Evidence for Overlapping and Distinct Biological Activities and Transcriptional Targets Triggered by Fibroblast Growth Factor Receptor 2b Signaling between Mid- and Early Pseudoglandular Stages of Mouse Lung Development

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement and Husbandry
2.2. Mouse Model to Inhibit Fgfr2b Ligands
2.3. Embryonic Lung Dissection and Imaging
2.4. DNA Isolation and PCR
2.5. Immunofluorescence
2.6. RNA Lsolation and RT-qPCR
2.7. Microarray
2.8. Microarray Expression Analysis
2.9. FACS
2.10. Gene Expression Patterns
2.11. Proliferation and Apoptosis
2.12. Relative Gene Expression from qPCR Data
2.13. Statistics
3. Results
3.1. Identification of the Early Fgfr2b Transcriptomic Signature at E14.5 Supports a Primary Role for Fgf Signaling in Proliferation
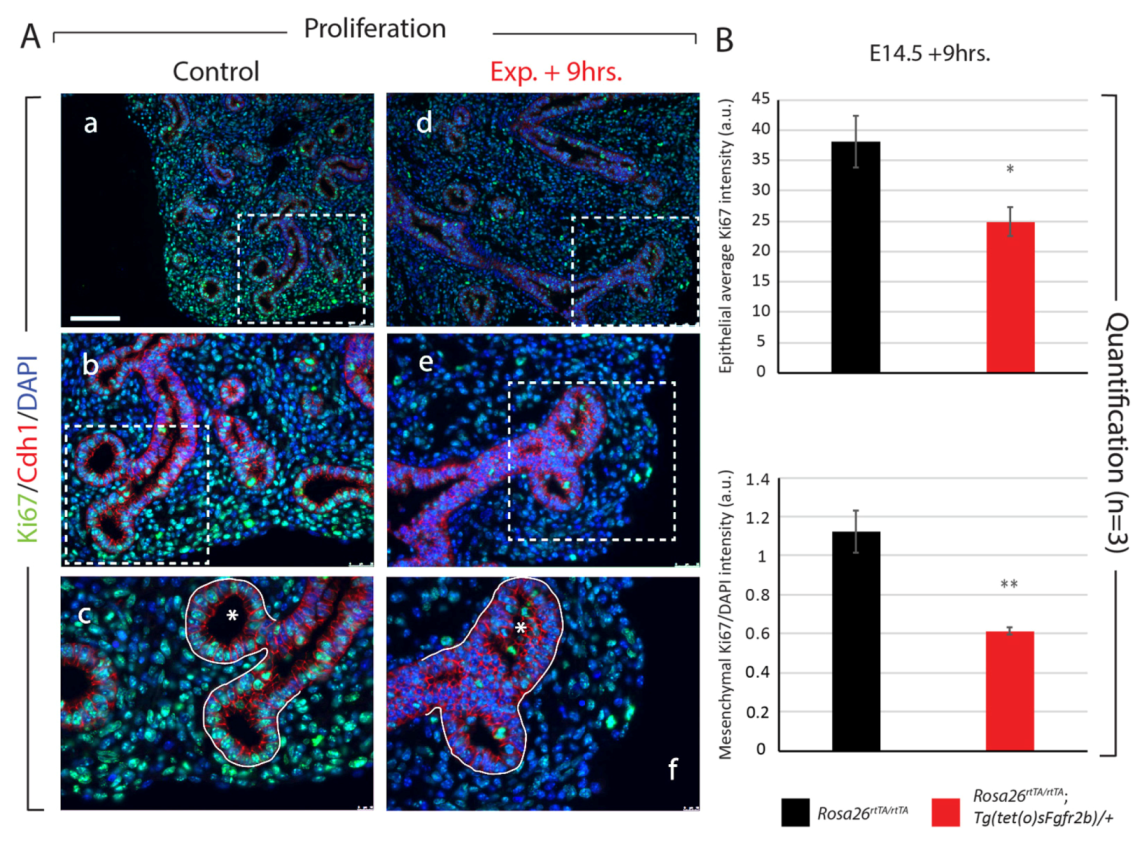
3.2. Inhibition of Fgfr2b Signaling for 24 h Drastically Decreases Proliferation and Impacts AT2 Differentiation
3.3. Continuous Inhibition of Fgfr2b Signaling from E14.5 to E18.5 Leads to Impaired AT1 and AT2 Formation
3.4. Transient Inhibition of Fgfr2b at E14.5 Leads to Irreversible Developmental Damages at E18.5
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Herriges, M.; Morrisey, E.E. Lung development: Orchestrating the generation and regeneration of a complex organ. Dev. 2014, 141, 502–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburton, D.; El-Hashash, A.; Carraro, G.; Tiozzo, C.; Sala, F.; Rogers, O.; De Langhe, S.; Kemp, P.J.; Riccardi, D.; Torday, J.; et al. Lung Organogenesis. In Current Topics in Developmental Biology; 2010; Volume 90, pp. 73–158. [Google Scholar]
- Metzger, R.J.; Klein, O.D.; Martin, G.R.; Krasnow, M.A. The branching programme of mouse lung development. Nature 2008, 453, 745–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schittny, J.C. Development of the lung. Cell Tissue Res. 2017, 367, 427–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, W.V.; Lü, J. Regulation of early lung morphogenesis: Questions, facts and controversies. Development 2006, 133, 1611–1624. [Google Scholar] [CrossRef] [Green Version]
- Morrisey, E.E.; Hogan, B.L.M. Preparing for the First Breath: Genetic and Cellular Mechanisms in Lung Development. Dev. Cell 2010, 18, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.M.; El Agha, E.; Tiozzo, C.; Minoo, P.; Bellusci, S. A breath of fresh air on the mesenchyme: Impact of impaired mesenchymal development on the pathogenesis of bronchopulmonary dysplasia. Front. Med. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Sekine, K.; Ohuchi, H.; Fujiwara, M.; Yamasaki, M.; Yoshizawa, T.; Sato, T.; Yagishita, N.; Matsui, D.; Koga, Y.; Itoh, N.; et al. Fgf10 is essential for limb and lung formation. Nat. Genet. 1999, 21, 138–141. [Google Scholar] [CrossRef]
- Parsa, S. Role of FGFR2b signaling pathway in the development of ectodermal derivatives. ProQuest Diss. Theses. 2010. [Google Scholar]
