1. Introduction
Breast cancer is endocrine-related cancer with high incidence and mortality rates in women worldwide. Over the past decades, the incidence rate of breast cancer has increased continuously despite an advance in early detection techniques and an awareness of breast cancer. Based on the molecular subtype, four different types of breast cancer have been identified: (1) Luminal A (HR+/HER2−) (71%), (2) luminal B (HR+/HER2+) (12%), (3) HER2-enriched (HR-/HER2+) (5%), and (4) triple-negative (HR−/HER2−) (12%) [
1]. Triple-negative breast cancer (TNBC) is responsible for 12%–15% of all breast cancer cases. The lack of estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor (HER2) in TNBC makes it highly aggressive and resistant to hormonal therapy [
2,
3]. The tendency of TNBC to metastasize to distinct organs is higher compared to other cancer phenotypes. Once cells get detached from the invasive front of TNBC, they migrate to distinct tissue sites through a capillary and proliferate in the new microenvironment [
4]. TNBC is more sensitive to chemotherapy compared to other types of breast cancer. However, even after a series of therapies (a traditional radiotherapy, an adjuvant chemotherapy, a systematic therapy, and a targeted therapy), the peak risk of TNBC recurrence takes place within 3–5 years, ultimately leading to death in most of the cases [
5,
6]. Therefore, to ease the burden of TNBC treatment, an optimal therapeutic approach should be found.
Wnt signaling is an essential cellular process associated with cell fate (development, differentiation, and proliferation), but it is inappropriately regulated in multiple types of cancer [
7,
8]. Currently, the Wnt signaling pathway is characterized into β-catenin-dependent Wnt signaling (canonical pathway), β-catenin-independent Wnt signaling (non-canonical pathway), and the noncanonical Wnt/calcium pathway. All three pathways are triggered by a Wnt protein-ligand binding to a Frizzled family receptor, leading to the activation of downstream proteins. The main function of the canonical Wnt pathway is to stabilize β-catenin and enhance its nuclear translocation. Next, inside the nucleus, the transcription factor LEF/TCF (lymphoid enhancer-binding factor1/T-cell factor) is activated by active β-catenin, which in turn activates the target genes such as c-Myc, cyclin D, Axin2, and Nkd1 [
9,
10,
11]. In the case of a lack of Wnt, β-catenin is degraded by the tumor suppressors such as adenomatous polyposis coli (APC), axis inhibitor (Axin), and glycogen synthase kinase 3 (GSK3), and a low level of β-catenin is maintained.
In breast cancer, the Wnt signaling pathway is aberrantly regulated and both canonical and noncanonical pathways are involved in TNBC tumorigenesis [
11,
12]. The mutation in APC and β-catenin is rare in breast cancer, but TNBC tumorigenesis is thought to be promoted by an elevated level of β-catenin [
13,
14]. Mouse cancer model studies showed that accumulation of β-catenin in the nucleus was correlated with cell migration, colony formation, stem-like features, and chemoresistance in TNBC, thus suggesting that Wnt/ β-catenin activation in cells could lead to TNBC tumorigenesis [
11]. The Wnt signaling destruction complex consists of Axin, the serine–threonine kinases casein kinase 1α/β (CK1), glycogen synthase kinase 3α/β (GSK3), and adenomatous polyposis coli (APC), which can play an important role in degrading β-catenin by phosphorylation [
15]. Abnormal assembly of the destruction complex prevents the phosphorylation of β-catenin, resulting in the accumulation of β-catenin in the cytosol and the translocation of β-catenin to the nucleus [
16,
17]. The β-catenin destruction complex is highly regulated by the Axin concentration, and the degradation of β-catenin increases in a cell line with Axin overexpression, meaning that the Axin level is strictly controlled to maintain Wnt signaling. Axin levels are controlled by many processes; in fact, Wnt signaling itself regulates the Axin expression by promoting Axin degradation [
18,
19]. Thus, it is considered that an antagonist against Wnt signaling is an attractive tool for cancer treatment.
XAV939 (a small molecule inhibitor of Tankyrase) was found to successfully inhibit Tankyrase activity and stabilize Axin in MDA-MB-231 breast cancer cells. Treatment with XAV939 alone effectively attenuated the Wnt signaling in serum-deprived breast cancer cells. Further, XAV939 also blocked the cell migration and suppressed the cell growth in the serum-deprived medium. However, XAV939 alone was not so effective in treating TNBC [
20].
Paclitaxel is a well-established drug for the treatment of breast cancer. Due to its side effects at a high dose, it is important to reduce the dose of paclitaxel while maintaining its anticancer effect. In our study, we aimed to examine a potential therapeutic effect of a low dose of paclitaxel combined with XAV939 on various breast cancer lines (MDA-MB-231, MDA-MB 468, BT549, MCF-7, and T-47D), MDA-MB-231-xenografted mice, and pristane-induced breast tumor. Here, we report that the combined treatment with XAV939 and a low dose of paclitaxel could treat TNBC effectively and synergistically.
2. Material and Methods
2.1. Reagents
XAV939 was purchased from Selleckchem (Houston, TX, USA), paclitaxel and pristane were purchased from Aladdin (Shanghai, China). Dulbecco’s modified Eagle’s medium (DMEM) and Roswell Park Memorial Institute (RPMI) 1640 medium were purchased from Gibco (ThermoFisher, Gaisburg, MD, USA). Fetal bovine serum (FBS) and penicillin streptomycin were purchased from (P&S, Gibco). Dimethyl sulfoxide (DMSO) was purchased from MP Biomedicals (Solon, OH, USA), crystal violate was purchased from KeyGen BioTECH (Nanjing, China). MTT cell proliferation and cytotoxicity assay kit were purchased from Phygene Life Sciences (Fuzhou, China). Haematoxylin and eosin (H&E) staining kit was purchased from Servicebio, (Wuhan, China). Tunel assay kit was purchased from Roche (Indianapolis, IN, USA).
The primary antibody against β-catenin (D10A8) XP®, Axin1 (C7B12), Cleaved PARP (Asp214) (D64E10) XP®, Cleaved Caspase-3 (Asp175) (9664), and Bcl-2 (124) (15071) were purchased from Cell Signaling Technology (Danvers, MA, USA). Primary antibodies for IHC anti-MMP9 (GB12132), anti-vimentin (GB11192), anti-E-cadherin (GB11082), anti-Ki-67 (GB13030) and anti-β-catenin (GB12015) were purchased from Serveicebio (Woburn, MA, USA). Secondary antibodies against anti-mouse and anti-rabbit were purchased from (Beijing Zhongshang Jinqiao Biotechnology Co., Ltd. Beijing, China).
2.2. Cell Culture
Human breast cancer cell lines (MDA-MB-231, MDA-MB-468, BT549, MCF-7, and T-47D) were obtained from Procell (Wuhan, China). Cells were maintained in DMEM and RPMI 1640 supplemented with 10% FBS and 1% penicillin-streptomycin (100 U/mL penicillin and 100 mg/mL streptomycin) in an incubator with a 5% CO2 humidified atmosphere at 37 °C.
