
Antifibrotic Effect of Selenium-Containing Nanoparticles on a Model of TAA-Induced Liver Fibrosis
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Injection Protocol
2.4. Method for Obtaining Selenium Nanoparticles
2.5. RNA Isolation, Reverse Transcription, RT-PCR
2.6. Western Blotting
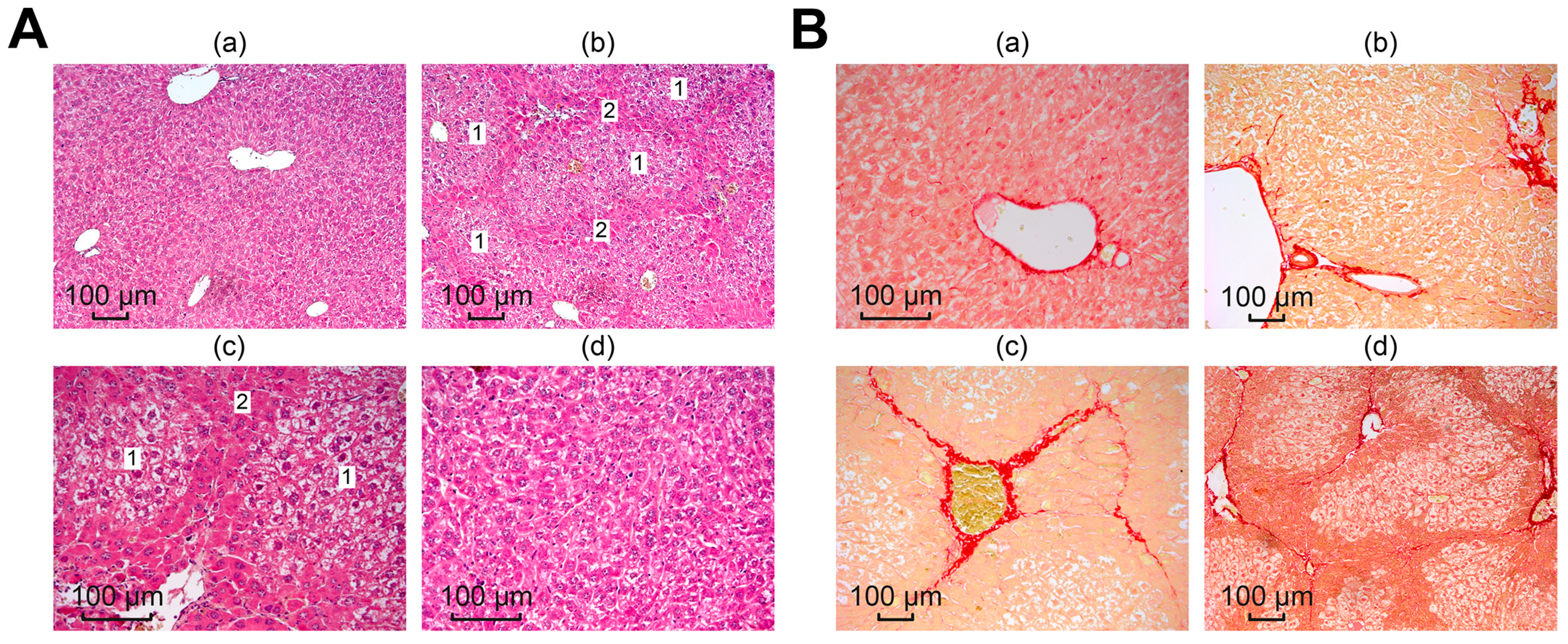
2.7. Histological Analysis
2.8. Measurement of ALT/AST Activities
2.9. Statistical Data Processing
3. Results and Discussion
3.1. General Characteristics of the Obtained Selenium Nanoparticles
3.2. The Weight of Animals Returns to Normal when Injected with Selenium-Containing Nanoparticles
3.3. Selenium-Containing Nanoparticles Have an Antifibrotic Effect
3.4. Selenium-Containing Nanoparticles Reduce Cytolysis, Normalizing ALT and AST Levels in the Blood of Animals
3.5. Selenium-Containing Nanoparticles Have an Anti-Inflammatory Effect
3.6. Selenium-Containing Nanoparticles Neutralize the Effects of Prolonged ER-Stress and Apoptosis in Liver Cells Caused by the Action of TAA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Iredale, J.P. Models of liver fibrosis: Exploring the dynamic nature of inflammation and repair in a solid organ. J. Clin. Investig. 2007, 117, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Kisseleva, T.; Brenner, D.A. Mechanisms of fibrogenesis. Exp. Biol. Med. 2008, 233, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Ekihiro, S.; Brenner, D.A. Recent advancement of molecular mechanisms of liver fibrosis. J. Hepato Biliary Pancreat. Sci. 2015, 22, 512–518. [Google Scholar]
- Brenner, D.A. Reversibility of liver fibrosis. Gastroenterol. Hepatol. 2013, 9, 737–739. [Google Scholar]
- Aydın, M.M.; Akçalı, K.C. Liver fibrosis. Turk. J. Gastroenterol. 2018, 29, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.M.; Adnane, L.; Newell, P.; Villanueva, A.; Llovet, J.M.; Lynch, M. Preclinical overview of sorafenib, a multikinase inhibitor that targets both Raf and VEGF and PDGF receptor tyrosine kinase signaling. Mol. Cancer Ther. 2008, 7, 3129–3140. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; de Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in advanced hepatocellular carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Escudier, B.; Eisen, T.; Stadler, W.M.; Szczylik, C.; Oudard, S.; Siebels, M.; Negrier, S.; Chevreau, C.; Solska, E.; Desai, A.A.; et al. Sorafenib in advanced clear-cell renal-cell carcinoma. N. Engl. J. Med. 2007, 356, 125–134. [Google Scholar] [CrossRef]
- Kroschinsky, F.; Stolzel, F.; von Bonin, S.; Beutel, G.; Kochanek, M.; Kiehl, M.; Schellongowski, P. New drugs, new toxicities: Severe side effects of modern targeted and immunotherapy of cancer and their management. Crit. Care 2017, 21, 89. [Google Scholar] [CrossRef]
