Comparison of the RNA Content of Extracellular Vesicles Derived from Paracoccidioides brasiliensis and Paracoccidioides lutzii

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Growth Conditions
2.2. EVs Isolation
2.3. Murine Monocyte-Derived CD11c+ Cells (MoDC)
2.4. Indirect Co-Culture of Murine Monocyte-Derived CD11c+ Cells (MoDC) and Pb18
2.5. RNA Isolation and Sequencing
2.6. Data Analysis
2.7. Data Access
2.8. In Vitro Translation and Protein Analysis
3. Results
3.1. Paracoccidioides EVs Carry Functional mRNA
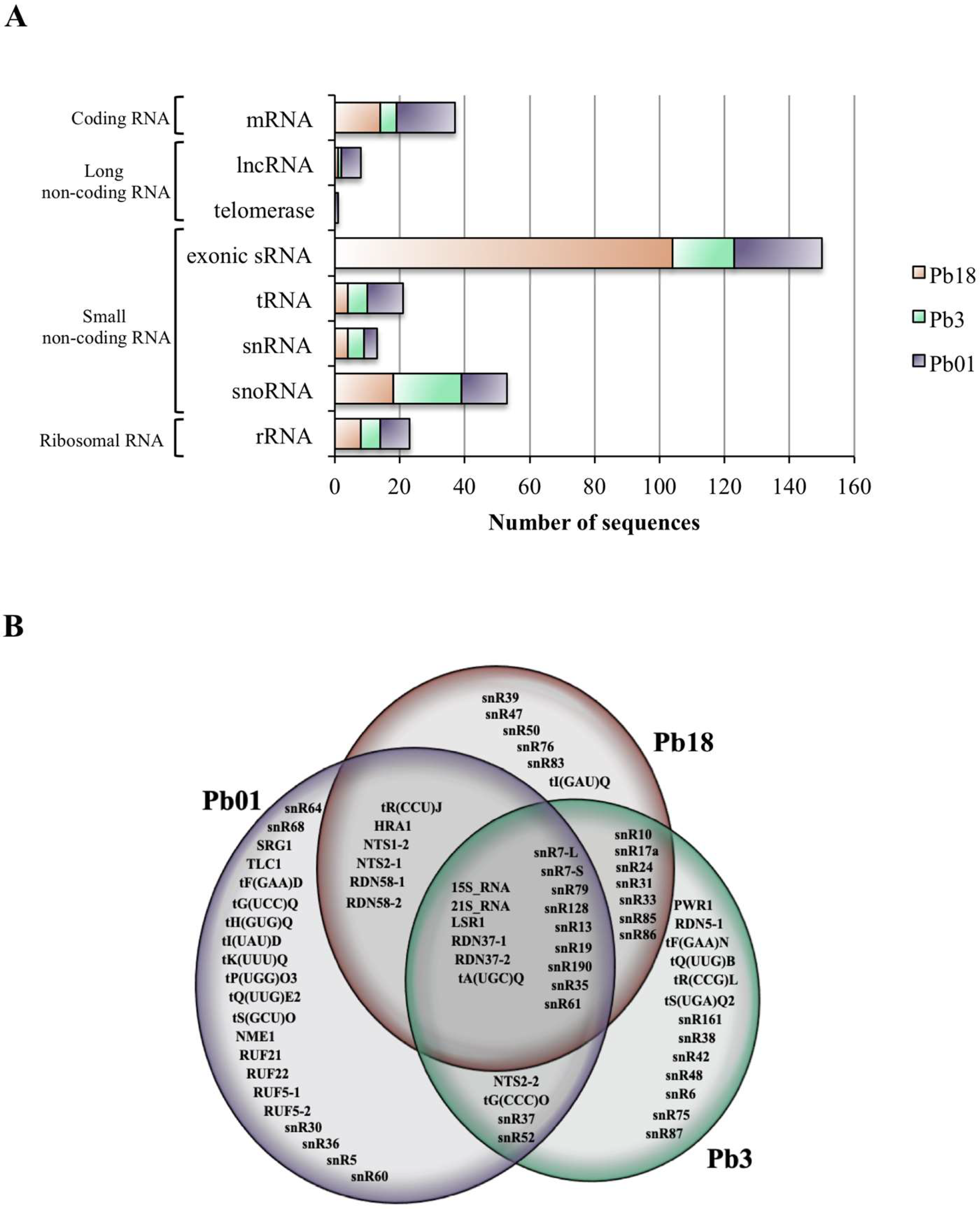
3.2. sRNA Sequences Aligning to mRNA Exons (Exonic sRNA)
3.3. Comparison of EV ncRNA Classes in Paracoccidioides EVs
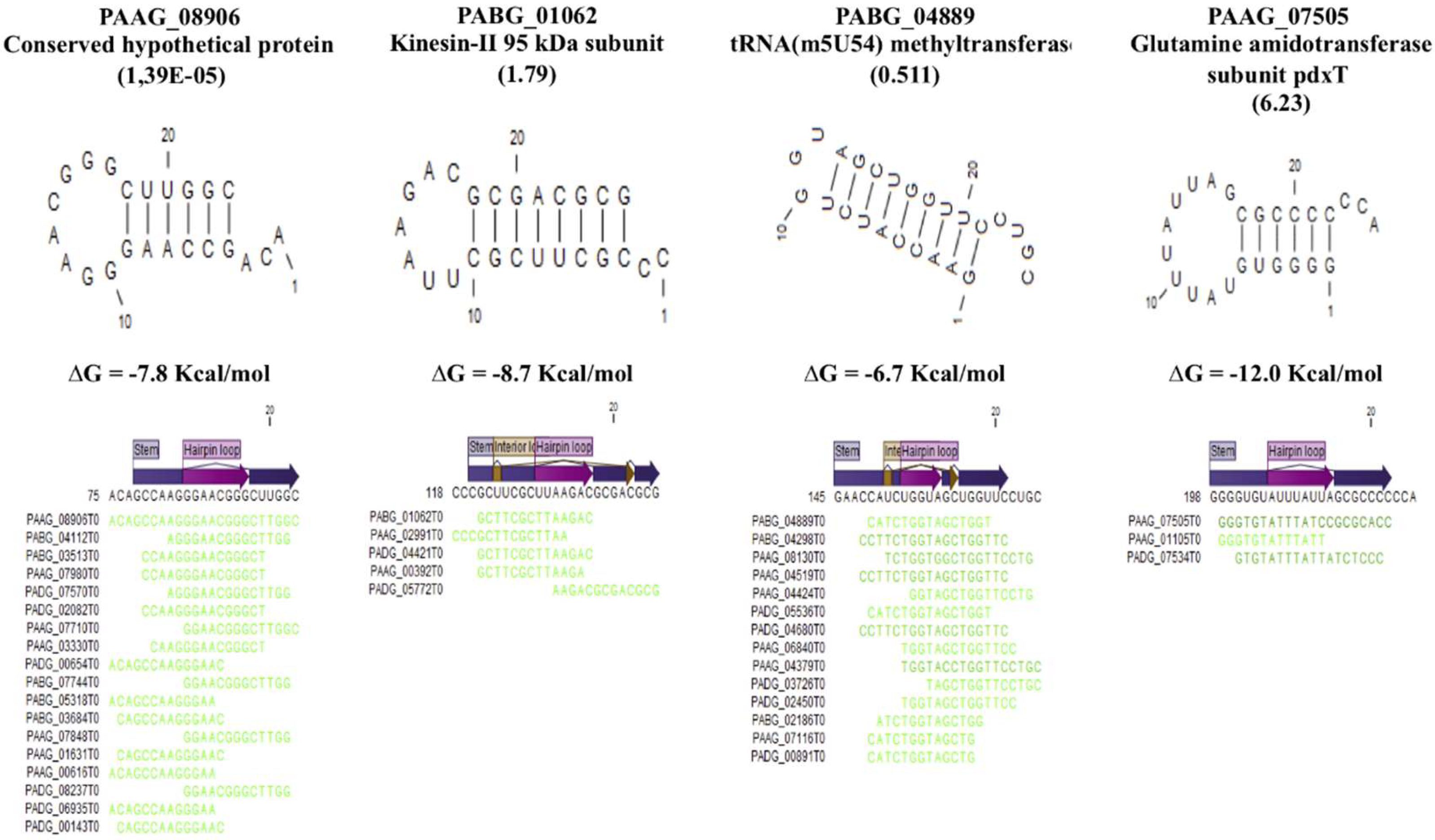
3.4. Secondary Structure in the EVs.
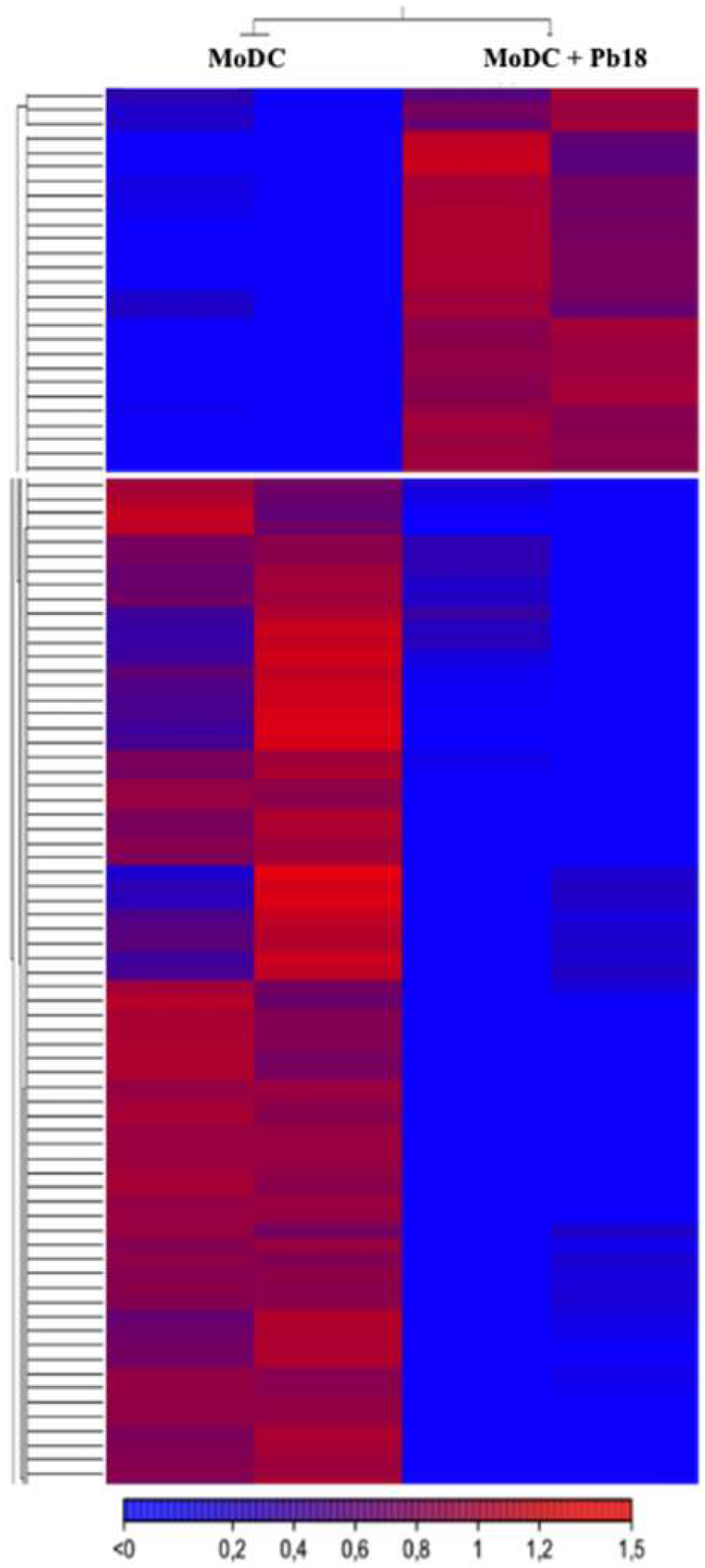
3.5. Paracoccidioides EVs Might Modulate the Transcriptome of Dendritic Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| EVs | Extracellular vesicles |
| GM-CSF | Granulocyte macrophage colony-stimulating factor |
| miRNA | Micro RNA |
| ncRNA | Non-coding RNA |
| PCM | paracoccidioidomycosis |
| sRNA | Small RNA |
| snoRNAs | Small nucleolar RNA |
| tRNA | Transporter RNA |
References
- Martinez, R. New trends in paracoccidioidomycosis epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Bocca, A.L.; Amaral, A.C.; Teixeira, M.M.; Sato, P.K.; Shikanai-Yasuda, M.A.; Soares Felipe, M.S. Paracoccidioidomycosis: Eco-epidemiology, taxonomy and clinical and therapeutic issues. Future Microbiol. 2013, 8, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.M.; Theodoro, R.C.; Nino-Vega, G.; Bagagli, E.; Felipe, M.S. Paracoccidioides species complex: Ecology, phylogeny, sexual reproduction, and virulence. PLoS Pathog. 2014, 10, e1004397. [Google Scholar] [CrossRef] [PubMed]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Matute, D.R.; Sepulveda, V.E.; Quesada, L.M.; Goldman, G.H.; Taylor, J.W.; Restrepo, A.; McEwen, J.G. Microsatellite analysis of three phylogenetic species of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2006, 44, 2153–2157. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, K.C.; Ganiko, L.; Batista, W.L.; Morais, F.V.; Marques, E.R.; Goldman, G.H.; Franco, M.F.; Puccia, R. Virulence of Paracoccidioides brasiliensis and gp43 expression in isolates bearing known pbgp43 genotype. Microbes Infect. 2005, 7, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Munoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.; et al. Genome diversity, recombination, and virulence across the major lineages of paracoccidioides. mSphere 2016, 1, e00213-16. [Google Scholar] [CrossRef] [PubMed]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: Artefacts no more. Trends Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Kalra, H.; Drummen, G.P.; Mathivanan, S. Focus on extracellular vesicles: Introducing the next small big thing. Int. J. Mol. Sci. 2016, 17, 170. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.L.; Franzen, A.J.; Nimrichter, L.; Miranda, K. Vesicular mechanisms of traffic of fungal molecules to the extracellular space. Curr. Opin. Microbiol. 2013, 16, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (misev2018): A position statement of the international society for extracellular vesicles and update of the misev2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Schorey, J.S.; Harding, C.V. Extracellular vesicles and infectious diseases: New complexity to an old story. J. Clin. Investig. 2016, 126, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Hosseini-Beheshti, E.; Grau, G.E.R. Extracellular vesicles as mediators of immunopathology in infectious diseases. Immunol. Cell Biol. 2018. [Google Scholar] [CrossRef]
