockett, N.A.; Romanos, J.; Mistry, V.; Szperl, A.; Bakker, S.F.; Bardella, M.T.; Bhaw-Rosun, L.; Castillejo, G.; et al. Dense Genotyping Identifies and Localizes Multiple Common and Rare Variant Association Signals in Celiac Disease. Nat. Genet. 2011, 43, 1193–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, K.A.; Zhernakova, A.; Turner, G.; Heap, G.A.R.; Franke, L.; Bruinenberg, M.; Romanos, J.; Dinesen, L.C.; Ryan, A.W.; Panesar, D.; et al. Newly Identified Genetic Risk Variants for Celiac Disease Related to the Immune Response. Nat. Genet. 2008, 40, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Vargas, J.; Green, P.H.R.; Bhagat, G. Innate Lymphoid Cells and Celiac Disease: Current Perspective. Cell Mol. Gastroenterol. Hepatol. 2021, 11, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Guandalini, S.; Assiri, A. Celiac Disease: A Review. JAMA Pediatr. 2014, 168, 272–278. [Google Scholar] [CrossRef]
- Vivier, E.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Eberl, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate Lymphoid Cells: 10 Years On. Cell 2018, 174, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Ochel, A.; Tiegs, G.; Neumann, K. Type 2 Innate Lymphoid Cells in Liver and Gut: From Current Knowledge to Future Perspectives. Int. J. Mol. Sci. 2019, 20, 1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, G.-Y.; Giles, D.A.; Kronenberg, M. The Role of Innate Lymphoid Cells in Response to Microbes at Mucosal Surfaces. Mucosal. Immunol. 2020, 13, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Zindel, J.; Kubes, P. DAMPs, PAMPs, and LAMPs in Immunity and Sterile Inflammation. Annu Rev. Pathol. 2020, 15, 493–518. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Han, Z.; Oppenheim, J.J. Alarmins and Immunity. Immunol. Rev. 2017, 280, 41–56. [Google Scholar] [CrossRef]
- Nie, Y.; Yang, D.; Oppenheim, J.J. Alarmins and Antitumor Immunity. Clin. Ther. 2016, 38, 1042–1053. [Google Scholar] [CrossRef] [Green Version]
- Danieli, M.G.; Antonelli, E.; Piga, M.A.; Claudi, I.; Palmeri, D.; Tonacci, A.; Allegra, A.; Gangemi, S. Alarmins in Autoimmune Diseases. Autoimmun. Rev. 2022, 21, 103142. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, A.; Lo Presti, E.; Chini, R.; Gammeri, L.; Inchingolo, R.; Lohmeyer, F.M.; Nucera, E.; Gangemi, S. Emerging Role of Alarmins in Food Allergy: An Update on Pathophysiological Insights, Potential Use as Disease Biomarkers, and Therapeutic Implications. J. Clin. Med. 2023, 12, 2699. [Google Scholar] [CrossRef]
- Pastorelli, L.; De Salvo, C.; Cominelli, M.A.; Vecchi, M.; Pizarro, T.T. Novel Cytokine Signaling Pathways in Inflammatory Bowel Disease: Insight into the Dichotomous Functions of IL-33 during Chronic Intestinal Inflammation. Therap. Adv. Gastroenterol. 2011, 4, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Stricker, S.; Müller, M.; Zimmer, K.-P.; Jacob, R. Altered Posttranslational Modification of Microtubules Contributes to Disturbed Enterocyte Morphology in Celiac Disease. Int. J. Mol. Sci. 2023, 24, 2635. [Google Scholar] [CrossRef]