- Hutson, T.E.; Al-Shukri, S.; Stus, V.P.; Lipatov, O.N.; Shparyk, Y.; Bair, A.H.; Rosbrook, B.; Andrews, G.I.; Vogelzang, N.J. Axitinib versus sorafenib in first-line metastatic renal cell carcinoma: Overall survival from a randomized phase III trial. Clin. Genitourin. Cancer 2017, 15, 72–76. [Google Scholar] [CrossRef]
- Chang, Y.; He, L.; Li, Z.; Zeng, L.; Song, Z.; Li, P.; Chan, L.; You, Y.; Yu, X.F.; Chu, P.K.; et al. Designing core-shell gold and selenium nanocomposites for cancer radiochemotherapy. ACS Nano 2017, 11, 4848–4858. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Lin, Z.; Li, Y.; Zhao, M.; Wang, C.; Guo, M.; Zhang, B.; Zhu, B. Targeted delivery of siRNA using RGDfC-conjugated functionalized selenium nanoparticles for anticancer therapy. J. Mater. Chem. 2017, 5, 6941–6952. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Gudkov, S.V.; Plotnikov, E.Y.; Turovsky, E.A. Size-Dependent Cytoprotective Effects of Selenium Nanoparticles during Oxygen-Glucose Deprivation in Brain Cortical Cells. Int. J. Mol. Sci. 2022, 23, 7464. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Khabatova, V.V.; Gudkov, S.V.; Turovsky, E.A. Ca2+-Dependent Effects of the Selenium-Sorafenib Nanocomplex on Glioblastoma Cells and Astrocytes of the Cerebral Cortex: Anticancer Agent and Cytoprotector. Int. J. Mol. Sci. 2023, 24, 2411. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Turovsky, E.A.; Blinova, E.V. Therapeutic Potential and Main Methods of Obtaining Selenium Nanoparticles. Int. J. Mol. Sci. 2021, 22, 10808. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Yang, F.; Fan, S.; Jin, H.; Liao, K.; Li, X.; Liu, G.B.; Liang, J.; Zhang, J.; Xu, J.F.; et al. Immunomodulatory roles of selenium nanoparticles: Novel arts for potential immunotherapy strategy development. Front Immunol. 2022, 26, 956181. [Google Scholar] [CrossRef]
- Fang, X.; Li, C.E.; Zheng, L.; Yang, F.; Chen, T. Dual-Targeted selenium nanoparticles for synergistic photothermal therapy and chemotherapy of tumors. Chem.—Asian J. 2018, 13, 996–1004. [Google Scholar] [CrossRef]
- Huang, G.; Liu, Z.; He, L.; Luk, K.H.; Cheung, S.T.; Wong, K.H.; Chen, T. Autophagy is an important action mode for functionalized selenium nanoparticles to exhibit anticolorectal cancer activity. Biomater. Sci. 2018, 6, 2508–2517. [Google Scholar] [CrossRef]
- Goltyaev, M.V.; Varlamova, E.G. The Role of Selenium Nanoparticles in the Treatment of Liver Pathologies of Various Natures. Int. J. Mol. Sci. 2023, 24, 10547. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Wong, Y.S.; Chen, T.; Zhang, H.; Liu, C. The reversal of cisplatin–induced nephrotoxicity by selenium nanoparticles functionalized with 11–mercapto–1–undecanol by inhibition of ROS–mediated apoptosis. Biomaterials 2011, 32, 9068–9076. [Google Scholar] [CrossRef]