- Mailleux, A.A.; Spencer-Dene, B.; Dillon, C.; Ndiaye, D.; Savona-Baron, C.; Itoh, N.; Kato, S.; Dickson, C.; Thiery, J.P.; Bellusci, S. Role of FGF10/FGFR2b signaling during mammary gland development in the mouse embryo. Development 2002, 129, 53–60. [Google Scholar]
- Bellusci, S.; Grindley, J.; Emoto, H.; Itoh, N.; Hogan, B.L.M. Fibroblast Growth Factor 10 (FGF10) and branching morphogenesis in the embryonic mouse lung. Development 1997, 124, 4867–4878. [Google Scholar]
- Rawlins, E.L.; Clark, C.P.; Xue, Y.; Hogan, B.L.M. The Id2+ distal tip lung epithelium contains individual multipotent embryonic progenitor cells. Development 2009, 136, 3741–3745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.R.; Dilai, S.; Lingampally, A.; Chao, C.M.; Danopoulos, S.; Carraro, G.; Mukhametshina, R.; Wilhelm, J.; Baumgart-Vogt, E.; Al Alam, D.; et al. A comprehensive analysis of fibroblast growth factor receptor 2b signaling on epithelial tip progenitor cells during early mouse lung branching morphogenesis. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.K.; Mailleux, A.A.; Gupte, V.V.; Mata, F.; Sala, F.G.; Veltmaat, J.M.; Del Moral, P.M.; De Langhe, S.; Parsa, S.; Kelly, L.K.; et al. Fgf10 dosage is critical for the amplification of epithelial cell progenitors and for the formation of multiple mesenchymal lineages during lung development. Dev. Biol. 2007, 307, 237–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.M.; Yahya, F.; Moiseenko, A.; Tiozzo, C.; Shrestha, A.; Ahmadvand, N.; El Agha, E.; Quantius, J.; Dilai, S.; Kheirollahi, V.; et al. Fgf10 deficiency is causative for lethality in a mouse model of bronchopulmonary dysplasia. J. Pathol. 2017, 241, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hokuto, I.; Perl, A.K.T.; Whitsett, J.A. Prenatal, but not postnatal, inhibition of fibroblast growth factor receptor signaling causes emphysema. J. Biol. Chem. 2003, 278, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danopoulos, S.; Parsa, S.; Al Alam, D.; Tabatabai, R.; Baptista, S.; Tiozzo, C.; Carraro, G.; Wheeler, M.; Barreto, G.; Braun, T.; et al. Transient Inhibition of FGFR2b-Ligands Signaling Leads to Irreversible Loss of Cellular β-Catenin Organization and Signaling in AER during Mouse Limb Development. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.; Bellusci, S. Imaging and Analysis of Mouse Embryonic Whole Lung, Isolated Tissue, and Lineage-Labelled Cell Culture. Methods Mol. Biol. 2019, 1940, 109–127. [Google Scholar]
- R Core Team (Ed.) A Language and Environment for Statistical Computing, version 3.5.3; European Environment Agency: Vienna, Austria, 2019. [Google Scholar]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015. [Google Scholar] [CrossRef]