2.3. Cytotoxicity Assay
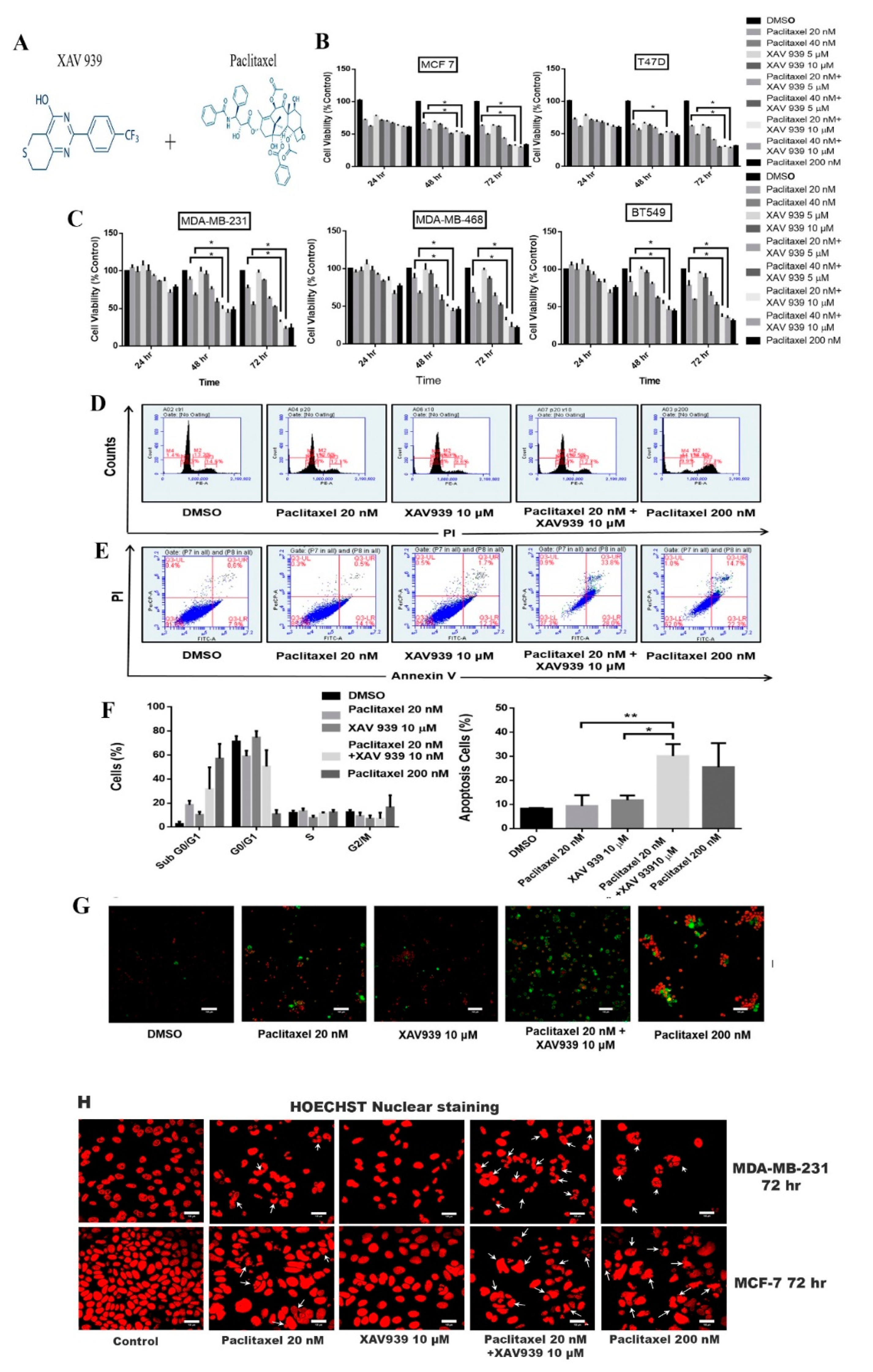
To assess the cytotoxicity effect of paclitaxel and/or XAV939 on MDA-MB-231, MDA-MB-468, BT549, MCF-7, and T-47D cells, the colorimetric MTT assay was performed as per the manufacturer’s instructions. Briefly, 8000 cells were seeded in a 96-well plate and treated with DMSO (3 µL/mL), different concentrations of paclitaxel (20 nM, 40 nM, and 200 nM) and XAV939 (5 µM and 10 µM), and a combination of paclitaxel and XAV939 (20 nM + 5 µM, 20 nM + 10 µM, 40 nM + 5 µM, and 40 nM + 10 µM) for 24 h, 48 h, and 72 h. After incubation, 110 µL of the MTT solution was added and incubated for 4 h. The purple precipitate was dissolved by adding 100 µL isopropanol and then measured at 570 nm using an EnSpire Multimode Plate Reader (PerkinElmer, Waltham, MA, USA).
2.4. Cell Cycle by Flow Cytometery
Briefly, MDA-MB-231 cells (1 × 106) were seeded in 6-well plates and after 24 h cells were treated with DMSO (3 µL/mL), paclitaxel (20 nM and 200 nM), XAV939 (10 µM), and paclitaxel + XAV939 (20 nM + 10 µM) for 48 h. After trypsinization, cells were fixed with 70% ethanol and incubated at 4 °C for 30 min. Next, cells were washed and suspended in PBS containing 0.5 mg/mL ribonuclease A (Sigma-Aldrich, St. Louis, MO, USA) and incubated for 30 min at 37 °C. Cells were stained with 50 μg/mL propidium iodide (Sigma-Aldrich, St. Louis, MO, USA) and analyzed with BD Accuri C6 Plus (BD) at an excitation wavelength of 488 nm and an emission wavelength of 630 nm. The cell cycle distribution was quantified by using BD Accuri C6 Plus software.
2.5. Annexin V/Propidium Iodide Staining
Annexin V/propidium iodide staining for apoptosis was conducted using Annexin V-FITC Detection kit (DojinDo, Kumamoto, Japan) according to the manufacturer’s instructions. Briefly, MDA-MB-231 cells (1 × 106) were seeded in a 6-well plate, and after 24 h cells were treated with DMSO (3 µL/mL), Paclitaxel (20 nM), XAV939 (10 µM), paclitaxel + XAV939 (20 nM + 10 µM), and paclitaxel (200 nM). After 48 h, cells were trypsinized, single-cell suspensions were prepared with 5 μL of annexin V-FITC and subsequently with a 5 μL PI solution in the dark, and cell suspensions were incubated for 15 min. Then, a 400 μL annexin V Binding solution was added to each cell suspension, and each cell solution was analyzed by flow cytometry. Necrotic, early, and late apoptosis cells were identified by BD Accuri C6 Plus software (Version 227, BD Biosciences, San Jose, CA, USA). For microscopic observation, cells were deposited on slides, air dried, and the cover slip was mounted with a drop of the antifluorescent quencher (Beyotime, Shanghai, China) and observed under a confocal microscope Nikon Ti-E-A1 (Tokyo, Japan).