- Tian, J.; Wei, X.; Zhang, W.; Xu, A. Effects of selenium nanoparticles combined with radiotherapy on lung cancer cells. Front. Bioeng. Biotechnol. 2020, 8, 598997. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Zhang, H.; Niu, Y.; Wu, Y.; Sun, W.; Li, H.; Kong, J.; Ding, K.; Shen, H.M.; Wu, H. Erratum to: Long non–coding RNA linc00673 regulated non–small cell lung cancer proliferation, migration, invasion and epithelial mesenchymal transition by sponging miR–150–5p. Mol. Cancer 2017, 16, 144. [Google Scholar] [CrossRef]
- Huang, T.; Holden, J.A.; Heath, D.E.; O’Brien–Simpson, N.M.; O’Connor, A.J. Engineering highly effective antimicrobial selenium nanoparticles through control of particle size. Nanoscale 2019, 11, 14937–14951. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zhao, J.; Liu, P.; Ji, D.; Zhang, L.; Zhang, M.; Li, Y.; Xiao, Y. Preparation and in vitro evaluation of multi–target–directed selenium–chondroitin sulfate nanoparticles in protecting against the Alzheimer’s disease. Int. J. Biol. Macromol. 2020, 142, 265–276. [Google Scholar] [CrossRef]
- Duma, N.; Santana–Davila, R.; Molina, J.R. Non–small cell lung cancer: Epidemiology, Screen. Diagn. Treat. Mayo Clin. Proc. 2019, 94, 1623–1640. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Li, C.; Huang, X.; Lin, X.; Lin, W.; Yang, F.; Chen, T. Thermosensitive hydrogels for sustained-release of sorafenib and selenium nanoparticles for localized synergistic chemoradiotherapy. Biomaterials 2019, 216, 119220. [Google Scholar] [CrossRef] [PubMed]
- Varlamova, E.G.; Goltyaev, M.V.; Simakin, A.V.; Gudkov, S.V.; Turovsky, E.A. Comparative Analysis of the Cytotoxic Effect of a Complex of Selenium Nanoparticles Doped with Sorafenib, “Naked” Selenium Nanoparticles, and Sorafenib on Human Hepatocyte Carcinoma HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 6641. [Google Scholar] [CrossRef]
- Baimler, I.V.; Simakin, A.V.; Chevokin, V.K.; Podvyaznikov, V.A.; Gudkov, S.V. Features of optical breakdown of aqueous colloidal solutions of ferric oxide (Fe2O3) nanoparticles occurring on individual or on two closely located nanoparticles. Chem. Phys. Lett. 2021, 776, 138697. [Google Scholar] [CrossRef]
- Baymler, I.V.; Simakin, A.V.; Gudkov, S.V. Investigation of the laser-induced breakdown plasma, acoustic vibrations and dissociation processes of water molecules caused by laser breakdown of colloidal solutions containing Ni nanoparticles. Plasma Sources Sci. Technol. 2021, 30, 125015. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Astashev, M.E.; Baimler, I.V.; Uvarov, O.V.; Voronov, V.V.; Simakin, A.V. Laser-Induced Optical Breakdown of an Aqueous Colloidal Solution Containing Terbium Nanoparticles: The Effect of Oxidation of Nanoparticles. J. Phys. Chem. 2022, 126, 5678–5688. [Google Scholar] [CrossRef]