- Porpora, M.; Conte, M.; Lania, G.; Bellomo, C.; Rapacciuolo, L.; Chirdo, F.G.; Auricchio, R.; Troncone, R.; Auricchio, S.; Barone, M.V.; et al. Inflammation Is Present, Persistent and More Sensitive to Proinflammatory Triggers in Celiac Disease Enterocytes. Int. J. Mol. Sci. 2022, 23, 1973. [Google Scholar] [CrossRef]
- Bracken, S.; Byrne, G.; Kelly, J.; Jackson, J.; Feighery, C. Altered Gene Expression in Highly Purified Enterocytes from Patients with Active Coeliac Disease. BMC Genomics 2008, 9, 377. [Google Scholar] [CrossRef] [Green Version]
- Reinecker, H.C.; MacDermott, R.P.; Mirau, S.; Dignass, A.; Podolsky, D.K. Intestinal Epithelial Cells Both Express and Respond to Interleukin 15. Gastroenterology 1996, 111, 1706–1713. [Google Scholar] [CrossRef]
- Barone, M.V.; Troncone, R.; Auricchio, S. Gliadin Peptides as Triggers of the Proliferative and Stress/Innate Immune Response of the Celiac Small Intestinal Mucosa. Int. J. Mol. Sci. 2014, 15, 20518–20537. [Google Scholar] [CrossRef] [Green Version]
- Di Sabatino, A.; Ciccocioppo, R.; Cupelli, F.; Cinque, B.; Millimaggi, D.; Clarkson, M.M.; Paulli, M.; Cifone, M.G.; Corazza, G.R. Epithelium Derived Interleukin 15 Regulates Intraepithelial Lymphocyte Th1 Cytokine Production, Cytotoxicity, and Survival in Coeliac Disease. Gut 2006, 55, 469–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, E.C. Interleukin 15 Is a Potent Stimulant of Intraepithelial Lymphocytes. Gastroenterology 1998, 115, 1439–1445. [Google Scholar] [CrossRef]
- Nanayakkara, M.; Lania, G.; Maglio, M.; Kosova, R.; Sarno, M.; Gaito, A.; Discepolo, V.; Troncone, R.; Auricchio, S.; Auricchio, R.; et al. Enterocyte Proliferation and Signaling Are Constitutively Altered in Celiac Disease. PLoS ONE 2013, 8, e76006. [Google Scholar] [CrossRef] [Green Version]
- Clottu, A.S.; Humbel, M.; Fluder, N.; Karampetsou, M.P.; Comte, D. Innate Lymphoid Cells in Autoimmune Diseases. Front. Immunol. 2021, 12, 789788. [Google Scholar] [CrossRef]
- Artis, D.; Spits, H. The Biology of Innate Lymphoid Cells. Nature 2015, 517, 293–301. [Google Scholar] [CrossRef]
- Geremia, A.; Arancibia-Cárcamo, C.V. Innate Lymphoid Cells in Intestinal Inflammation. Front. Immunol. 2017, 8, 1296. [Google Scholar] [CrossRef]
- Marafini, I.; Monteleone, I.; Di Fusco, D.; Cupi, M.L.; Paoluzi, O.A.; Colantoni, A.; Ortenzi, A.; Izzo, R.; Vita, S.; De Luca, E.; et al. TNF-α Producing Innate Lymphoid Cells (ILCs) Are Increased in Active Celiac Disease and Contribute to Promote Intestinal Atrophy in Mice. PLoS ONE 2015, 10, e0126291. [Google Scholar] [CrossRef]
- Uhde, M.; Yu, X.; Bunin, A.; Brauner, C.; Lewis, S.K.; Lebwohl, B.; Krishnareddy, S.; Alaedini, A.; Reizis, B.; Ghosh, S.; et al. Phenotypic Shift of Small Intestinal Intra-Epithelial Type 1 Innate Lymphoid Cells in Celiac Disease Is Associated with Enhanced Cytotoxic Potential. Clin. Exp. Immunol. 2020, 200, 163–175. [Google Scholar] [CrossRef]
- Ercolano, G.; Moretti, A.; Falquet, M.; Wyss, T.; Tran, N.L.; Senoner, I.; Marinoni, M.; Agosti, M.; Salvatore, S.; Jandus, C.; et al. Gliadin-Reactive Vitamin D-Sensitive Proinflammatory ILCPs Are Enriched in Celiac Patients. Cell Rep. 2022, 39, 110956. [Google Scholar] [CrossRef]