2.6. Wound Healing Assay
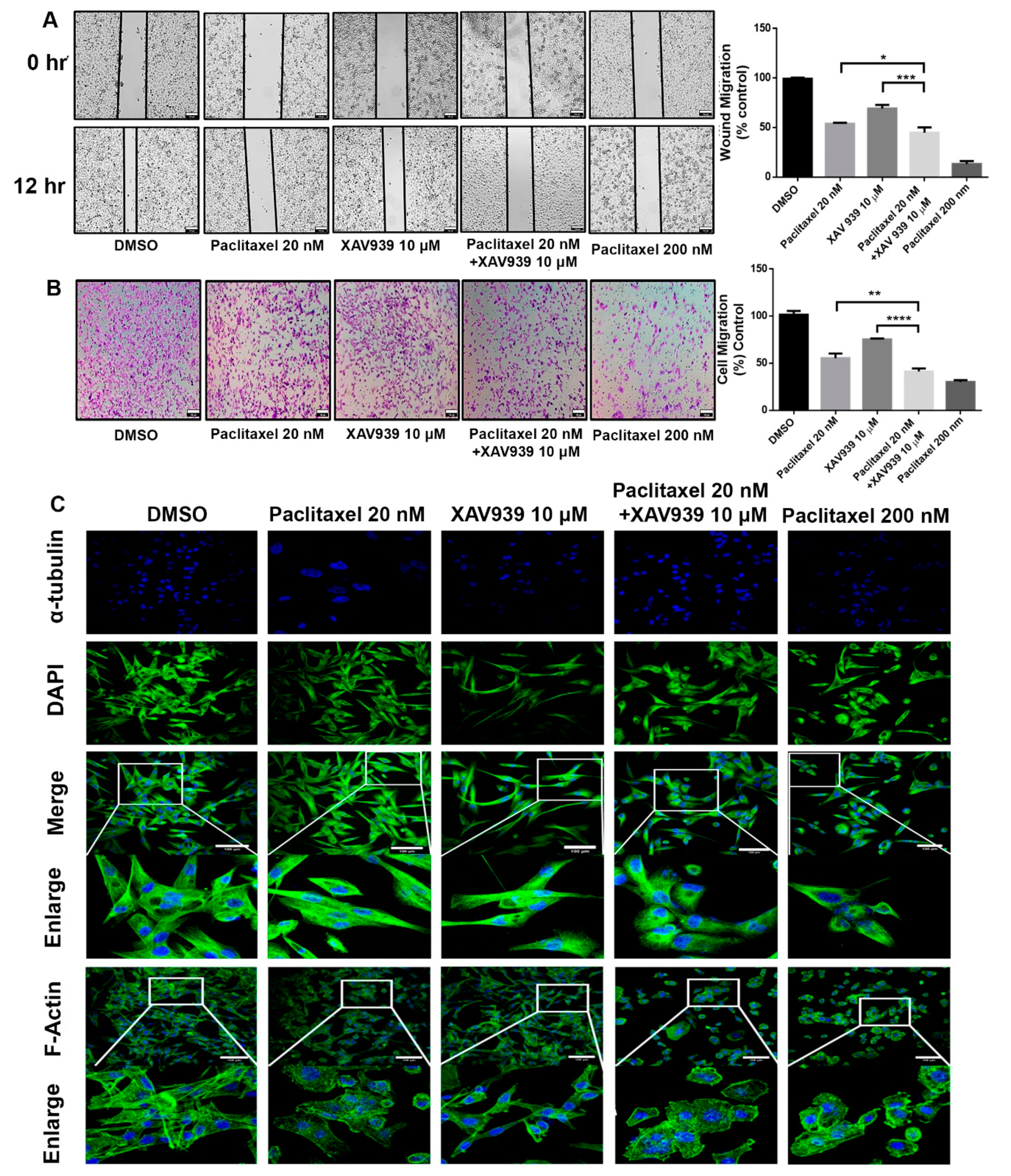
To study the wound migration assay, MDA-MB-231 cells were used. Briefly, MDA-MB-231 cells (5 × 10
5) were seeded in 24-well plates and grown in a monolayer. At 100% confluence, the sterile tip was used to make a straight wound, and then cells were treated with DMSO (3 µL/mL), paclitaxel (20 nM), XAV939 (10µ M), paclitaxel + XAV939 (20 nM + 10 µM), and paclitaxel (200 nM). Wound photos were taken at 0 h and 12 h under a microscope (Olympus). The extent of migration was measured by calculating the area of the remaining wound using Image J software (ImageJ Free ware, NIH, Bethesda, MD, USA,
http//rsb.nih.gov/ij/) and expressed in the form of bar graphs.
2.7. Transwell Assay
Invasive ability of MDA-MB-231 cells was studied using a Transwell cell culture chamber (Corning, NY, USA). Briefly, MDA-MB-231 cells were treated with DMSO (3 µL/mL), paclitaxel (20 nM), XAV939 (10 µM), paclitaxel + XAV939 (20 nM + 10 µM), and paclitaxel (200 nM) for 24 h. MDA-MB-231 cells (2.5 × 104) were harvested and seeded on the upper part (8.0 μm pore polycarbonate membrane) of the Boyden chamber coated with Matrigel (Corning). The Matrigel coating was conducted at a serum-free DMEM: Matrigel ratio (V/V) of 8 to 1 at 37 °C for 2 h. At the lower chamber, a 900 μL complete medium containing 5% FBS was added as chemoattractant. After 24 h incubation, migrated cells on the lower surface of the transwell membrane were fixed with paraformaldehyde and stained with 1% crystal violate. The number of migrated cells was counted from each well and photographed at 20× (Olympus). The experiment was repeated three times.
2.8. Nuclear Staining
To study the nuclear morphology of MDA-MB-231 and MCF-7 cells, Hoechst staining was performed. MBA-MB-231 (1 × 105 cells) and MCF-7 (1 × 105 cells) were seeded separately on the cover slip in a 24-well plate and then each cell line was treated with paclitaxel (20 nM), XAV939 (10 µM), paclitaxel + XAV939 (20 nM + 10 µM), and paclitaxel (200 nM) for 72 h, respectively. The cells were fixed for 20 min with 4% PFA, washed with PBS, and incubated with 0.3% Triton-X 100 for 5 min. The cells were washed with PBS, and Hoechst 33258 (Beyotime, Shanghai, China) was added to cells for 30 min in the dark. The cover slip was mounted with a drop of the antifluorescent quencher (Beyotime, Shanghai, China). The stained nuclei were photographed under the LSM 710 Laser scanning confocal microscope at 40× (Carl Zeiss, Oberkochen, German).
2.9. Immunofluorescence Study
To study the cytoskeleton reorganization in a breast cancer cell line (MDA-MB-231), an immunofluorescence technique was used. MDA-MB-231 cells (1 × 105) were seeded on cover slips in a 24-well plate. Cells were treated with paclitaxel (20 nM), XAV939 (10 µM), paclitaxel + XAV939 (20 nM + 10 µM) and paclitaxel 200 nM, respectively for 48 h. Then, the treated cells were fixed with 2% paraformaldehyde for 15 min at room temperature and washed with PBS twice. Later, the cells were washed with a quenching solution (0.1% glycine in PBS) twice, permeated with 0.1% Triton X-100 for 10 min, and blocked using a blocking solution (10% FBS in PBS) for 1 h at room temperature. Afterwards, the cells were treated overnight with a primary antibody against Tubulin (anti α-Tubulin, 1:100, 2125, CST) followed by a secondary antibody—Alexa Fluor 488 conjugated goat anti-rabbit IgG (1:50, ZF-0511, Zhongshan Goldenbridge Biotechnology, Corp). To study the actin cytoskeleton reorganization, also the cells were stained with FITC conjugated phalloidin (Fushen, Shanghai, China). Also, nuclei were stained with 4,6-diamidino-2 phenylindole DAPI, and the cover slip was mounted with a drop of antifluorescent quencher (Beyotime, Shanghai, China). Immunofluorescence images were captured under a confocal microscope (Nikon Ti-E-A, Tokyo, Japan). Digital images were optimized for image resolution (a final resolution of 300 dpi), brightness, and contrast using Adobe Photoshop 7.0 (Adobe Systems, San Jose, CA, USA). No alteration was made to the image, such as addition or removal of image details.