- Nagaev, E.I.; Baimler, I.V.; Baryshev, A.S.; Astashev, M.E.; Gudkov, S.V. Effect of Laser-Induced Optical Breakdown on the Structure of Bsa Molecules in Aqueous Solutions: An Optical Study. Molecules 2022, 27, 6752. [Google Scholar] [CrossRef]
- Wallace, M.C.; Hamesch, K.; Lunova, M.; Kim, Y.; Weiskirchen, R.; Strnad, P.; Friedman, S.L. Standard operating procedures in experimental liver research: Thioacetamide model in mice and rats. Lab. Anim. 2015, 49, 21–29. [Google Scholar] [CrossRef]
- Inoue, T.; Ishizaka, Y.; Sasaki, E.; Lu, J.; Mineshige, T.; Yanase, M.; Sasaki, E.; Shimoda, M. Thioacetamide-induced hepatic fibrosis in the common marmoset. Exp. Anim. 2018, 67, 321–327. [Google Scholar] [CrossRef]
- Matsuo, M.; Murata, S.; Hasegawa, S.; Hatada, Y.; Ohtsuka, M.; Taniguchi, H. Novel liver fibrosis model in Macaca fascicularis induced by thioacetamide. Sci. Rep. 2020, 10, 2450. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.J.; Ha, G.-H.; Seo, W.-Y.; Kim, C.K.; Kim, K.J.; Lee, S.B. Human collagen alpha-2 type I stimulates collagen synthesis, wound healing, and elastin production in normal human dermal fibroblasts (HDFs). BMB Rep. 2020, 53, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Park, R.W. Regulation of type I collagen genes expression. Int. Rev. Immunol. 1995, 12, 177–185. [Google Scholar] [CrossRef]
- Myllyharju, J.; Kivirikko, K.I. Collagens and collagen-related diseases. Ann. Med. 2001, 33, 7–21. [Google Scholar] [CrossRef]
- Trojanowska, M.; LeRoy, E.C.; Eckes, B.; Krieg, T. Pathogenesis of fibrosis: Type 1 collagen and the skin. J. Mol. Med. 1998, 76, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Jimenez, S.A. Modulation of collagen gene expression: Its relation to fibrosis in systemic sclerosis and other disorders. Ann. Intern. Med. 1995, 122, 60–62. [Google Scholar] [CrossRef]
- Leask, A.; Abraham, D.J. TGF-β signaling and the fibrotic response. FASEB J. 2004, 18, 816–827. [Google Scholar] [CrossRef]
- Zhou, D.J.; Mu, D.; Jiang, M.D.; Zheng, S.M.; Zhang, Y.; He, S.; Weng, M.; Zeng, W.Z. Hepatoprotective effect of juglone on dimethylnitrosamine-induced liver fibrosis and its effect on hepatic antioxidant defence and the expression levels of α-SMA and collagen III. Mol. Med. Rep. 2015, 12, 4095–4102. [Google Scholar] [CrossRef]
- Higashi, T.; Friedman, S.; Hoshida, Y. Hepatic stellate cells as key target in liver fibrosis. Adv. Drug Deliv. Rev. 2017, 121, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Syed, S.; Kehar, S.I. Glial fibrillary acidic protein (GFAP) as a mesenchymal marker of early hepatic stellate cells activation in liver fibrosis in chronic hepatitis C infection. Pak. J. Med. Sci. 2014, 30, 1027–1032. [Google Scholar] [CrossRef]