- Cseh, Á.; Vásárhelyi, B.; Szalay, B.; Molnár, K.; Nagy-Szakál, D.; Treszl, A.; Vannay, Á.; Arató, A.; Tulassay, T.; Veres, G. Immune Phenotype of Children with Newly Diagnosed and Gluten-Free Diet-Treated Celiac Disease. Dig. Dis. Sci. 2011, 56, 792–798. [Google Scholar] [CrossRef]
- Bernardo, D.; van Hoogstraten, I.M.W.; Verbeek, W.H.M.; Peña, A.S.; Mearin, M.L.; Arranz, E.; Garrote, J.A.; Scheper, R.J.; Schreurs, M.W.J.; Bontkes, H.J.; et al. Decreased Circulating INKT Cell Numbers in Refractory Coeliac Disease. Clin. Immunol. 2008, 126, 172–179. [Google Scholar] [CrossRef]
- Perez, F.; Ruera, C.N.; Miculan, E.; Carasi, P.; Chirdo, F.G. Programmed Cell Death in the Small Intestine: Implications for the Pathogenesis of Celiac Disease. Int. J. Mol. Sci. 2021, 22, 7426. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Geha, R.S. Thymic Stromal Lymphopoietin. Ann. N. Y. Acad. Sci. 2010, 1183, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Takai, T. TSLP Expression: Cellular Sources, Triggers, and Regulatory Mechanisms. Allergol. Int. 2012, 61, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Rimoldi, M.; Chieppa, M.; Salucci, V.; Avogadri, F.; Sonzogni, A.; Sampietro, G.M.; Nespoli, A.; Viale, G.; Allavena, P.; Rescigno, M. Intestinal Immune Homeostasis Is Regulated by the Crosstalk between Epithelial Cells and Dendritic Cells. Nat. Immunol. 2005, 6, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Rescigno, M.; Di Sabatino, A. Dendritic Cells in Intestinal Homeostasis and Disease. J. Clin. Investig. 2009, 119, 2441–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Jeong, D.Y.; Peyrin-Biroulet, L.; Eisenhut, M.; Shin, J.I. Insight into the Role of TSLP in Inflammatory Bowel Diseases. Autoimmun. Rev. 2017, 16, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C. IL-33, an Alarmin of the IL-1 Family Involved in Allergic and Non Allergic Inflammation: Focus on the Mechanisms of Regulation of Its Activity. Cells 2021, 11, 107. [Google Scholar] [CrossRef]
- Martin, N.T.; Martin, M.U. Interleukin 33 Is a Guardian of Barriers and a Local Alarmin. Nat. Immunol. 2016, 17, 122–131. [Google Scholar] [CrossRef]
- Cayrol, C.; Girard, J.-P. Interleukin-33 (IL-33): A Nuclear Cytokine from the IL-1 Family. Immunol. Rev. 2018, 281, 154–168. [Google Scholar] [CrossRef]
- Liew, F.Y.; Girard, J.-P.; Turnquist, H.R. Interleukin-33 in Health and Disease. Nat. Rev. Immunol. 2016, 16, 676–689. [Google Scholar] [CrossRef]
- Yang, H.; Lundbäck, P.; Ottosson, L.; Erlandsson-Harris, H.; Venereau, E.; Bianchi, M.E.; Al-Abed, Y.; Andersson, U.; Tracey, K.J. Redox Modifications of Cysteine Residues Regulate the Cytokine Activity of HMGB1. Mol. Med. 2021, 27, 58. [Google Scholar] [CrossRef]
- Taverna, S.; Tonacci, A.; Ferraro, M.; Cammarata, G.; Cuttitta, G.; Bucchieri, S.; Pace, E.; Gangemi, S. High Mobility Group Box 1: Biological Functions and Relevance in Oxidative Stress Related Chronic Diseases. Cells 2022, 11, 849. [Google Scholar] [CrossRef]