- Jones, M.R.; Lingampally, A.; Dilai, S.; Shrestha, A.; Stripp, B.; Helmbacher, F.; Chen, C.; Chao, C.M.; Bellusci, S. Characterization of Tg(Etv4-GFP)and Etv5RFP reporter lines in the context of fibroblast growth factor 10 signaling during mouse embryonic lung development. Front. Genet. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.H.; Siah, K.W.; Lo, A.W. Estimation of clinical trial success rates and related parameters. Biostatistics 2019, 20, 273–286. [Google Scholar] [CrossRef]
- Danopoulos, S.; Thornton, M.E.; Grubbs, B.H.; Frey, M.R.; Warburton, D.; Bellusci, S.; Al Alam, D. Discordant roles for FGF ligands in lung branching morphogenesis between human and mouse. J. Pathol. 2019, 247, 254–265. [Google Scholar] [CrossRef]
- Danopoulos, S.; Shiosaki, J.; Al Alam, D. FGF signaling in lung development and disease: Human versus mouse. Front. Genet. 2019, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Treutlein, B.; Brownfield, D.G.; Wu, A.R.; Neff, N.F.; Mantalas, G.L.; Espinoza, F.H.; Desai, T.J.; Krasnow, M.A.; Quake, S.R. Reconstructing lineage hierarchies of the distal lung epithelium using single-cell RNA-seq. Nature 2014, 509, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, D.B.; Penkala, I.J.; Zepp, J.A.; Sivakumar, A.; Linares-Saldana, R.; Zacharias, W.J.; Stolz, K.G.; Pankin, J.; Lu, M.Q.; Wang, Q.; et al. Early lineage specification defines alveolar epithelial ontogeny in the murine lung. Proc. Natl. Acad. Sci. USA 2019, 116, 4362–4371. [Google Scholar] [CrossRef] [PubMed] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, M.R.; Lingampally, A.; Wu, J.; Sedighi, J.; Ahmadvand, N.; Wilhelm, J.; Vazquez-Armendariz, A.I.; Herold, S.; Chen, C.; Zhang, J.-S.; et al. Evidence for Overlapping and Distinct Biological Activities and Transcriptional Targets Triggered by Fibroblast Growth Factor Receptor 2b Signaling between Mid- and Early Pseudoglandular Stages of Mouse Lung Development. Cells 2020, 9, 1274. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9051274
Jones MR, Lingampally A, Wu J, Sedighi J, Ahmadvand N, Wilhelm J, Vazquez-Armendariz AI, Herold S, Chen C, Zhang J-S, et al. Evidence for Overlapping and Distinct Biological Activities and Transcriptional Targets Triggered by Fibroblast Growth Factor Receptor 2b Signaling between Mid- and Early Pseudoglandular Stages of Mouse Lung Development. Cells. 2020; 9(5):1274. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9051274
Chicago/Turabian StyleJones, Matthew R., Arun Lingampally, Jin Wu, Jamschid Sedighi, Negah Ahmadvand, Jochen Wilhelm, Ana Ivonne Vazquez-Armendariz, Susanne Herold, Chengshui Chen, Jin-San Zhang, and et al. 2020. "Evidence for Overlapping and Distinct Biological Activities and Transcriptional Targets Triggered by Fibroblast Growth Factor Receptor 2b Signaling between Mid- and Early Pseudoglandular Stages of Mouse Lung Development" Cells 9, no. 5: 1274. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9051274