- Taura, K.; De Minicis, S.; Seki, E.; Hatano, E.; Iwaisako, K.; Osterreicher, C.H.; Kodama, Y.; Miura, K.; Ikai, I.; Uemoto, S.; et al. Hepatic stellate cells secrete angiopoietin 1 that induces angiogenesis in liver fibrosis. Gastroenterology 2008, 135, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Kisseleva, T.; Brenner, D.A. Deficiency of NOX1 or NOX4 prevents liver inflammation and fibrosis in mice through inhibition of hepatic stellate cell activation. PLoS ONE 2015, 10, e0129743. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Ma, H.Y.; Zhong, Z.; Dhar, D.; Liu, X.; Xu, J.; Koyama, Y.; Nishio, T.K.D.; Karin, G.; Mccubbin, R.; et al. NADPH oxidase 1 in liver macrophages promotes inflammation and tumor development in mice. Gastroenterology 2019, 156, 1156–1172. [Google Scholar] [CrossRef] [PubMed]
- Hammerich, L.; Heymann, F.; Tacke, F. Role of IL-17 and Th17 cells in liver diseases. Clin. Dev. Immunol. 2011, 2011, 345803. [Google Scholar] [CrossRef]
- Fabre, T.; Molina, M.F.; Soucy, G.; Goulet, J.P.; Willems, B.; Villeneuve, J.P.; Bilodeau, M.; Shoukry, N.H. Type 3 cytokines IL-17A and IL-22 drive TGF-beta-dependent liver fibrosis. Sci. Immunol. 2018, 3, 7754. [Google Scholar] [CrossRef]
- Meng, F.; Wang, K.; Aoyama, T.; Grivennikov, S.I.; Paik, Y.; Scholten, D.; Cong, M.; Iwaisako, K.; Liu, X.; Zhang, M.; et al. Interleukin-17 signaling in inflammatory, Kupffer cells, and hepatic stellate cells exacerbates liver fibrosis in mice. Gastroenterology 2012, 143, e13. [Google Scholar] [CrossRef]
- Kong, X.; Feng, D.; Wang, H.; Hong, F.; Bertola, A.; Wang, F.S.; Gao, B. Interleukin-22 induces hepatic stellate cell senescence and restricts liver fibrosis in mice. Hepatology 2012, 56, 1150–1159. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Z.; Luan, Y.; Zou, Z.; Sun, Y.; Li, Y.; Jin, L.; Zhou, C.; Fu, J.; Gao, B.; et al. Pathological functions of interleukin-22 in chronic liver inflammation and fibrosis with hepatitis B virus infection by promoting T helper 17 cell recruitment. Hepatology 2014, 59, 1331–1342. [Google Scholar] [CrossRef]
- Radaeva, S.; Sun, R.; Pan, H.N.; Hong, F.; Gao, B. Interleukin 22 (IL-22) plays a protective role in T cell-mediated murine hepatitis: IL-22 is a survival factor for hepatocytes via STAT3 activation. Hepatology 2004, 39, 1332–1342. [Google Scholar] [CrossRef]