- Volgers, C.; Benedikter, B.J.; Grauls, G.E.; Hellebrand, P.H.M.; Savelkoul, P.H.M.; Stassen, F.R.M. Effects of n-acetyl-l-cysteine on the membrane vesicle release and growth of respiratory pathogens. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Mateescu, B.; Kowal, E.J.; van Balkom, B.W.; Bartel, S.; Bhattacharyya, S.N.; Buzas, E.I.; Buck, A.H.; de Candia, P.; Chow, F.W.; Das, S.; et al. Obstacles and opportunities in the functional analysis of extracellular vesicle rna - an isev position paper. J. Extracell. Vesicles 2017, 6, 1286095. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mrnas and micrornas is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.; de Gruijl, T.D.; Wurdinger, T.; Middeldorp, J.M. Functional delivery of viral mirnas via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, R.; Van Roosbroeck, K.; Calin, G.A. Cell-to-cell communication: Micrornas as hormones. Mol. Oncol. 2017, 11, 1673–1686. [Google Scholar] [CrossRef] [PubMed]
- Fatima, F.; Nawaz, M. Vesiculated long non-coding rnas: Offshore packages deciphering trans-regulation between cells, cancer progression and resistance to therapies. Non-Coding RNA 2017, 3, 10. [Google Scholar] [CrossRef]
- Peres da Silva, R.; Puccia, R.; Rodrigues, M.L.; Oliveira, D.L.; Joffe, L.S.; Cesar, G.V.; Nimrichter, L.; Goldenberg, S.; Alves, L.R. Extracellular vesicle-mediated export of fungal rna. Sci. Rep. 2015, 5, 7763. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, M.C.; Matsuo, A.L.; Ganiko, L.; Medeiros, L.C.; Miranda, K.; Silva, L.S.; Freymuller-Haapalainen, E.; Sinigaglia-Coimbra, R.; Almeida, I.C.; Puccia, R. The pathogenic fungus Paracoccidioides brasiliensis exports extracellular vesicles containing highly immunogenic α-galactosyl epitopes. Eukaryot. Cell 2011, 10, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Sükösd, Z.; Knudsen, B.; Kjems, J.; Pedersen, C.N.S. Ppfold 3.0: Fast rna secondary structure prediction using phylogeny and auxiliary data. Bioinformatics 2012, 28, 2691–2692. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.M.; Fu, Z.; Dulloor, P.; Yen, T.; Barron-Casella, E.; Savage, W.; Van Eyk, J.E.; Casella, J.F.; Everett, A. Hemoglobin depletion from plasma: Considerations for proteomic discovery in sickle cell disease and other hemolytic processes. Proteom. Clin. Appl. 2010, 4, 926–930. [Google Scholar] [CrossRef]
- Nicolas, F.E.; Moxon, S.; de Haro, J.P.; Calo, S.; Grigoriev, I.V.; Torres-Martinez, S.; Moulton, V.; Ruiz-Vazquez, R.M.; Dalmay, T. Endogenous short rnas generated by dicer 2 and rna-dependent rna polymerase 1 regulate mrnas in the basal fungus Mucor circinelloides. Nucleic Acids Res. 2010, 38, 5535–5541. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.; Hube, B. Importance of the Candida albicans cell wall during commensalism and infection. Curr. Opin. Microbiol. 2012, 15, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Thind, S.K.; Taborda, C.P.; Nosanchuk, J.D. Dendritic cell interactions with Histoplasma and Paracoccidioides. Virulence 2015, 6, 424–432. [Google Scholar] [CrossRef]
- Alves, L.R.; Peres da Silva, R.; Sanchez, D.A.; Zamith-Miranda, D.; Rodrigues, M.L.; Goldenberg, S.; Puccia, R.; Nosanchuk, J.D. Extracellular vesicle-mediated rna release in Histoplasma capsulatum. mSphere 2019, 4, e00176-19. [Google Scholar] [CrossRef]