- Tang, D.; Shi, Y.; Kang, R.; Li, T.; Xiao, W.; Wang, H.; Xiao, X. Hydrogen Peroxide Stimulates Macrophages and Monocytes to Actively Release HMGB1. J. Leukoc. Biol. 2007, 81, 741–747. [Google Scholar] [CrossRef] [Green Version]
- Biancheri, P.; Di Sabatino, A.; Rescigno, M.; Giuffrida, P.; Fornasa, G.; Tsilingiri, K.; Pender, S.L.F.; Papadia, C.; Wood, E.; Pasini, A.; et al. Abnormal Thymic Stromal Lymphopoietin Expression in the Duodenal Mucosa of Patients with Coeliac Disease. Gut 2016, 65, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Sziksz, E.; Molnár, K.; Lippai, R.; Pap, D.; Onody, A.; Veres-Székely, A.; Vörös, P.; Szabó, D.; Győrffy, H.; Veres, G.; et al. Peroxisome Proliferator-Activated Receptor-γ and Thymic Stromal Lymphopoietin Are Involved in the Pathophysiology of Childhood Coeliac Disease. Virchows Arch. 2014, 465, 385–393. [Google Scholar] [CrossRef]
- Kahramanoğlu Aksoy, E.; Akpınar, M.Y.; Pirinççi Sapmaz, F.; Doğan, Ö.; Uzman, M.; Nazlıgül, Y. Thymic Stromal Lymphopoietin Levels Are Increased in Patients with Celiac Disease. Bosn. J. Basic Med. Sci. 2019, 19, 282–287. [Google Scholar] [CrossRef] [Green Version]
- López-Casado, M.A.; Lorite, P.; Palomeque, T.; Torres, M.I. Potential Role of the IL-33/ST2 Axis in Celiac Disease. Cell Mol. Immunol. 2017, 14, 285–292. [Google Scholar] [CrossRef]
- Perez, F.; Ruera, C.N.; Miculan, E.; Carasi, P.; Dubois-Camacho, K.; Garbi, L.; Guzman, L.; Hermoso, M.A.; Chirdo, F.G. IL-33 Alarmin and Its Active Proinflammatory Fragments Are Released in Small Intestine in Celiac Disease. Front. Immunol. 2020, 11, 581445. [Google Scholar] [CrossRef]
- Manti, S.; Cuppari, C.; Tardino, L.; Parisi, G.; Spina, M.; Salpietro, C.; Leonardi, S. HMGB1 as a New Biomarker of Celiac Disease in Children: A Multicenter Study. Nutrition 2017, 37, 18–21. [Google Scholar] [CrossRef]
- Wilson, S.S.; Wiens, M.E.; Smith, J.G. Antiviral Mechanisms of Human Defensins. J. Mol. Biol. 2013, 425, 4965–4980. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ding, J.; Liao, C.; Xu, J.; Liu, X.; Lu, W. Defensins: The Natural Peptide Antibiotic. Adv. Drug Deliv. Rev. 2021, 179, 114008. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.L.; Salzman, N.H. Paneth Cells, Antimicrobial Peptides and Maintenance of Intestinal Homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef]
- Zong, X.; Fu, J.; Xu, B.; Wang, Y.; Jin, M. Interplay between Gut Microbiota and Antimicrobial Peptides. Anim. Nutr. 2020, 6, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Shumatova, T.A.; Zernova, E.S.; Grigoryan, L.A.; Shishatskaya, S.N. Antimicrobial Peptides and Proteins in the Diagnosis of Malabsorption Syndrome in Children. Modern Prob. Sci. Educ. 2015, 3. [Google Scholar]
- Almeida, T.T.G.; Monteiro, M.V.B.; Guimarães, R.C.; Casseb, A.R.; Huffman, M.A.; Gonçalves, E.C.; Monteiro, F.O.B.; Silva Filho, E. Effect of Gluten Diet on Blood Innate Immune Gene Expressions and Stool Consistency in Spix’s Saddleback Tamarin (Leontocebus Fuscicollis) Raised in Captivity. Mol Biol Rep 2019, 46, 3617–3623. [Google Scholar] [CrossRef]