- McHedlidze, T.; Waldner, M.; Zopf, S.; Walker, J.; Rankin, A.L.; Schuchmann, M.; Voehringer, D.; McKenzie, A.N.; Neurath, M.F.; Pflanz, S.; et al. Interleukin-33-dependent innate lymphoid cells mediate hepatic fibrosis. Immunity 2013, 39, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Shi, M.N.; Zheng, W.D.; Zhang, L.J.; Chen, Z.X.; Wang, X.Z. Therapeutic effect of interleukin-10 on CCl4-induced hepatic fibrosis in rats. World J. Gastroenterol. 2006, 12, 1386–1391. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Chen, Y.X.; Zhang, L.J.; Chen, Z.X.; Wang, X.Z. Hydrodynamics-based transfection of rat interleukin-10 gene attenuates porcine serum-induced liver fibrosis in rats by inhibiting the activation of hepatic stellate cells. Int. J. Mol. Med. 2014, 34, 677–686. [Google Scholar] [CrossRef]
- Rathinam, V.A.; Fitzgerald, K.A. Inflammasome Complexes: Emerging Mechanisms and Effector Functions. Cell 2016, 165, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Friedman, S.L. Mechanisms of Hepatic Stellate Cell Activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H. Cleavage of Gsdmd by Inflammatory Caspases Determines Pyroptotic Cell Death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Gan, C.; Cai, Q.; Tang, C.; Gao, J. Inflammasomes and Pyroptosis of Liver Cells in Liver Fibrosis. Front. Immunol. 2022, 30, 896473. [Google Scholar] [CrossRef]
- Pinzani, M.; Marra, F. Cytokine receptors and signaling in hepatic stellate cells. Semin. Liver Dis. 2001, 21, 397–416. [Google Scholar] [CrossRef]
- Corpechot, C.; Barbu, V.; Wendum, D.; Kinnman, N.; Rey, C.; Poupon, R.; Housset, C.; Rosmorduc, O. Hypoxia-induced VEGF and collagen I expressions are associated with angiogenesis and fibrogenesis in experimental cirrhosis. Hepatology 2002, 35, 1010–1021. [Google Scholar] [CrossRef]
- Moon, J.O.; Welch, T.P.; Gonzalez, F.J.; Copple, B.L. Reduced liver fibrosis in hypoxia-inducible factor-1alpha-deficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G582–G592. [Google Scholar] [CrossRef]
- Ajay, A.K.; Zhao, L.; Vig, S.; Fujiwara, M.; Thakurela, S.; Jadhav, S.; Cho, A.; Chiu, I.J.; Ding, Y.; Ramachandran, K.; et al. Deletion of STAT3 from Foxd1 cell population protects mice from kidney fibrosis by inhibiting pericytes trans-differentiation and migration. Cell Rep. 2022, 38, 110473. [Google Scholar] [CrossRef]
- Chen, W.; Yuan, H.; Cao, W.; Wang, T.; Chen, W.; Yu, H.; Fu, Y.; Jiang, B.; Zhou, H.; Guo, H.; et al. Blocking interleukin-6 trans-signaling protects against renal fibrosis by suppressing STAT3 activation. Theranostics 2019, 9, 3980–3991. [Google Scholar] [CrossRef]
- Maiyo, F.; Singh, M. Selenium nanoparticles: Potential in cancer gene and drug delivery. Nanomedicine (Lond) 2017, 12, 1075–1089. [Google Scholar] [CrossRef]
- Nath, D.; Kaur, L.; Sohal, H.S.; Malhi, D.S.; Garg, S.; Thakur, D. Application of Selenium Nanoparticles in Localized Drug Targeting for Cancer Therapy. Anticancer Agents Med Chem. 2022, 22, 2715–2725. [Google Scholar] [CrossRef] [PubMed]