2.10. Western Blotting Analysis
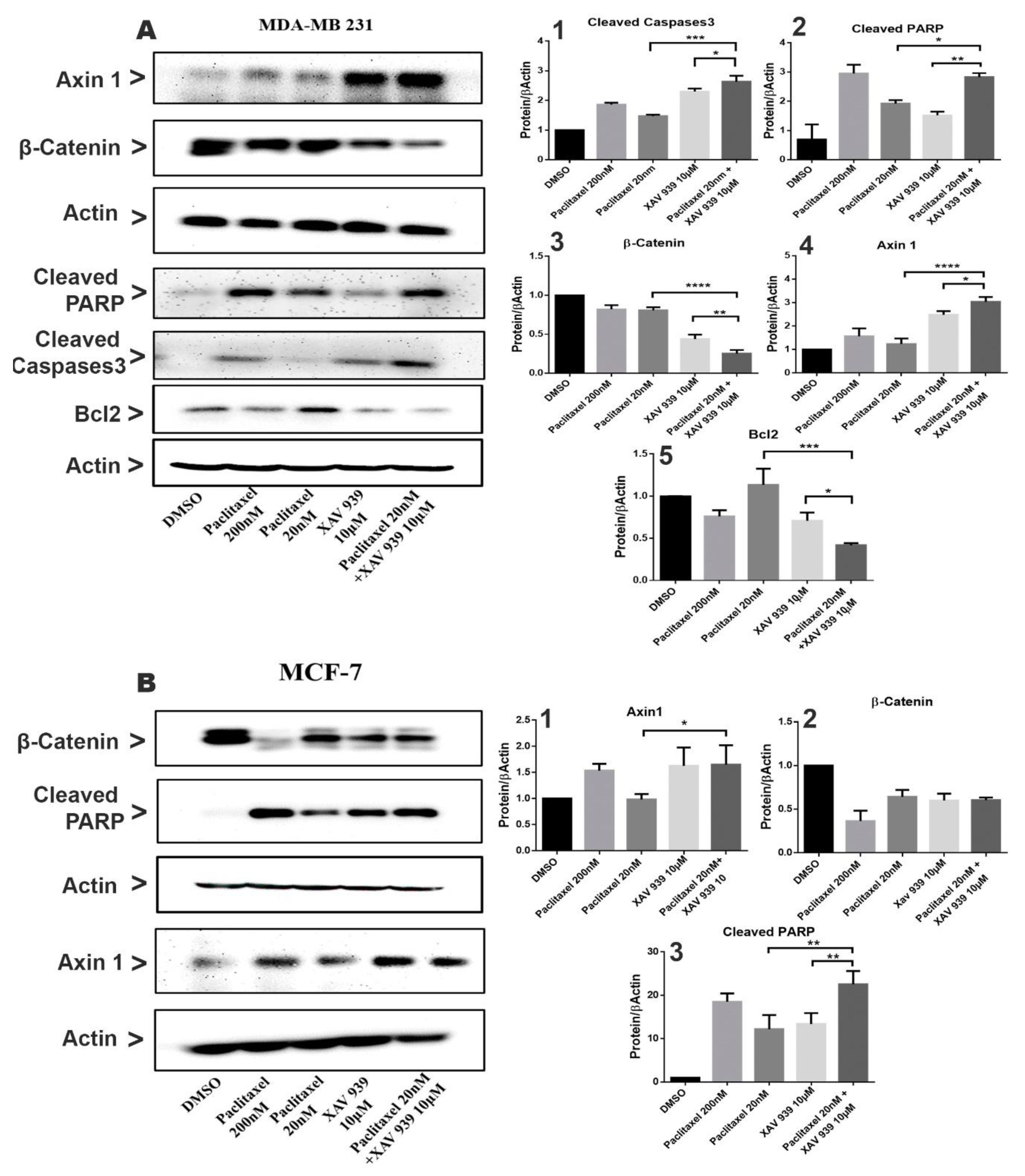
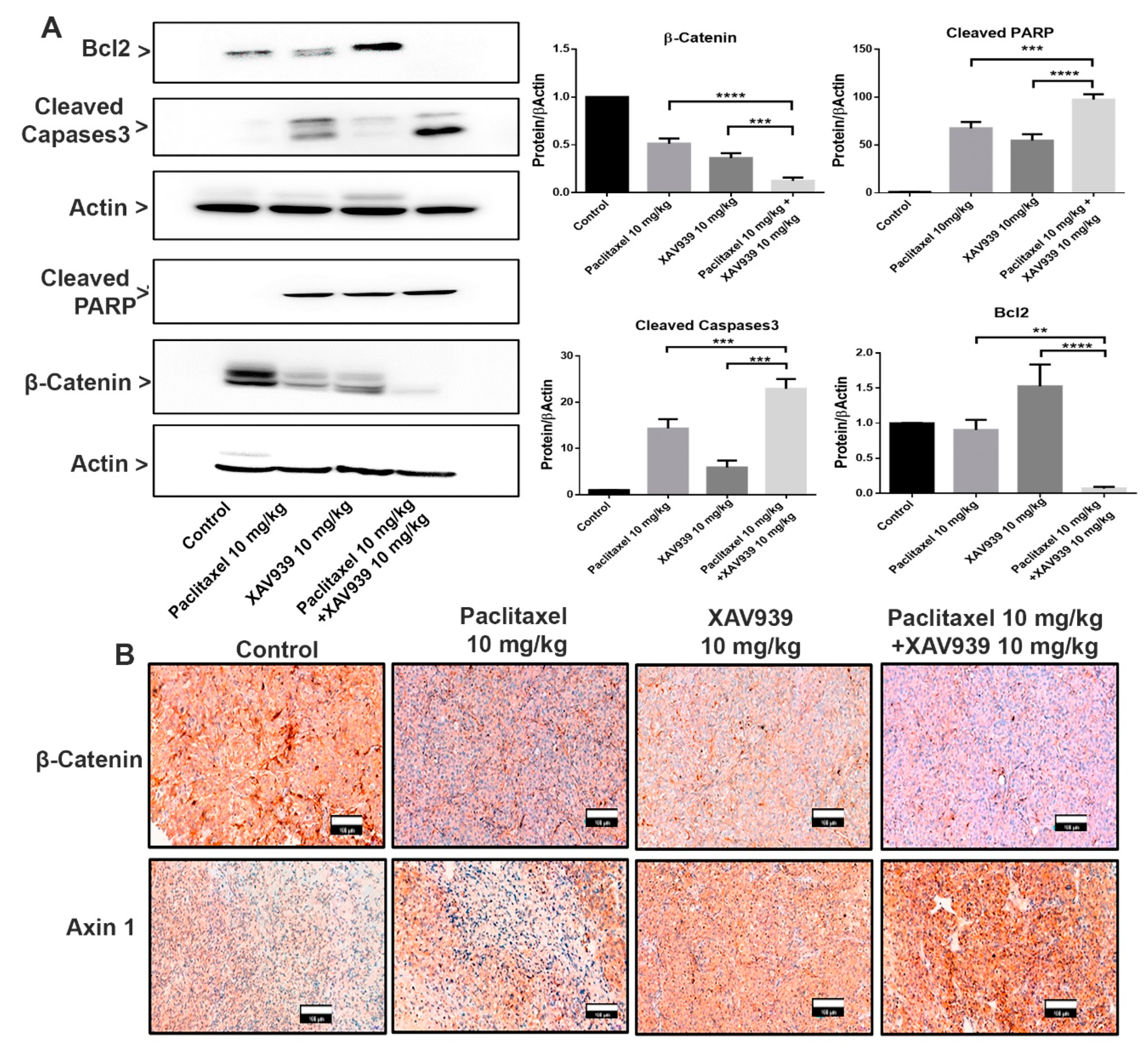
Two breast cancer cell lines, MDA-MB-231 and MCF-7 cells (1 × 106 cells/each well in 6-well plates), were treated with DMSO (3 µL/mL), paclitaxel (20 nM), XAV939 (10 µM), paclitaxel + XAV939 (20 nM + 10 µM), and paclitaxel (200 nM) for 48 h. Then, cells were lysed, and their proteins were isolated by using 1 mL of RIPA (Solarbio, Beijing, China) plus 10 µL of PMSF. The protein concentration of each cell lysate was measured with a BCA Protein Assay Kit (Sangon Biotech, Shanghai, China). Western blot analysis was carried out by separating 40 µg of each cell lysate via sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and by electro-transferring it onto a polyvinylidene difluoride (PVDF) membrane (240 mA, 4 h). The membranes were blocked with 5% nonfat milk, washed with TBST buffer, and then incubated with primary antibodies—β-catenin (D10A8), Axin1 (C7B12), cleaved PARP (Asp214), cleaved caspase-3 (Asp175), and Bcl-2 (124) (15071) (Cell Signaling Technology) at a dilution of 1:1000 in Primary Antibody Dilution Buffer (Beyotime. Shanghai, China) at 4 °C overnight. Next, membranes treated with primary antibodies were washed 3 times with TBST and incubated with an anti-rabbit HRP-linked antibody at a 1:5000 dilutions for 1 h at room temperature. β-Actin was used as an internal reference. The signals were visualized by Immobilon Western Chemiluminescent HRP Substrate (Millipore Corporation, Billerica, MA, USA) with the Amersham Imager 600 imagers (GE Healthcare Life Science, Pittsburgh, PA, USA).