- Rappleye, C.A.; Eissenberg, L.G.; Goldman, W.E. Histoplasma capsulatum α-(1,3)-glucan blocks innate immune recognition by the beta-glucan receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 1366–1370. [Google Scholar] [CrossRef]
- Jiang, Y.; Dukik, K.; Muñoz, J.F.; Sigler, L.; Schwartz, I.S.; Govender, N.P.; Kenyon, C.; Feng, P.; van den Ende, B.G.; Stielow, J.B.; et al. Phylogeny, ecology and taxonomy of systemic pathogens and their relatives in Ajellomycetaceae (onygenales): Blastomyces, Emergomyces, Emmonsia, Emmonsiellopsis. Fungal Divers. 2018, 90, 245–291. [Google Scholar] [CrossRef]
- da Silva, T.A.; Roque-Barreira, M.C.; Casadevall, A.; Almeida, F. Extracellular vesicles from Paracoccidioides brasiliensis induced m1 polarization in vitro. Sci. Rep. 2016, 6, 35867. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, U.; Qazi, K.R.; Johansson, C.; Hultenby, K.; Karlsson, M.; Lundeberg, L.; Gabrielsson, S.; Scheynius, A. Nanovesicles from Malassezia sympodialis and host exosomes induce cytokine responses--novel mechanisms for host-microbe interactions in atopic eczema. PLoS ONE 2011, 6, e21480. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.L.; Freire-de-Lima, C.G.; Nosanchuk, J.D.; Casadevall, A.; Rodrigues, M.L.; Nimrichter, L. Extracellular vesicles from Cryptococcus neoformans modulate macrophage functions. Infect. Immun. 2010, 78, 1601–1609. [Google Scholar] [CrossRef] [PubMed]
- Vargas, G.; Rocha, J.D.; Oliveira, D.L.; Albuquerque, P.C.; Frases, S.; Santos, S.S.; Nosanchuk, J.D.; Gomes, A.M.; Medeiros, L.C.; Miranda, K.; et al. Compositional and immunobiological analyses of extracellular vesicles released by Candida albicans. Cell Microbiol. 2014. [Google Scholar] [CrossRef]
- Ikeda, M.A.K.; de Almeida, J.R.F.; Jannuzzi, G.P.; Cronemberger-Andrade, A.; Torrecilhas, A.C.T.; Moretti, N.S.; da Cunha, J.P.C.; de Almeida, S.R.; Ferreira, K.S. Extracellular vesicles from Sporothrix brasiliensis are an important virulence factor that induce an increase in fungal burden in experimental sporotrichosis. Front. Microbiol. 2018, 9, 2286. [Google Scholar] [CrossRef] [PubMed]
- Walker, L.; Sood, P.; Lenardon, M.D. The viscoelastic properties of the fungal cell wall allow traffic of ambisome as intact liposome vesicles. MBio 2018, 9. [Google Scholar] [CrossRef]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.M.; Palmquist, J.; Huang, S.D.; Jin, H. Plants send small rnas in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef]
- Bielska, E.; Sisquella, M.A.; Aldeieg, M.; Birch, C.; O’Donoghue, E.J.; May, R.C. Pathogen-derived extracellular vesicles mediate virulence in the fatal human pathogen Cryptococcus Gattii. Nat. Commun. 2018, 9, 1556. [Google Scholar] [CrossRef] [PubMed]
- Cleare, L.G.; Zamith-Miranda, D.; Nosanchuk, J.D. Heat shock proteins in Histoplasma and Paracoccidioides. Clin. Vaccine Immunol. 2017, 24, e00221-17. [Google Scholar] [CrossRef]
- Nicolas, F.E.; Ruiz-Vazquez, R.M. Functional diversity of rnai-associated srnas in fungi. Int. J. Mol. Sci. 2013, 14, 15348–15360. [Google Scholar] [CrossRef]
- Son, H.; Park, A.R.; Lim, J.Y.; Shin, C.; Lee, Y.W. Genome-wide exonic small interference rna-mediated gene silencing regulates sexual reproduction in the homothallic fungus Fusarium graminearum. PLoS Genet. 2017, 13, e1006595. [Google Scholar] [CrossRef]
- Camacho, E.; Sepulveda, V.E.; Goldman, W.E.; San-Blas, G.; Nino-Vega, G.A. Expression of paracoccidioides brasiliensis amy1 in a Histoplasma capsulatum amy1 mutant, relates an alpha-(1,4)-amylase to cell wall alpha-(1,3)-glucan synthesis. PLoS ONE 2012, 7, e50201. [Google Scholar] [CrossRef]