- Goltyaev, M.V.; Varlamova, E.G.; Novoselov, S.V.; Fesenko, E.E. Activation of Signal Pathways of Apoptosis under Conditions of Prolonged ER-Stress Caused by Exposure of Mouse Testicular Teratoma Cells to Selenium-Containing Compounds. Dokl. Biochem. Biophys. 2020, 490, 9–11. [Google Scholar] [CrossRef]
- Goltyaev, M.V.; Mal’tseva, V.N.; Varlamova, E.G. Expression of ER-resident selenoproteins and activation of cancer cells apoptosis mechanisms under ER-stress conditions caused by methylseleninic acid. Gene 2020, 10, 144884. [Google Scholar] [CrossRef]
- Yoneda, T.; Imaizumi, K.; Oono, K.; Yui, D.; Gomi, F.; Katayama, T.; Tohyama, M. Activation of caspase-12, an endoplasmic reticulum (ER) resident caspase, through tumor necrosis factor receptorassociated factor 2-dependent mechanism in response to the ER-stress. J. Biol. Chem. 2001, 276, 13935–13940. [Google Scholar] [CrossRef] [PubMed]
- Niture, S.K.; Kaspar, J.W.; Shen, J.; Jaiswal, A.K. Nrf2 signaling and cell survival. Toxicol. Appl. Pharmacol. 2010, 244, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Zhu, H.; Morishima, N.; Li, E.; Xu, J.; Yankner, B.A.; Yuan, J. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-beta. Nature 2000, 403, 98–103. [Google Scholar] [CrossRef]
- Wang, X.Z.; Ron, D. Stress-induced phosphorylation and activation of the transcription factor CHOP (GADD153) by p38 MAP Kinase. Science 1996, 272, 1347–1349. [Google Scholar] [CrossRef] [PubMed]
- Senkal, C.E.; Ponnusamy, S.; Bielawski, J.; Hannun, Y.A.; Ogretmen, B. Antiapoptotic roles of ceramide-synthase-6-generated C16-ceramide via selective regulation of the ATF6/CHOP arm of ER-stress-response pathways. FASEB J. 2010, 24, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Xu, W.; Reed, J.C. Cell death and endoplasmic reticulum stress: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2010, 7, 1013–1030. [Google Scholar] [CrossRef] [PubMed]
- Bromati, C.R.; Lellis-Santos, C.; Yamanaka, T.S.; Nogueira, T.C.; Leonelli, M.; Caperuto, L.C. UPR induces transient burst of apoptosis in islets of early lactating rats through reduced AKT phosphorylation via ATF4/CHOP stimulation of TRB3 expression. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R92–R100. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Forward Primer 5′- > 3′ Reverse Primer 5′- > 3′ | Gene Names | Forward Primer 5′- > 3′ Reverse Primer 5′- > 3′ |
|---|---|---|---|
| TGF-β | ATGCAATGGGCTTAGTGTTCTG TCCTGTTGGCTGAGTTGTGAC | HIF-1α | GGCGACTGTGCACCTACTATG TGATCCAAAGCTCTGAGTAATTC |
| EGF | CCTTGGTTTGTGGTCCTAGAG CTGGGGTCCTCTGTCACTTG | XBP1s | AGTCCGCAGCACAGCAGGT AGAGAAAGGGAGGCTGGTAAG |
| Col1a1 | CATCACCTATCACTGCAAGAACAGGTCTTGGTGGTTTTGTTATTC | ATF-4 | TCGGGTTTGGGGGCTGAAG AAACAGAGCATCGAAGTCAAAC |
| Col1a2 | TCTCAGAACATCACCTACCACCACGGAATTCTTGGTCAGCAC | ATF-6 | AGGAGGGGAGATACGTTTTAC CGAGGAGCTTTTGATGTGGAG |