- Wang, G. Human Antimicrobial Peptides and Proteins. Pharmaceuticals 2014, 7, 545–594. [Google Scholar] [CrossRef] [Green Version]
- Frye, M.; Bargon, J.; Lembcke, B.; Wagner, T.O.; Gropp, R. Differential Expression of Human Alpha- and Beta-Defensins MRNA in Gastrointestinal Epithelia. Eur. J. Clin. Investig. 2000, 30, 695–701. [Google Scholar] [CrossRef]
- Forsberg, G.; Fahlgren, A.; Hörstedt, P.; Hammarström, S.; Hernell, O.; Hammarström, M.-L. Presence of Bacteria and Innate Immunity of Intestinal Epithelium in Childhood Celiac Disease. Am. J. Gastroenterol. 2004, 99, 894–904. [Google Scholar] [CrossRef]
- Taha, A.S.; Faccenda, E.; Angerson, W.J.; Balsitis, M.; Kelly, R.W. Natural Antibiotic Expression in Celiac Disease—Correlation with Villous Atrophy and Response to a Gluten-Free Diet. Dig. Dis. Sci. 2005, 50, 791–795. [Google Scholar] [CrossRef]
- Di Sabatino, A.; Miceli, E.; Dhaliwal, W.; Biancheri, P.; Salerno, R.; Cantoro, L.; Vanoli, A.; De Vincenzi, M.; Blanco, C.D.V.; MacDonald, T.T.; et al. Distribution, Proliferation, and Function of Paneth Cells in Uncomplicated and Complicated Adult Celiac Disease. Am. J. Clin. Pathol. 2008, 130, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Vordenbäumen, S.; Pilic, D.; Otte, J.-M.; Schmitz, F.; Schmidt-Choudhury, A. Defensin-MRNA Expression in the Upper Gastrointestinal Tract Is Modulated in Children with Celiac Disease and Helicobacter Pylori-Positive Gastritis. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 596–600. [Google Scholar] [CrossRef]
- Intrieri, M.; Rinaldi, A.; Scudiero, O.; Autiero, G.; Castaldo, G.; Nardone, G. Low Expression of Human Beta-Defensin 1 in Duodenum of Celiac Patients Is Partially Restored by a Gluten-Free Diet. Clin. Chem. Lab. Med. 2010, 48, 489–492. [Google Scholar] [CrossRef]
- Fernandez-Jimenez, N.; Castellanos-Rubio, A.; Plaza-Izurieta, L.; Gutierrez, G.; Castaño, L.; Vitoria, J.C.; Bilbao, J.R. Analysis of Beta-Defensin and Toll-like Receptor Gene Copy Number Variation in Celiac Disease. Hum. Immunol. 2010, 71, 833–836. [Google Scholar] [CrossRef]
- Kamilova, A.T.; Azizova, G.K.; Umarnazarova, Z.E.; Abdullaeva, D.A.; Geller, S.I. The Activity of Antimicrobial Peptides in Pediatric Celiac Disease. Front. Pediatr. 2022, 10, 873793. [Google Scholar] [CrossRef]
- Jukic, A.; Bakiri, L.; Wagner, E.F.; Tilg, H.; Adolph, T.E. Calprotectin: From Biomarker to Biological Function. Gut 2021, 70, 1978–1988. [Google Scholar] [CrossRef]
- Donato, R.; Cannon, B.R.; Sorci, G.; Riuzzi, F.; Hsu, K.; Weber, D.J.; Geczy, C.L. Functions of S100 Proteins. Curr. Mol. Med. 2013, 13, 24–57. [Google Scholar] [CrossRef] [Green Version]
- Montalto, M.; Santoro, L.; Curigliano, V.; D’Onofrio, F.; Cammarota, G.; Panunzi, S.; Ricci, R.; Gallo, A.; Grieco, A.; Gasbarrini, A.; et al. Faecal Calprotectin Concentrations in Untreated Coeliac Patients. Scand. J. Gastroenterol. 2007, 42, 957–961. [Google Scholar] [CrossRef]