2.11. Pristane-Induced Mammary Tumorigenesis
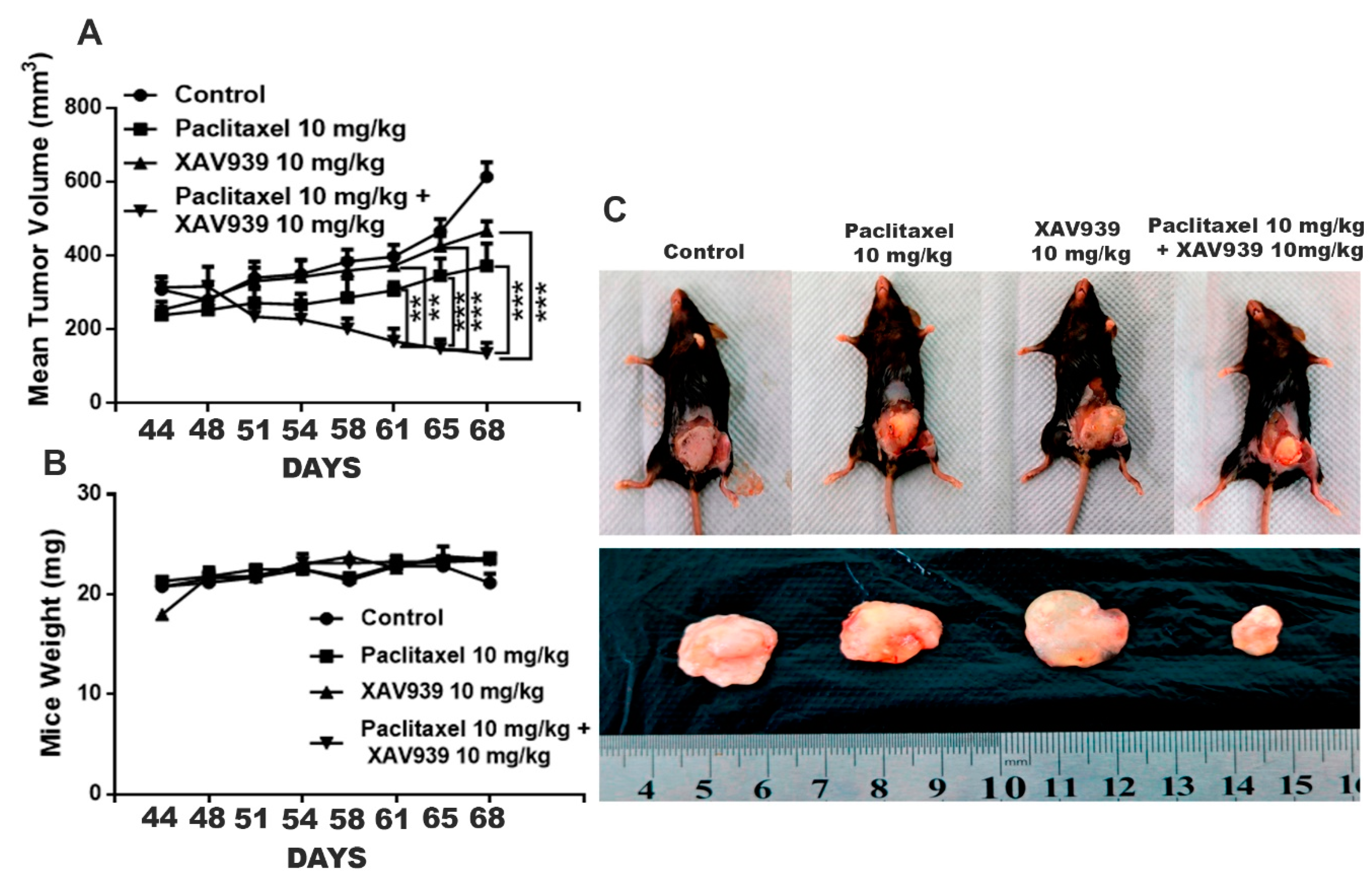
All mice experiments were performed according to the institutional guidelines, following a protocol approved by the South China University of Technology Experimental Animal Center, Guangzhou, China. Pristane (200 µL, Aladdin Industrial, Shanghai, China) was injected in the left inguinal mammary fat pad of 6-week-old C57 female mice twice a week for up to 4 weeks [
21]. About 50% of the pristane-treated mice population were able to induce a tumor near the mammary fat pad. Once the tumor volume became around 200 mm
3–300 mm
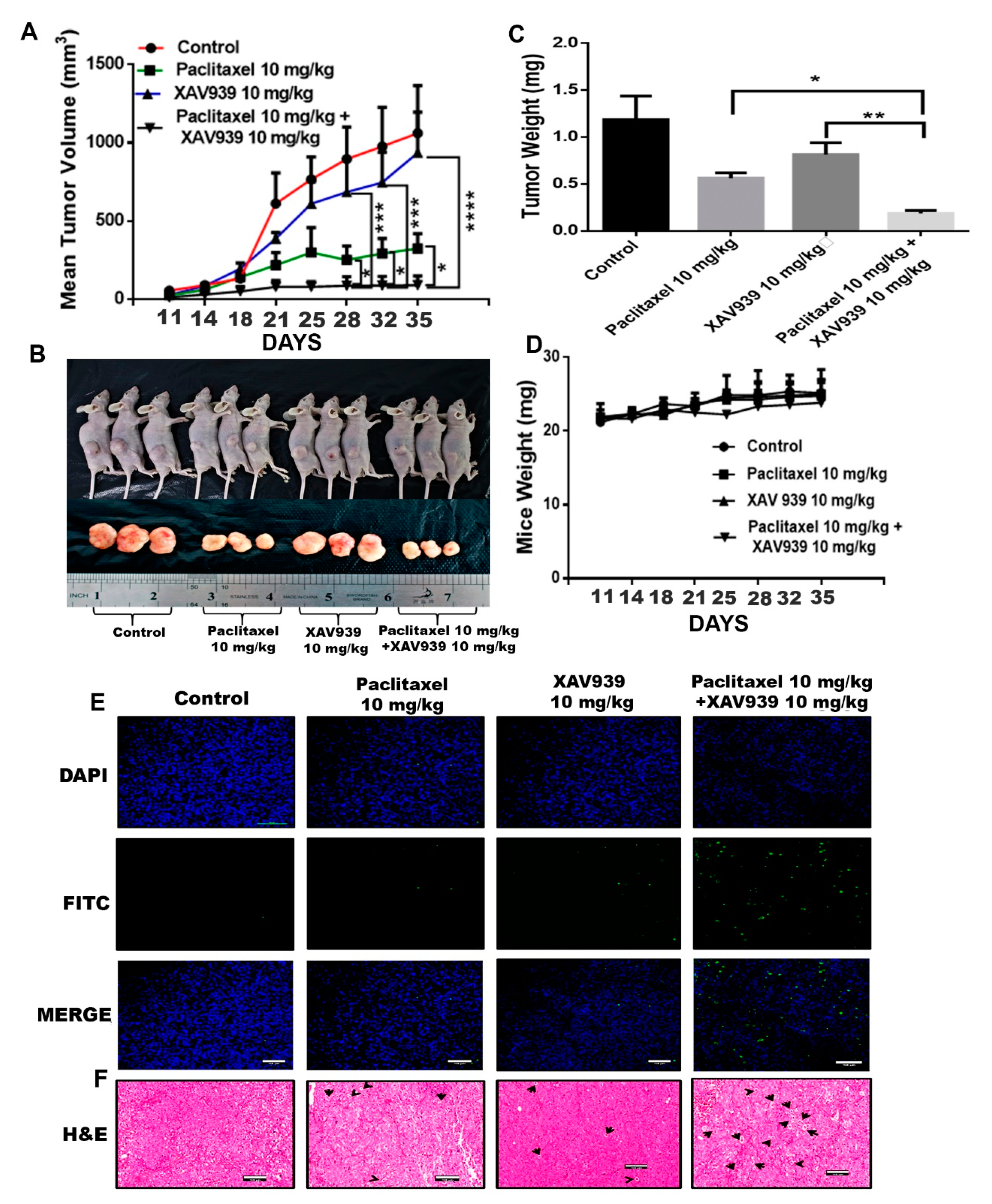
3, mice were segregated into four groups (3 mice/each group), and then mice were treated with paclitaxel (10 mg/kg), XAV939 (10 mg/kg), and a combination of paclitaxel (10 mg/kg) + XAV939 (10 mg/kg) at pristane-induced tumor sites twice a week for 4 weeks. Mice body weight and tumor volume were measured every week using a weighing machine and Vernier caliper. The tumor volume was calculated via using the formula V = (W (2) × L)/2 where V is the volume of the tumor, W is width of the tumor, and L is the length of the tumor. At designated time points, mice were sacrificed and photographed using a camera Canon 750d (Tokyo, Japan)
2.12. Tumor-Xenografted Mice Model
All mice experiments were performed according to the institutional guidelines, following a protocol approved by the South China University of Technology Experimental Animal Center, Guangzhou, China. MDA-MB-231 cells (2.5–5 × 106) were suspended in PBS and mixed with Matrigel at a ratio of 1:1. 150 µL. Suspended cells were injected subcutaneously into 6-week-old female BALB/c nude mice. Once a tumor was formed, mice were randomly segregated into four different groups (three mice/each group). Then, mice were treated with paclitaxel (10 mg/kg), XAV939 (10 mg/kg), and a combination of paclitaxel (10 mg/kg) + XAV939 (10 mg/kg) via intraperitoneal injection twice a week for 4 weeks. Mice body weight and tumor volume were measured every week with a weighing machine and Vernier caliper. The tumor volume was calculated via using the formula V = (W (2) × L)/2 where V is the volume of the tumor, W is the width of the tumor, and L is the length of the tumor. At designated time points, mice were sacrificed, and the tumor tissue in each mouse was weighed and photographed using a camera (Canon750d). For Western blot and PCR analyses, tumor tissues were snap-frozen. In addition, some of the tumor tissues were stored in 4% PFA for the immunohistochemistry and H&E staining.