- Kumar, P.; Kuscu, C.; Dutta, A. Biogenesis and function of transfer rna-related fragments (trfs). Trends Biochem. Sci. 2016, 41, 679–689. [Google Scholar] [CrossRef]
- Garcia-Silva, M.R.; Cabrera-Cabrera, F.; das Neves, R.F.; Souto-Padron, T.; de Souza, W.; Cayota, A. Gene expression changes induced by Trypanosoma cruzi shed microvesicles in mammalian host cells: Relevance of trna-derived halves. Biomed Res. Int. 2014, 2014, 305239. [Google Scholar] [CrossRef]
- Jiang, N.; Yang, Y.; Janbon, G.; Pan, J.; Zhu, X. Identification and functional demonstration of mirnas in the fungus Cryptococcus neoformans. PLoS ONE 2012, 7, e52734. [Google Scholar] [CrossRef]
- Chen, R.; Jiang, N.; Jiang, Q.; Sun, X.; Wang, Y.; Zhang, H.; Hu, Z. Exploring microrna-like small rnas in the filamentous fungus Fusarium oxysporum. PLoS ONE 2014, 9, e104956. [Google Scholar] [CrossRef]
- Lau, S.K.; Chow, W.N.; Wong, A.Y.; Yeung, J.M.; Bao, J.; Zhang, N.; Lok, S.; Woo, P.C.; Yuen, K.Y. Identification of microrna-like rnas in mycelial and yeast phases of the thermal dimorphic fungus Penicillium marneffei. PLoS Negl. Trop. Dis. 2013, 7, e2398. [Google Scholar] [CrossRef]
- Lee, H.C.; Li, L.; Gu, W.; Xue, Z.; Crosthwaite, S.K.; Pertsemlidis, A.; Lewis, Z.A.; Freitag, M.; Selker, E.U.; Mello, C.C.; et al. Diverse pathways generate microrna-like rnas and dicer-independent small interfering rnas in fungi. Mol. Cell 2010, 38, 803–814. [Google Scholar] [CrossRef]
- Curcio, J.S.; Batista, M.P.; Paccez, J.D.; Novaes, E.; Soares, C.M.A. In silico characterization of micrornas-like sequences in the genome of Paracoccidioides brasiliensis. Genet. Mol. Biol. 2019, 42, 95–107. [Google Scholar] [CrossRef]
- Santhekadur, P.K.; Das, S.K.; Gredler, R.; Chen, D.; Srivastava, J.; Robertson, C.; Baldwin, A.S., Jr.; Fisher, P.B.; Sarkar, D. Multifunction protein staphylococcal nuclease domain containing 1 (snd1) promotes tumor angiogenesis in human hepatocellular carcinoma through novel pathway that involves nuclear factor kappab and mir-221. J. Biol. Chem. 2012, 287, 13952–13958. [Google Scholar] [CrossRef]
- Vallejo, M.C.; Nakayasu, E.S.; Matsuo, A.L.; Sobreira, T.J.; Longo, L.V.; Ganiko, L.; Almeida, I.C.; Puccia, R. Vesicle and vesicle-free extracellular proteome of Paracoccidioides brasiliensis: Comparative analysis with other pathogenic fungi. J. Proteome Res. 2012, 11, 1676–1685. [Google Scholar] [CrossRef]
- Chung, E.Y.; Liu, J.; Homma, Y.; Zhang, Y.; Brendolan, A.; Saggese, M.; Han, J.; Silverstein, R.; Selleri, L.; Ma, X. Interleukin-10 expression in macrophages during phagocytosis of apoptotic cells is mediated by homeodomain proteins pbx1 and prep-1. Immunity 2007, 27, 952–964. [Google Scholar] [CrossRef]
- Yu, S.; Zhao, D.M.; Jothi, R.; Xue, H.H. Critical requirement of gabpalpha for normal T cell development. J. Biol. Chem. 2010, 285, 10179–10188. [Google Scholar] [CrossRef]
- Unsinger, J.; Burnham, C.A.; McDonough, J.; Morre, M.; Prakash, P.S.; Caldwell, C.C.; Dunne, W.M., Jr.; Hotchkiss, R.S. Interleukin-7 ameliorates immune dysfunction and improves survival in a 2-hit model of fungal sepsis. J. Infect. Dis. 2012, 206, 606–616. [Google Scholar] [CrossRef]
- Ruan, S.; Samuelson, D.R.; Assouline, B.; Morre, M.; Shellito, J.E. Treatment with interleukin-7 restores host defense against Pneumocystis in cd4+ t-lymphocyte-depleted mice. Infect. Immun. 2016, 84, 108–119. [Google Scholar] [CrossRef]
