Impacts of the Degraded Alpine Swamp Meadow on Tensile Strength of Riverbank: A Case Study of the Upper Yellow River

 ,
,
Abstract
:1. Introduction
2. Experiences at Field Scale and Procedure for Determining the Cantilanver Bank Collapse
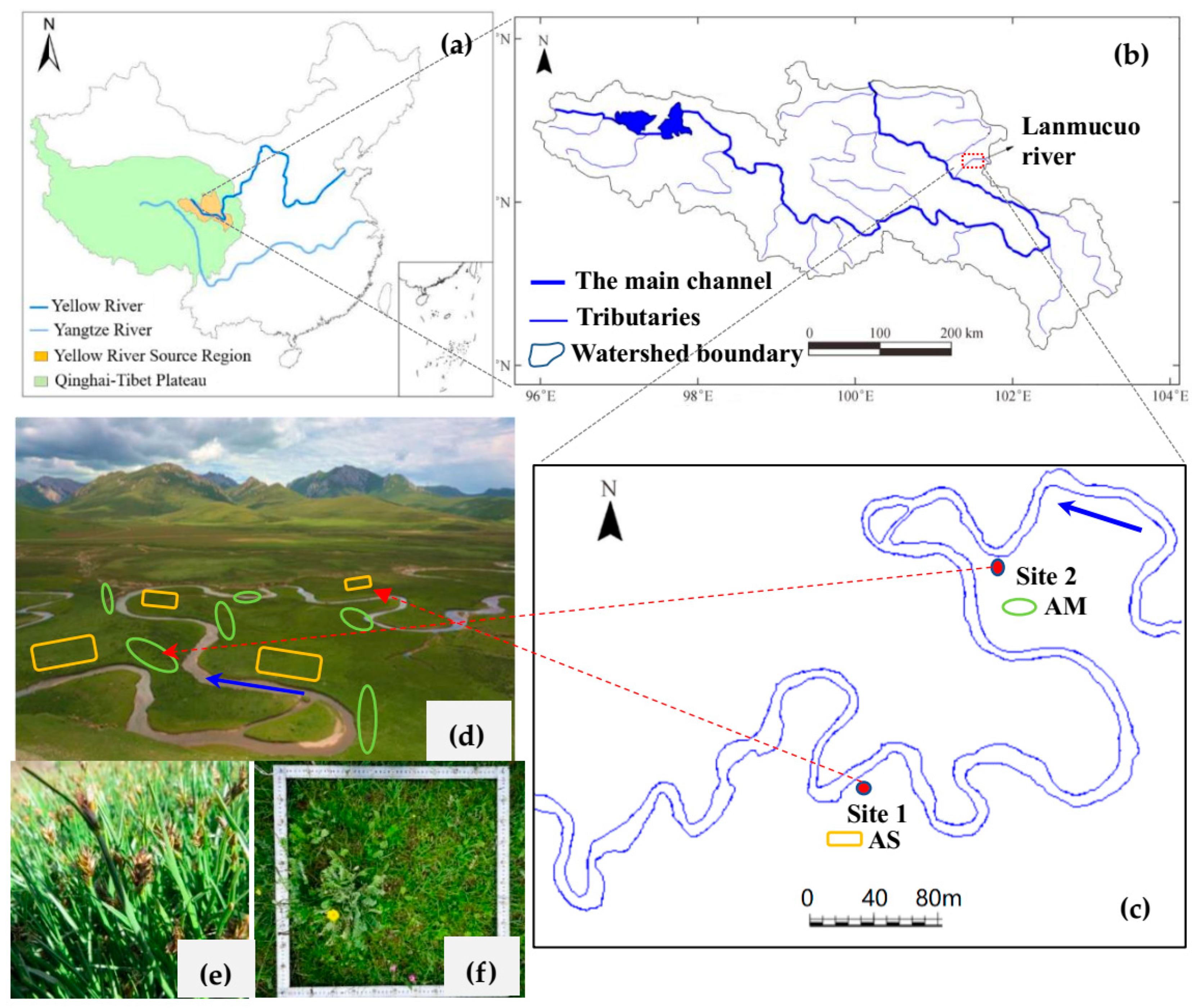
2.1. Study Sites and Degradation of Alpine Swamp Meadow
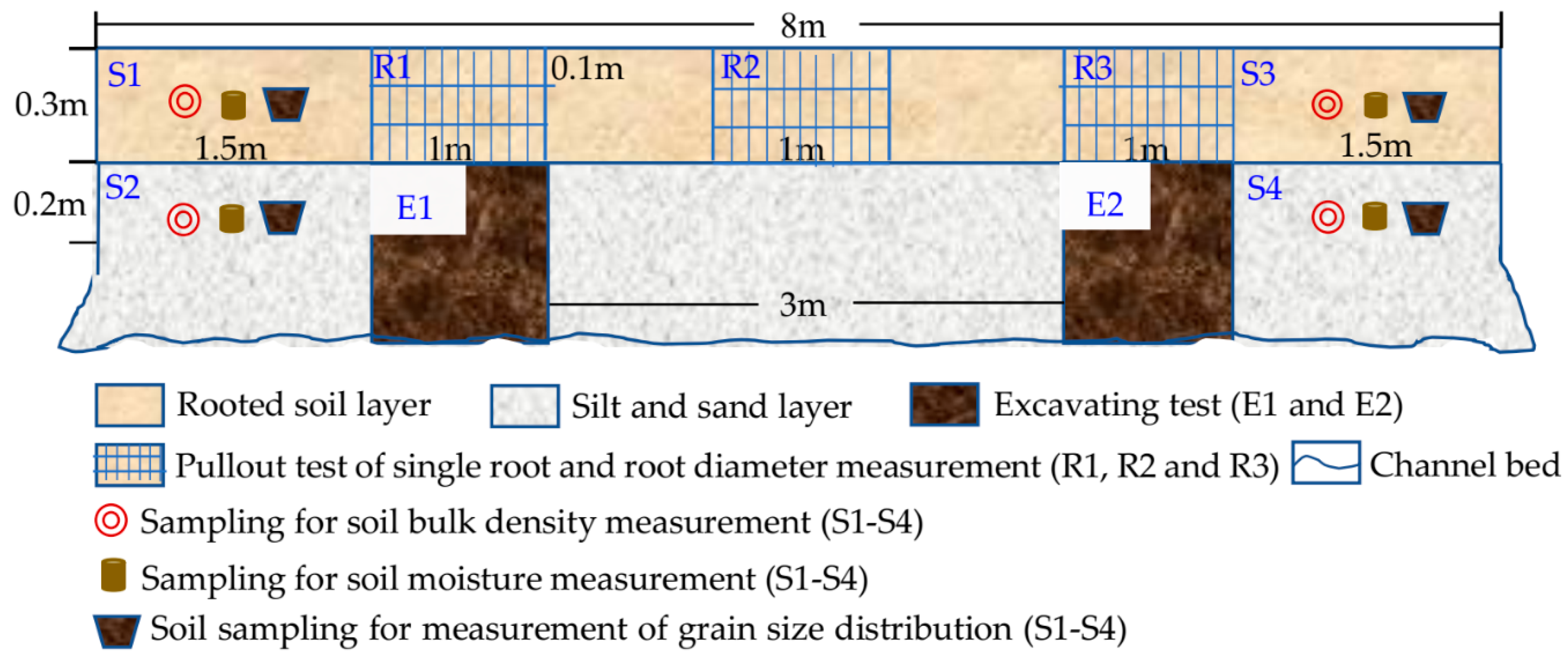
2.2. Field Experiments and Measurements
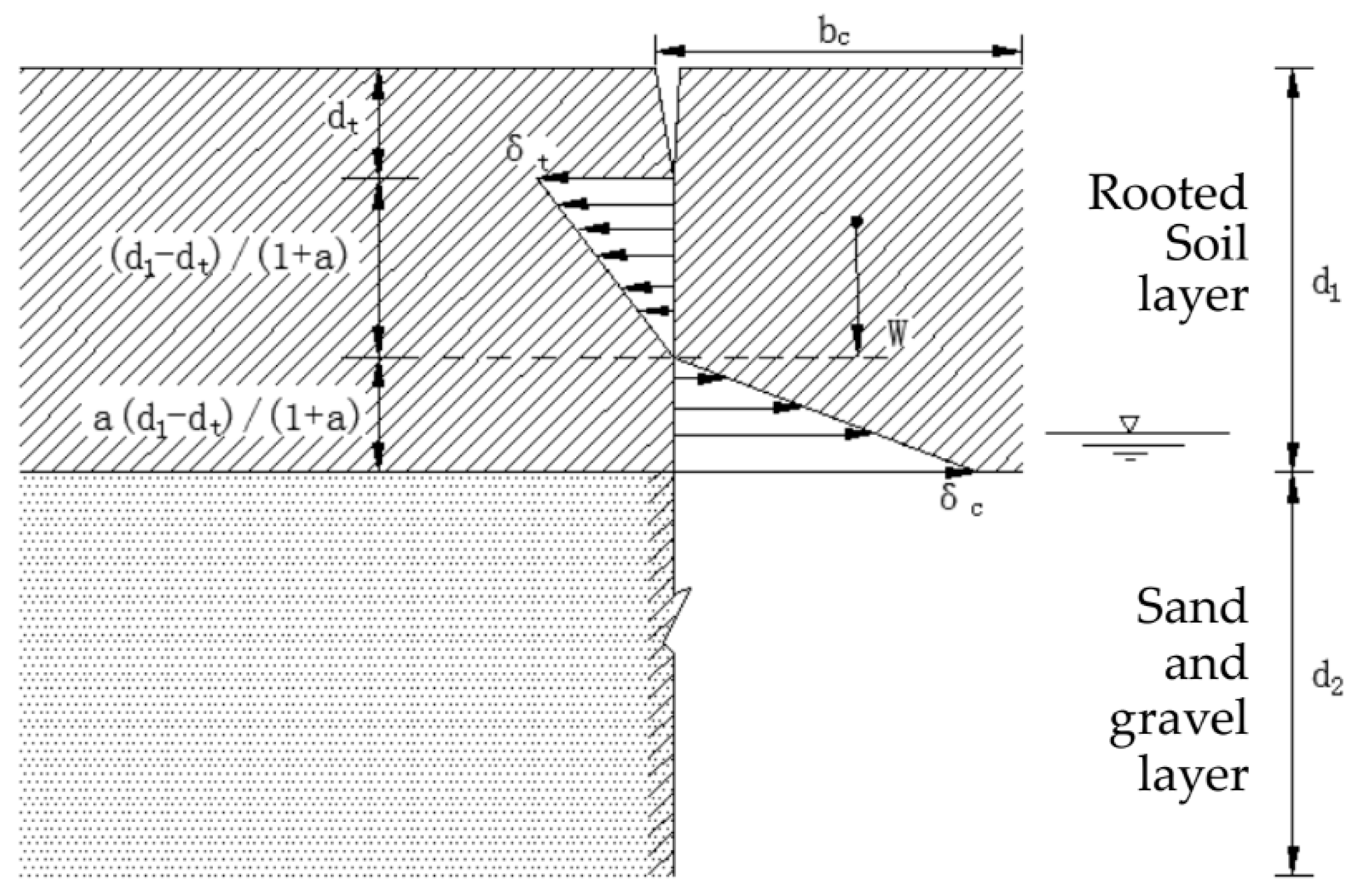