2.13. Quantitative Real-Time RT-PCR
Total tumor RNA was isolated from snap-frozen tumor samples using TRIzol reagent (Sangon Biotech, Shanghai, China) according to the manufacture’s instruction. Single-stranded RNA was reverse transcribed into complementary DNA (cDNA) using ReverTra Ace
® qPCR RT Master Mix (TOYOBO LIFE SCIENCE, Shanghai, China) in T100™ Thermal Cycler (BIO RAD, Hercules, CA, USA). The sample mixture was prepared with 1.5 μL of cDNA in 10 μL of ChamQ SYBR qPCR Master Mix (Vazyme Biotech, Nanjing, China), primers, and ddH
2O for a final volume of 20 μL. The primers for specific genes, which were purchased from Sangon Biotech (Shanghai, China), are mentioned below (
Table 1). The LightCycler
® 96 Real-Time PCR System (Roche, Switzerland) was used to perform qPCR under the standard thermal conditions: 95 °C for 30 s, 40 cycles of 95 °C for 10 s, and 60 °C for 30 s, followed by 95 °C for 10 s, 65 °C for 60 s, and 97 °C for 1 s. GAPDH was used as an endogenous housekeeping gene. The threshold cycle (CT) was calculated in accordance with the 2
−ΔΔCt method relative to the expression of GAPDH. Three independent experiments were performed for each reaction in triplicate.
2.14. Haematoxylin and Eosin Stain (H&E Stain)
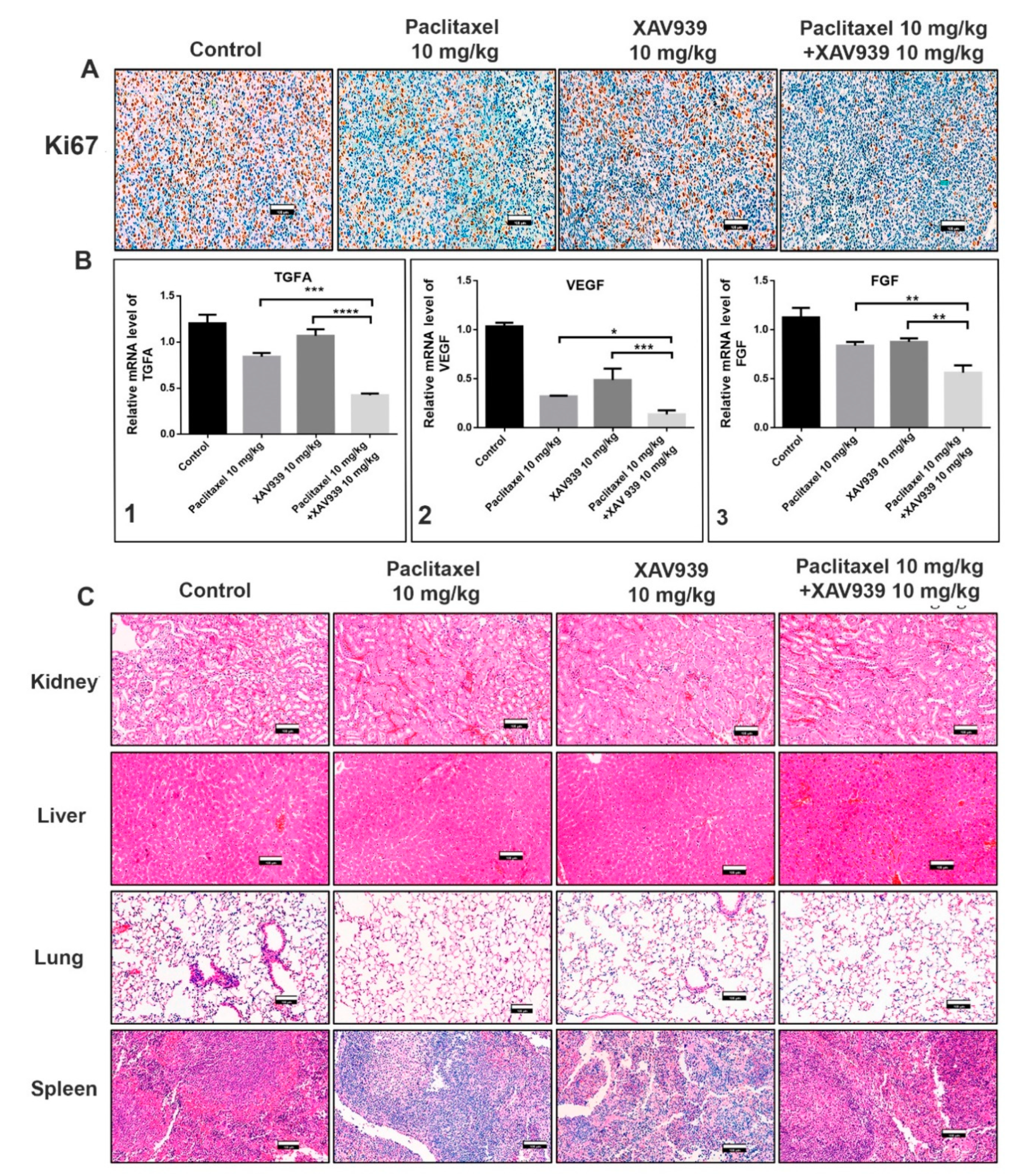
Hematoxylin eosin staining was performed on paraffin-embedded tumor, kidney, liver, lung, and spleen tissues of MDA-MB-231-xenografted mice. Sections with a thickness of 4 µm were prepared, deparaffinized in xylene (two times), and rehydrated in a graded series of alcohol (two times 100% alcohol and 75% alcohol) and distilled water. Later, sections were stained with a hematoxylin solution, rinsed in water, passed through a 70% ethanol solution containing 1% HCl, and rinsed again with tap water. Sections were stained with eosin for 5 min and rinsed with absolute alcohol and xylene for 5 min. The images were taken by Pannoramic 250 Flash III (3D HISTECH, The Digital Pathology Company, Budapest, Hungary).