- Calich, V.L.; da Costa, T.A.; Felonato, M.; Arruda, C.; Bernardino, S.; Loures, F.V.; Ribeiro, L.R.; de Cassia Valente-Ferreira, R.; Pina, A. Innate immunity to Paracoccidioides brasiliensis infection. Mycopathologia 2008, 165, 223–236. [Google Scholar] [CrossRef]
- Peres da Silva, R.; Heiss, C.; Black, I.; Azadi, P.; Gerlach, J.Q.; Travassos, L.R.; Joshi, L.; Kilcoyne, M.; Puccia, R. Extracellular vesicles from Paracoccidioides pathogenic species transport polysaccharide and expose ligands for dc-sign receptors. Sci. Rep. 2015, 5, 14213. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Pb18 Feature ID | Pb3 Feature ID | Pb01 Feature ID | Sequence Description | Coverage | GO |
|---|---|---|---|---|---|
| Protein Modification | |||||
| PAAG_08003 | hsp70-like protein | 100% | ion binding | ||
| PAAG_05980 | ubiquitin-conjugating enzyme e2-16 kda | 80% | ligase activity | ||
| PADG_07715 | hsp90-like protein | 70% | protein folding, response to stress | ||
| PADG_01605 | PABG_03078 | PAAG_07080 | polyubiquitin | 70% | protein modification process |
| PADG_11111 | nuclear transport factor 2 | 75% | protein targeting | ||
| PAAG_05679 | ATP-dependent molecular chaperone hsc82 | 90% | nucleic acid binding transcription factor activity, protein folding | ||
| Carbohydrate Metabolism | |||||
| PADG_02145 | glycogen phosphorylase | 70% | carbohydrate metabolic process | ||
| Translation | |||||
| PADG_05025 | 60s ribosomal protein l26 | 70% | translation | ||
| PADG_00692 | PAAG_11418 | elongation factor 1-alpha | 100% | translation | |
| PADG_03326 | 40s ribosomal protein s9 | 80% | translation | ||
| PABG_05744 | pre-mrna splicing factor | 70% | translation | ||
| Oxidation-Reduction | |||||
| PAAG_03216 | thiol-specific antioxidant | 90% | oxidoreductase activity | ||
| Transport | |||||
| PAAG_11262 | hsp7-like protein | 100% + MR | transmembrane transport | ||
| PAAG_03058 | high-affinity methionine permease | 100% + 5′R | transmembrane transport | ||
| PAAG_07634 | gtp-binding protein rhoa | 70% | vesicle-mediated transport | ||
| DNA Metabolism or Biogenesis | |||||
| PADG_00873 | PABG_02444 | PAAG_07099 | histone h3 | 100% | chromosome organization |
| PABG_05588 | PAAG_08917 | histone h2a | 90% | chromosome organization | |
| PAAG_08918 | histone h2b | 70% | chromosome organization | ||
| PADG_06568 | tctp family protein | 50% | cell differentiation | ||
| PABG_03449 | PAAG_08247 | calmodulin | 70% | ion binding | |
| Other/Unknown Function | |||||
| PADG_02280 | hypothetical protein | 75% | Unknown | ||
| PADG_02399 | calcium-binding protein | 70% | Unknown | ||
| PADG_04049 | hypothetical protein | 60% | Unknown | ||
| PADG_08402 | hypothetical protein | 100% | Unknown | ||
| PADG_12385 | ser thr protein phosphatase family protein | 60% | Unknown | ||
| PAAG_00340 | conserved hypothetical protein | 85% | unknown | ||
| PAAG_12435 | hypothetical protein | 80% | unknown | ||
| PAAG_12692 | ATP synthase subunit beta | 95% reverse | unknown | ||
| PAAG_12694 | plant senescence-associated protein | 100% reverse | unknown | ||
| PAAG_02087 | kelch-like protein 38 | 70% | unknown | ||
| Feature ID | Name | RPKM Mean |
|---|---|---|
| PADG_00648 | conserved hypothetical protein | 7 |
| PADG_04056 | 14-3-3 protein epsilon | 25 |
| PADG_04810 | GTP-binding nuclear protein GSP1/Ran | 37 |
| PADG_06159 | sulfate transporter | 12 |
| PADG_07715 | Hsp90-like protein | 127 |
| Pb18 Feature ID | Alignment | Pb3 Feature ID | Alignment | Pb01 Feature ID | Alignment | Sequence Description | GO |