| Angpt1 | GGGACAGCAGGCAAACAGAG CTGGGCCCTTTGAAGTAGTG | CASP-1 | AGAGAAATGAAGTTGCTGCTGG ATCACCTTGGGCTTGTCTTTC |
| PDGF | ATTGAGATTGTGCGAAAGAAGC GGGGGCAATACAGCAAATACC | CASP-3 | CTCTTCATCATTCAGGCCTGC GACCCGTCCTTTGAATTTCTC |
| α-SMA | AGGAAGGATCTCTATGCTAACAAC ACTTAGAAGCATTTGCGGTGG | CASP-12 | TGTTGGTGTTATCATTTGGAGG TTTTCTTTTCTTCTCAGCTACAG |
| GFAP | CCGCCACCTGCAGGAGTAC TGTATTGTGAGCCTTTTGAGAG | BIM | AATGGCCGGCTATGGATGATG GCCAATTGGGTTCACTGTCTG |
| IL-17 | CCCTCAAAGCTCAGCGTGTC CCAGCTTTCCCTCCGCATTG | GADD34 | GAGTCCCATGAAGAGATTGTAC ACCAGCCCAGCAGCCACTTAG |
| IL-1β | CGTGCTGTCGGACCCATATG GCTCTTGACTTCTATCTTGTTG | PUMA | TGAAGATCTGCGCCGGGAG GAGAGGGACATGACGCGTG |
| IL-33 | TTTTGGAGAATGGATGTTATGTG TTTGTGAAGGACGAAGAAGGC | CHOP | CAGCTGGGAGCTGGAAGCCTG GACCACTCTGTTTCCGTTTCC |
| IL-22 | GCTCCCCCAGTCAGACAGG TAGAAGGCAGGAAGGAGCAG | BAX | TAAAGTGCCCGAGCTGATCAGAAC CTTCCCAGCCACCCTGGTCTT |
| IL-10 | AGCATGGCCCAGAAATCAAGG AGACTCAATACACACTGCAGG | BAK | CAGATGGATCGCACAGAGAG GCGTCTTTGCCCTGGGGAG |
| IL-6 | TCCAGAGATACAAAGAAATGATG TTGGAAATTGGGGTAGGAAGG | BCL-2 | AAGTCAACACAAACCCCAAGTCCTC GCAGATCTTCAGGTTCCTCCTGAGA |
| NFkB | TTAAAGAAACACTCAACAGCCAG TTCAGCACTCGCACGGACAC | BCL-XL | AGAGTGAGCCCAGCAGAACC GCAAGTTGGATGGCCACCTATC |
| STAT3 | CCCCGTACCTGAAGACCAAG ATGGGGTTCGGCTGCTTAGG | NRF-2 | CACATTGGGATTCACGCATAGGAGCACT TCCTGGACGGGACTATTGAAGGCTG |
| TNF-α | TGGAAAGACAGAGGGTGCAG TTGTCCCTTGAAGAGAACCTG | JNK1 | AGAAGCAGAAGCCCCACCAC ACTGCTGTCTGTATCCGAGG |
| INF-γ | GTGACATGAAAATCCTGCAGAG TGAGGCTGGATTCCGGCAAC | P53 | TGTTTAGGTCAAGGTGTCTCC GAACACAGCCCCTAACACAG |
| NOX1 | ACAAGAGATGGAGGAATTAGG TTCCTAGGATCCAGACTCGAG | P38 | GTCGACCTACTGGAGAAGATC AGTGAGATAGACAGAACAGAAAC |
| NOX4 | TACCTCAGTCAAACAGATGGG TGTCCCATATGAGTTGTTCCG | GAPDH | GTAAAGACCTCTATGCCAACAC GGTGCACGATGGAGGGGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varlamova, E.G.; Goltyaev, M.V.; Rogachev, V.V.; Gudkov, S.V.; Karaduleva, E.V.; Turovsky, E.A. Antifibrotic Effect of Selenium-Containing Nanoparticles on a Model of TAA-Induced Liver Fibrosis. Cells 2023, 12, 2723. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12232723
Varlamova EG, Goltyaev MV, Rogachev VV, Gudkov SV, Karaduleva EV, Turovsky EA. Antifibrotic Effect of Selenium-Containing Nanoparticles on a Model of TAA-Induced Liver Fibrosis. Cells. 2023; 12(23):2723. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12232723
Chicago/Turabian StyleVarlamova, Elena G., Michail Victorovich Goltyaev, Vladimir Vladimirovich Rogachev, Sergey V. Gudkov, Elena V. Karaduleva, and Egor A. Turovsky. 2023. "Antifibrotic Effect of Selenium-Containing Nanoparticles on a Model of TAA-Induced Liver Fibrosis" Cells 12, no. 23: 2723. https://0-doi-org.brum.beds.ac.uk/10.3390/cells12232723