2.15. TUNEL Assay
TUNEL assay was performed to see nuclear DNA fragmentation in apoptotic cells using in situ Cell Death Detection Kit (Roche), according to the manufacturer’s instruction. In brief, paraffin sections of MDA-MB-231-xenografted mice tumor samples were deparaffinized in xylene and rehydrated in a series of graded alcohol, and permeabilized with proteinase K for 25 min 37 °C. The tissue sections were then incubated with TUNEL reaction buffer in a 37 °C humidified chamber for 2 h, and the nucleus was counterstained with DAPI for 1 min at room temperature. Section slides were covered with a drop of the antifade mounting medium. Stained apoptotic cells were visualized by confocal microscopy (Nikon Ti-E-A1).
2.16. Immunohistochemistry
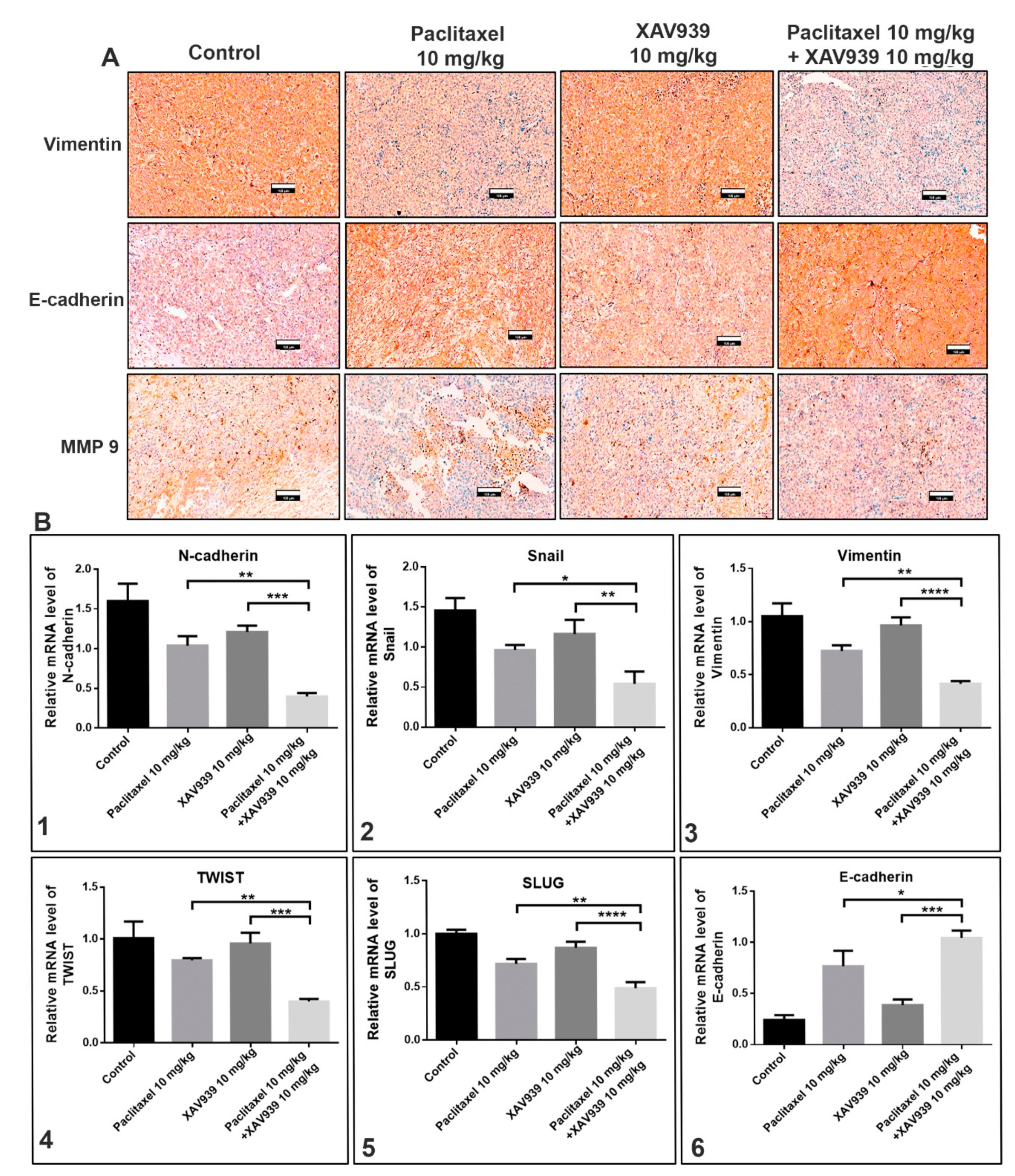
Immunohistochemistry (IHC) was performed on 4 µm paraffin-embedded tumor sections of MDA-MB-231-xenografted mice. Sections were deparaffinized with xylene and rehydrated in pure ethanol and gradient ethanol (85% and 75%) for 5 min per step. Then, section slides were placed in EDTS (pH9.0) and pressure-cooked with water. Slides were blocked using 3% H2O2 and washed with PBS. Primary antibodies against MMP9 (1:800 Serveicebio), E-cad (1:500 Serveicebio), vimentin (1:800 Serveicebio), Axin1 (1:50 CST), β-catenin (1:400 Serveicebio), and Ki-67(1:500 Serveicebio) were incubated for overnight at 4 °C. Next morning, slides were washed with PBS. Secondary antibody goat anti-rabbit IgG H&L (HRP) labelled with HRP was added and incubated for 50 min. A freshly prepared DAB chromogenic reagent was added to the dried slides as per the manufacture’s instruction, and the slides were counter-stained with hematoxylin. The slides were then rinsed with xylene for 5 min and mounted with a resin mounting medium. The images were taken by Pannoramic 250 Flash III (3D HISTECH, The Digital Pathology Company, Budapest, Hungary).
2.17. Statistical Analysis
All the experimental data are expressed as mean ± SD (standard deviation, n = 3). Statistical analysis was performed by one-way ANOVA by using GraphPad Prism 6.01 software. In all cases, p < 0.05 was considered statistically significant.