|---|---|---|---|---|---|---|---|
| Protein modification | |||||||
| PADG_01365 | 5′R/3′F | disulfide isomerase | protein folding | ||||
| PADG_03114 | MR | phospho-2-dehydro-3-deoxyheptonate aldolase | amino acid metabolic process | ||||
| PADG_04092 | 3′F | peptidyl-prolyl cis-trans isomerase b | protein modification process | ||||
| PADG_05011 | MF | peptidyl-prolyl cis-trans isomerase-like 3 | protein modification process | ||||
| PADG_05560 | 3′F | 26s proteasome regulatory subunit rpn-1 | small molecule metabolic process | ||||
| PADG_05731 | 3′F | hypothetical protein | amino acid metabolic process | ||||
| PADG_07241 | 5′F | dihydroxy-acid dehydratase | amino acid metabolic process | ||||
| PADG_07550 | MR | PABG_04093 | MR | microsomal signal peptidase subunit | peptidase activity | ||
| PAAG_05962 | 5′F | proteasome regulatory particle subunit | catabolic process | ||||
| Carbohydrate Metabolism | |||||||
| PADG_02145 | 5′F | PABG_06801 | 5′F | glycogen phosphorylase | carbohydrate metabolic process | ||
| PADG_03169 | MR | alpha-glucan synthase ags2 | carbohydrate metabolic process | ||||
| PADG_04432 | MF | alpha-amylase | carbohydrate metabolic process | ||||
| PADG_04922 | 3′F | cell wall glucanase | carbohydrate metabolic process | ||||
| PADG_05870 | 5′R/5′F | glucan synthesis regulatory protein | carbohydrate metabolic process | ||||
| Lipid Metabolism | |||||||
| PADG_12430 | 5′F | PABG_07295 | 3′F | amp-binding | long-chain fatty acid metabolic process | ||
| Oxidation-Reduction | |||||||
| PADG_04419 | 3′R | PABG_01064 | 3′R | proline oxidase | catabolic process | ||
| PAAG_05378 | MF | d-3-phosphoglycerate dehydrogenase | oxidoreductase activity | ||||
| PAAG_01937 | 3′F | duf887 domain-containing protein | biosynthetic process | ||||
| PAAG_11262 | mRNA/MR | hsp7-like protein | transmembrane transpor | ||||
| PADG_05080 | 5′F | pyridoxamine phosphate oxidase family protein | ion binding | ||||
| PADG_06181 | 3′F | c-5 sterol desaturase | lipid metabolic process | ||||
| PADG_07431 | 5′R/3′F/3R | chloroperoxidase-like protein | homeostatic process | ||||
| PADG_12214 | 5′R | alcohol dehydrogenase | oxidoreductase activity | ||||
| Translation | |||||||
| PADG_00995 | MF | ubiquitin-40s ribosomal protein s27a | translation | ||||
| PADG_02452 | MF | rna polymerase rpb1 c-terminal repeat domain-containing protein | |||||
| PADG_02484 | MF | valyl-trna synthetase | tRNA metabolic process | ||||
| PADG_05025 | MF | 60s ribosomal protein l26 | translation | ||||
| PADG_06082 | 3′F | pre-mrna splicing factor | translation | ||||
| PADG_06160 | MF | eukaryotic translation initiation factor 2 alpha subunit | translation | ||||
| PADG_06191 | 5′F | PABG_06964 | 3′R | trna isopentenyltransferase | tRNA metabolic process | ||
| PADG_06522 | 5′F | glycine--trna ligase | |||||
| PADG_06833 | 5′F | ATP-dependent rna helicase drs1 | ribosome biogenesis | ||||
| PADG_08605 | 5′F | 40s ribosomal protein s28 | translation | ||||
| PADG_01891 | MF | translation initiation factor rli1 | translation | ||||
| PADG_02317 | 3′R | translation machinery-associated protein 17 | |||||
| Signaling Process | |||||||
| PADG_05447 | 3′F | vacuolar membrane-associated protein iml1 | signal transduction | ||||
| PADG_06642 | 5′F | ste ste7 protein kinase | response to stress | ||||
| PADG_08337* | 3′F | gtp-binding protein rhoa | signal transduction | ||||
| Transport | |||||||
| PADG_00326 | MF | adp-ribosylation factor-like protein 1 | vesicle-mediated transport | ||||