- Ertekin, V.; Selimoğlu, M.A.; Turgut, A.; Bakan, N. Fecal Calprotectin Concentration in Celiac Disease. J. Clin. Gastroenterol. 2010, 44, 544–546. [Google Scholar] [CrossRef]
- Balamtekın, N.; Baysoy, G.; Uslu, N.; Orhan, D.; Akçören, Z.; Özen, H.; Gürakan, F.; Saltik-Temızel, İ.N.; Yüce, A. Fecal Calprotectin Concentration Is Increased in Children with Celiac Disease: Relation with Histopathological Findings. Turk. J. Gastroenterol. 2012, 23, 503–508. [Google Scholar] [CrossRef]
- Capone, P.; Rispo, A.; Imperatore, N.; Caporaso, N.; Tortora, R. Fecal Calprotectin in Coeliac Disease. World J. Gastroenterol. 2014, 20, 611–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szaflarska-Popławska, A.; Romańczuk, B.; Parzęcka, M. Faecal Calprotectin Concentration in Children with Coeliac Disease. Prz. Gastroenterol. 2020, 15, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Corren, J.; Garcia Gil, E.; Griffiths, J.M.; Parnes, J.R.; van der Merwe, R.; Sałapa, K.; O’Quinn, S. Tezepelumab Improves Patient-Reported Outcomes in Patients with Severe, Uncontrolled Asthma in PATHWAY. Ann. Allergy Asthma Immunol. 2021, 126, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Ebina-Shibuya, R.; Leonard, W.J. TSLP-Targeting Therapy: Beyond Allergy? Clin. Transl. Med. 2023, 13, e1241. [Google Scholar] [CrossRef]
- Ebina-Shibuya, R.; Leonard, W.J. Role of Thymic Stromal Lymphopoietin in Allergy and Beyond. Nat. Rev. Immunol. 2023, 23, 24–37. [Google Scholar] [CrossRef]
- Tonner, H.; Hunn, S.; Auler, N.; Schmelter, C.; Beutgen, V.M.; von Pein, H.D.; Pfeiffer, N.; Grus, F.H. A Monoclonal Anti-HMGB1 Antibody Attenuates Neurodegeneration in an Experimental Animal Model of Glaucoma. Int. J. Mol. Sci. 2022, 23, 4107. [Google Scholar] [CrossRef]
- Nishibori, M.; Mori, S.; Takahashi, H.K. Anti-HMGB1 Monoclonal Antibody Therapy for a Wide Range of CNS and PNS Diseases. J. Pharmacol. Sci. 2019, 140, 94–101. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Ouchi, Y. Antimicrobial Peptide Defensin: Identification of Novel Isoforms and the Characterization of Their Physiological Roles and Their Significance in the Pathogenesis of Diseases. Proc. Jpn Acad. Ser. B Phys. Biol. Sci. 2012, 88, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Fellermann, K.; Wehkamp, J.; Herrlinger, K.R.; Stange, E.F. Crohn’s Disease: A Defensin Deficiency Syndrome? Eur. J. Gastroenterol. Hepatol. 2003, 15, 627–634. [Google Scholar] [CrossRef]
- Wehkamp, J.; Schmid, M.; Fellermann, K.; Stange, E.F. Defensin Deficiency, Intestinal Microbes, and the Clinical Phenotypes of Crohn’s Disease. J. Leukoc. Biol. 2005, 77, 460–465. [Google Scholar] [CrossRef] [Green Version]
- Kolbinger, F.; Loesche, C.; Valentin, M.-A.; Jiang, X.; Cheng, Y.; Jarvis, P.; Peters, T.; Calonder, C.; Bruin, G.; Polus, F.; et al. β-Defensin 2 Is a Responsive Biomarker of IL-17A-Driven Skin Pathology in Patients with Psoriasis. J. Allergy Clin. Immunol. 2017, 139, 923–932.e8. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, W.; Shi, H.; Fu, Y.; Chen, X.; Chen, X.; Liu, Y.; Kan, B.; Wang, Y. Gene Therapy with Beta-Defensin 2 Induces Antitumor Immunity and Enhances Local Antitumor Effects. Hum. Gene Ther. 2014, 25, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Cieślik, M.; Bagińska, N.; Górski, A.; Jończyk-Matysiak, E. Human β-Defensin 2 and Its Postulated Role in Modulation of the Immune Response. Cells 2021, 10, 2991. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Alarmins | Author | Age | F/M | N° of Patients | Country | Year [Ref.] |