4. Discussion
The Wnt signaling pathway is a crucial process in embryonic development and tissue homeostasis where cell surface receptors are initiated by a Wnt stimulus leading to β-catenin activation and transcription of Wnt-responsive genes. Based on the relation with β-catenin, Wnt signaling is classified into canonical (β-catenin-dependent) and noncanonical (β-catenin-independent) pathways. Dysregulation of Wnt signaling is a hallmark in many types of cancer, and breast cancer is no exception [
7]. Studies have shown that canonical and noncanonical pathways are highly expressed in TNBC tumorigenesis and metastasis [
11,
12]. Moreover, in TNBC patients, the Wnt/β-catenin pathway is highly dysregulated, which leads to lung and brain secondary metastasis [
22]. Over the past decades, inhibition of Wnt signaling was a universal strategy for treating cancer [
23]. Recently, XAV939, a small molecule inhibitor of Wnt signaling has been identified [
24]. XAV939 was found to selectively promote β-catenin degradation by stabilizing Axin1 and to suppress the Wnt signaling pathway. However, it has been reported that XAV939 alone could not efficiently inhibit metastasis and cell growth in the breast cancer cell line [
24]. Therefore, a new strategy should be applied for treating TNBC.
In this study, we tested a well-known cancer drug (paclitaxel) at a low toxic dose in combination with XAV939 for treating breast cancer. MTT results indicate that the combination treatment with paclitaxel and XAV939 is more effective in inhibiting cell proliferation of TNBC cell lines (MDA-MB-231, MDA-MB-468, and BT549) and ER+ve cell lines (MCF-7 and T-47D) compared to the single treatment with either paclitaxel or XAV939. The therapeutic effect of the combination regimen with a low dose of paclitaxel (20 nM) plus XAV939 (10 µM) was comparable to that of paclitaxel with a high dose (200 nM), confirming that the combination treatment is synergistically effective in inhibiting the growth of various breast cancer cells. The combination treatment with paclitaxel and XAV939 suppressed the proliferation of MDA-MB-231 more significantly than that of MCF-7. MDA-MB-231 cells are triple negative and highly aggressive, and their Wnt signaling is aberrantly regulated. It is important to find a strategy to tackle the growth and metastasis of MDA-MB-231 [
25,
26]. Cell cycle arrest is a major requirement of any cancer drug; most of the cancer drugs available on the market can induce cell cycle arrest in various phases of the cell cycle in cancer cells. Paclitaxel is known to bind to microtubules and to prevent their reorganization during cell division, subsequently resulting in cell cycle arrest. Also, the combination treatment with paclitaxel and XAV939 was found to interfere with the microfilament and microtubule structure in cancer cell lines. Moreover, in the treatment with paclitaxel and XAV939, cell apoptosis in MDA-MB-231 was induced at 48 h, and their DNA fragmentation was observed at 72 h, which suggests that the prolonged treatment with paclitaxel and XAV939 could trigger the apoptosis pathway like the treatment with a high dose of paclitaxel. Also, the combination treatment inhibited the canonical pathway in Wnt signaling by suppressing β-catenin expression and induced apoptosis in TNBC cells, in ER+ve cells, and in the tumor sample of the tumor-xenografted mice by suppressing Bcl-2 expression and by activating caspases3 that mediated PARP cleavage, ultimately leading to DNA condensation and fragmentation. In previous reports, during apoptosis, the expression of Bcl-2 was downregulated to facilitate the mitochondrial apoptotic pathway in which cytochrome C was released, and caspase 9 and caspase3 were activated, ultimately resulting in PARP cleavage [
27,
28]. Upregulation of PARP cleavage triggered an apoptotic signal and translocated the mitochondrial apoptosis-inducing factor (AIF) into the nucleus and provoked chromatin condensation and fragmentation [
29]. The molecular mechanism by which the combination treatment can inhibit Wnt signaling and mediate apoptosis in MDA-MB-231 cells needs to be further studied. Nevertheless, the current study provides a basic platform for the further evaluation of the combination treatment against breast cancer.
EMT (epithelial to mesenchymal transition) is well associated with TNBC and plays a critical role in metastasis and progression of cancer [
30]. In a recent study in TNBC patients, EMT markers have been established (vimentin, smooth muscle actin, osteonectin, and N-cadherin; loss of E-cadherin), and EMT inducers (ZEB1 and CD146) play an important role in EMT [
31]. EMT is a complex process in which tumor cells secrete matrix metalloproteinases (MMPs). MMPs help in the degradation of the extracellular matrix (ECM) and allow tumors to lose their epithelial characteristics and to acquire mesenchymal phenotypes that are highly regulated by transcriptional factors such as Zeb1, Slug, Snail, TGF-β, and Twist [
32,
33]. Some of the transcriptional factors have been reported to play a role in angiogenesis [
34]. Some studies have reported that the Wnt pathway plays an important role in promoting EMT and angiogenesis in cancer. Overexpression of Wnt3 ligand activated the Wnt/β-catenin pathway, whereas epidermal growth factor (EGF) secretion activated PI3K/Akt pathways, which accounted for β-catenin accumulation and mediated EMT and angiogenesis [
35,
36,
37]. Since β-catenin is a major protein in the Wnt signaling pathway, targeting Wnt signaling can be a good strategy to control EMT and tumorigenesis. A previous report demonstrated that a small molecule inhibitor of Wnt signaling could inhibit cell migration and invasion in vitro [
27]. Our results further disclose that a combination of a low dose of paclitaxel and a small molecule inhibitor (XAV939) can significantly suppress cancer cell migration and invasion in vitro and in vivo. In the TNBC-xenografted mice model, the combination treatment with paclitaxel and XAV939 efficiently downregulated the expression of β-catenin and stabilized Axin1, indicating the effective inhibition of the canonical pathway of Wnt signaling. Based upon the previous results, we speculate that our combination treatment with paclitaxel and XAV939 might inhibit the EMT pathway and angiogenesis in vivo. In fact, the combination treatment suppressed the expression of EMT regulating transcription factors (SNAIL, SLUG and Twist) at the mRNA level and inhibited the expression of pro-EMT markers (Vimentin, N-cadherin, and MMP 9) at the protein and mRNA levels. Besides, our combination treatment increased the E-cadherin level, which potentially corresponds to the inhibition of EMT. This inhibition was further correlated with the downregulation of proangiogenic molecules like Ki-67 in tumor samples, whereas the expression levels of VEGF, TGFA, and FGF were downregulated at mRNA level. Taken together, all these results suggest that the combination treatment with paclitaxel and XAV939 shows good potential in inhibiting various hallmarks of breast cancer.
In recent years, an external carcinogen is believed to be one of the major causes for cancer development. One study reported that an external carcinogen played a major role in altering the gene expression and activated oncogene genes, eventually inducing cancer progression [
38]. Also, we studied the therapeutic effect of the combination treatment (paclitaxel and XAV939) on a carcinogen-induced breast tumor. The direct administration of the combination regimen significantly attenuated the growth and development of the pristane-induced breast cancer.
Taken together, the single treatment with either a low dose of paclitaxel or XAV939 inhibited the Wnt pathway but could not significantly suppress TNBC breast cancer. Importantly, our strategy of combining a low dose of paclitaxel and XAV939 exhibited the strong antitumor activity in TNBC and inhibited EMT and angiogenesis by suppressing the canonical pathway of Wnt signaling in the MDA-MB-231-xenografted mice model. For the first time, we demonstrated that a combination of Tankyrase inhibitor i.e., XAV939 and a low dose of paclitaxel treatment could be a potential approach for treating TNBC cancer and carcinogen-induced breast cancer with minimum side effects.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}