| PADG_01303 | 3′F | abc transporter | biosynthetic process | ||||
| PADG_01567 | 3′R | sorting nexin 3 | vesicle-mediated transport | ||||
| PADG_05084 | 5′R | high affinity copper transporter | transmembrane transport | ||||
| PADG_05821 | 5′F | importin | |||||
| PADG_06982 | 3′F | ncs1 family nucleobase:cation symporter-1 | transmembrane transport | ||||
| PADG_08101 | 3′F | mrna cleavage factor complex component pcf11 | transport | ||||
| PADG_03535 | MR | PABG_01859 | MR | PAAG_01288 | MR | nucleotide binding | ion binding, ligase activity |
| PABG_11660 | 3′F | endoplasmic reticulum vesicle protein 25 | vesicle-mediated transport | ||||
| PAAG_03058 | mRNA/5′R | high-affinity methionine permease | transmembrane transport | ||||
| PAAG_03479* | 3′R | mfs multidrug | transmembrane transport | ||||
| PAAG_11682 | 3′R | duf1903-domain-containing protein | transport | ||||
| PAAG_12134 | 3′F | mfs drug transporter | transmembrane transport | ||||
| DNA Metabolism or Biogenesis | |||||||
| PADG_00916 | MR | PABG_02494 | 5′R | PAAG_07153 | 5′R | transcription factor tfiiib complex subunit brf1 | biosynthetic process |
| PADG_03251 | 3′F | c6 finger domain protein acr- | biosynthetic process | ||||
| PADG_05475 | 5′R | dna-directed rna polymerases and iii 145 kda polypeptide | biosynthetic process | ||||
| PADG_06799 | MR | camk camkl kin4 protein kinase | cell division | ||||
| PADG_07652* | MR | PABG_06307* | MR | PAAG_05737* | MR | calcium calmodulin-dependent protein kinase | cytoskeleton organization, cell division |
| PADG_11268 | 3′F | tyrosine recombinase -like | DNA metabolic process | ||||
| PADG_11500 | MF | fungal specific transcription | biosynthetic process | ||||
| PADG_12343 | 3′F | serine threonine protein kinase | regulation of transcription | ||||
| PABG_00984 | 5′F | PAAG_03968 | 5′F | c6 transcription | biosynthetic process | ||
| PABG_03356 | 3′R | homeobox transcription | biosynthetic process | ||||
| Other/Unknown Function | |||||||
| PADG_00069 | 3′R | hypothetical protein | |||||
| PADG_00138 | 5′R | hypothetical protein | |||||
| PADG_00639 | 5′R/5′F | hypothetical protein | |||||
| PADG_01127 | 5′R | rna-binding protein | ion binding | ||||
| PADG_01198 | 3′F | vps9 domain | |||||
| PADG_01219 | MF/MR | hypothetical protein | |||||
| PADG_01476 | 3′F | PAAG_06944 | 3′F | c2h2 finger domain | ion binding | ||
| PADG_01739 | 3′R | ||||||
| PADG_01808 | MF | hypothetical protein | |||||
| PADG_01880 | 3′F | u-box domain-containing protein | |||||
| PADG_01892 | 3′R | formin binding protein | |||||
| PADG_02119 | MR | PABG_03545 | MR | protein | |||
| PADG_02181 | 5′R | had superfamily | isomerase activity | ||||
| PADG_02764 | 5′F | disulfide bond formation protein d | |||||
| PADG_02871 | 3′R | cfem domain-containing protein | |||||
| PADG_02926 | 3′R | tam domain methyltransferase | |||||
| PADG_03103 | 3′R | phytase | phosphatase activity | ||||
| PADG_03162 | 3′R | domain protein | |||||
| PADG_03436 | MF | 3 exoribonuclease family protein | |||||
| PADG_03788 | MF | polyadenylation factor subunit 64 | |||||
| PADG_04049 | 5′R/5′F | hypothetical protein | |||||
| PADG_04157 | 5′R | cellobiose dehydrogenase | |||||
| PADG_04417 | 3′F | hypothetical protein | |||||
| PADG_04448 | 5′F | polarized growth protein | |||||
| PADG_04473 | MR | duf647 domain-containing protein | |||||
| PADG_04629 | 3′R | protein | |||||
| PADG_04760 | 5′F | multiple myeloma tumor-associated protein 2 like | |||||
| PADG_04828 | MF/MR | adenylosuccinate lyase | biosynthetic process | ||||