|---|---|---|---|---|---|---|
| Studies on humans | ||||||
| Nuclear | ||||||
| TSLP | Biancheri et al. | 39 years | 6/6 * | 114 | Italy, UK | 2016 [45] |
| Sziksz et al. | 9 years | 16/9 | 25 | Hungary | 2014 [46] | |
| Kahramanoğlu Aksoy et al. | 41 years | 53/20 | 73 | Turkey | 2019 [47] | |
| IL-33 | López-Casado et al. | 6 years | 12/8 | 20 | Spain | 2017 [48] |
| Perez et al. | 19 years | 39/25 | 64 | Argentina, Chile | 2020 [49] | |
| HMGB1 | Manti et al. | 6 years | 16/33 | 49 | Italy | 2016 [50] |
| Granule-derived | ||||||
| Defensin (α, β) | Forsberg et al. | 6 years | 95/55 | 150 | Sweden | 2004 [59] |
| Taha et al. | 46 years | Unknown | 13 | UK | 2005 [60] | |
| Di Sabatino et al. | 36 years | Unknown | 14 | Italy | 2008 [61] | |
| Vordenbäumen et al. | 4–16 years | 8/3 | 11 | Germany | 2010 [62] | |
| Intrieri et al. | 35 years | Unknown | 21 | Italy | 2010 [63] | |
| Fernandez-Jimenez et al. | 4 years | 241/135 | 376 | Spain | 2010 [64] | |
| Cytoplasmic | ||||||
| Calprotectine | Montalto et al. | 38 years | 12/16 | 28 | Italy | 2007 [68] |
| Ertekin et al. | 6 years | 15/14 | 29 | Turkey | 2010 [69] | |
| Balamtekın et al. | 8 years | 30/34 | 64 | Turkey | 2012 [70] | |
| Capone et al. | Unknown | Unknown | 50 | Italy | 2014 [71] | |
| Szaflarska-Popławska et al. | 10 years | 34/38 | 72 | Poland | 2020 [72] | |
| Studies on animal models | ||||||
| Β-Defensin | Almeida et al. | \ | \ | \ | Brazil | 2019 [56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzi, A.; Di Gioacchino, M.; Gammeri, L.; Inchingolo, R.; Chini, R.; Santilli, F.; Nucera, E.; Gangemi, S. The Emerging Role of Innate Lymphoid Cells (ILCs) and Alarmins in Celiac Disease: An Update on Pathophysiological Insights, Potential Use as Disease Biomarkers, and Therapeutic Implications. Cells 2023, 12, 1910. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12141910
Rizzi A, Di Gioacchino M, Gammeri L, Inchingolo R, Chini R, Santilli F, Nucera E, Gangemi S. The Emerging Role of Innate Lymphoid Cells (ILCs) and Alarmins in Celiac Disease: An Update on Pathophysiological Insights, Potential Use as Disease Biomarkers, and Therapeutic Implications. Cells. 2023; 12(14):1910. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12141910
Chicago/Turabian StyleRizzi, Angela, Mario Di Gioacchino, Luca Gammeri, Riccardo Inchingolo, Raffaella Chini, Francesca Santilli, Eleonora Nucera, and Sebastiano Gangemi. 2023. "The Emerging Role of Innate Lymphoid Cells (ILCs) and Alarmins in Celiac Disease: An Update on Pathophysiological Insights, Potential Use as Disease Biomarkers, and Therapeutic Implications" Cells 12, no. 14: 1910. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12141910