| PADG_05226 | MF | protein | |||||
| PADG_05352 | 5′R | ubiquitin carboxyl-terminal hydrolase 19 | |||||
| PADG_05378 | 5′F | protein | |||||
| PADG_05589 | MR | protein | |||||
| PADG_05603 | 3′R | increased rdna silencing protein 4 | ion binding | ||||
| PADG_06044 | 3′F | ankyrin repeat containing protein | |||||
| PADG_06240 | 3′F | hypothetical protein | |||||
| PADG_06449 | 3′R | phosphotransferase enzyme family protein | |||||
| PADG_07205 | 5′F | protein | |||||
| PADG_07675 | MF/MR | cellular morphogenesis protein | |||||
| PADG_07897 | 5′R/5′F/5′NS | hypothetical protein | |||||
| PADG_07988 | 3′F/3′R | conserved hypothetical portein | |||||
| PADG_07990 | MF | tam domain methyltransferase | |||||
| PADG_08617 | 3′F/3′ R | PABG_07734 | 3′R | hypothetical protein | |||
| PADG_11034 | 5′R/5′F | PABG_11827 | 5′R/5′F | protein | |||
| PADG_11035 | 3′F | protein | |||||
| PADG_11277 | 5′R | protein | |||||
| PADG_11439 | 5′R | PABG_00126 | 5′R | PAAG_11926 | 5′R | hypothetical protein | |
| PADG_11473 | 5′R | hypothetical protein | |||||
| PADG_11562 | MF | hypothetical protein | |||||
| PADG_11613 | 5′R | hypothetical protein | |||||
| PADG_11652 | 3′R | PABG_01675 | 3′R | kh domain rna binding protein | RNA binding | ||
| PADG_11758 | 3′R/3′F/3′NS | hypothetical protein | |||||
| PADG_11762 | MF | hypothetical protein | |||||
| PADG_12001 | 3′F | hypothetical protein | |||||
| PADG_12385 | 5′R/5′F | PABG_06506 | 3′R | ser thr protein phosphatase family protein | |||
| PABG_06943 | 3′R | hypothetical protein | |||||
| PABG_12403 | MR/MF/MNS | hypothetical protein | |||||
| PAAG_01376 | 5′R | hypothetical protein | |||||
| PAAG_01424 | 5′F/MF | iron-sulfur cluster assembly accessory protein | biosynthetic process | ||||
| PAAG_01967 | MR | hypothetical protein | |||||
| PAAG_03361 | MF | predicted protein | |||||
| PAAG_04613 | MR | vacuolar protein sorting-associated protein 51 | |||||
| PAAG_05089 | 3′F | ||||||
| PAAG_07600 | 3′F | ||||||
| PAAG_07877 | 3′F | protein | |||||
| PAAG_11750 | 5′F/5′R | hypothetical protein | |||||
| PAAG_12291 | 3′R | camp-dependent protein kinase pathway protein | kinase activity | ||||
| PAAG_12405 | MR | hypothetical protein | |||||
| PAAG_12534 | 5′R | mitochondrial 37s ribosomal protein nam9 | rRNA binding | ||||
| PAAG_12681 | MR | hypothetical protein | ion binding | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peres da Silva, R.; Longo, L.G.V.; Cunha, J.P.C.d.; Sobreira, T.J.P.; Rodrigues, M.L.; Faoro, H.; Goldenberg, S.; Alves, L.R.; Puccia, R. Comparison of the RNA Content of Extracellular Vesicles Derived from Paracoccidioides brasiliensis and Paracoccidioides lutzii. Cells 2019, 8, 765. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8070765
Peres da Silva R, Longo LGV, Cunha JPCd, Sobreira TJP, Rodrigues ML, Faoro H, Goldenberg S, Alves LR, Puccia R. Comparison of the RNA Content of Extracellular Vesicles Derived from Paracoccidioides brasiliensis and Paracoccidioides lutzii. Cells. 2019; 8(7):765. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8070765
Chicago/Turabian StylePeres da Silva, Roberta, Larissa G. V. Longo, Julia P. C. da Cunha, Tiago J. P. Sobreira, Marcio L. Rodrigues, Helisson Faoro, Samuel Goldenberg, Lysangela R. Alves, and Rosana Puccia. 2019. "Comparison of the RNA Content of Extracellular Vesicles Derived from Paracoccidioides brasiliensis and Paracoccidioides lutzii" Cells 8, no. 7: 765. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8070765