2.3. Calculation
3. Results and Analysis
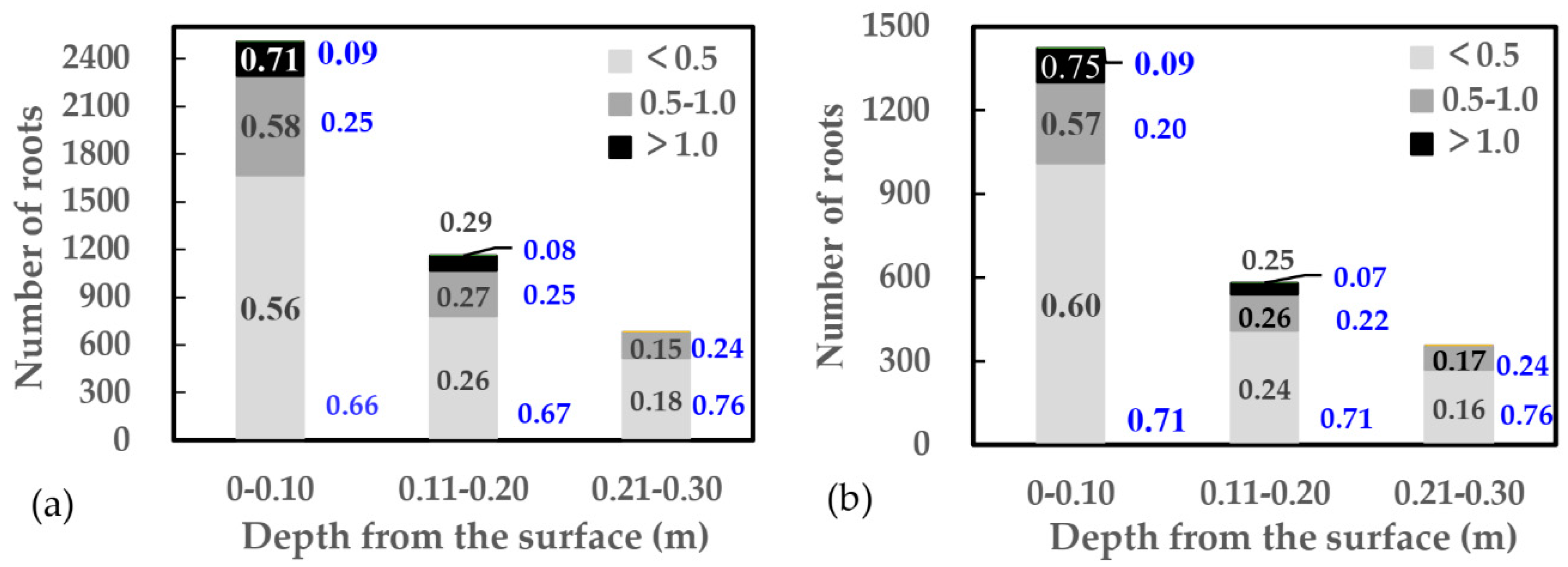
3.1. Characteristics of Soils and Root Distribution
3.2. Tensile Trength of Individual Roots and Soil-Root Mixture
4. Discussion
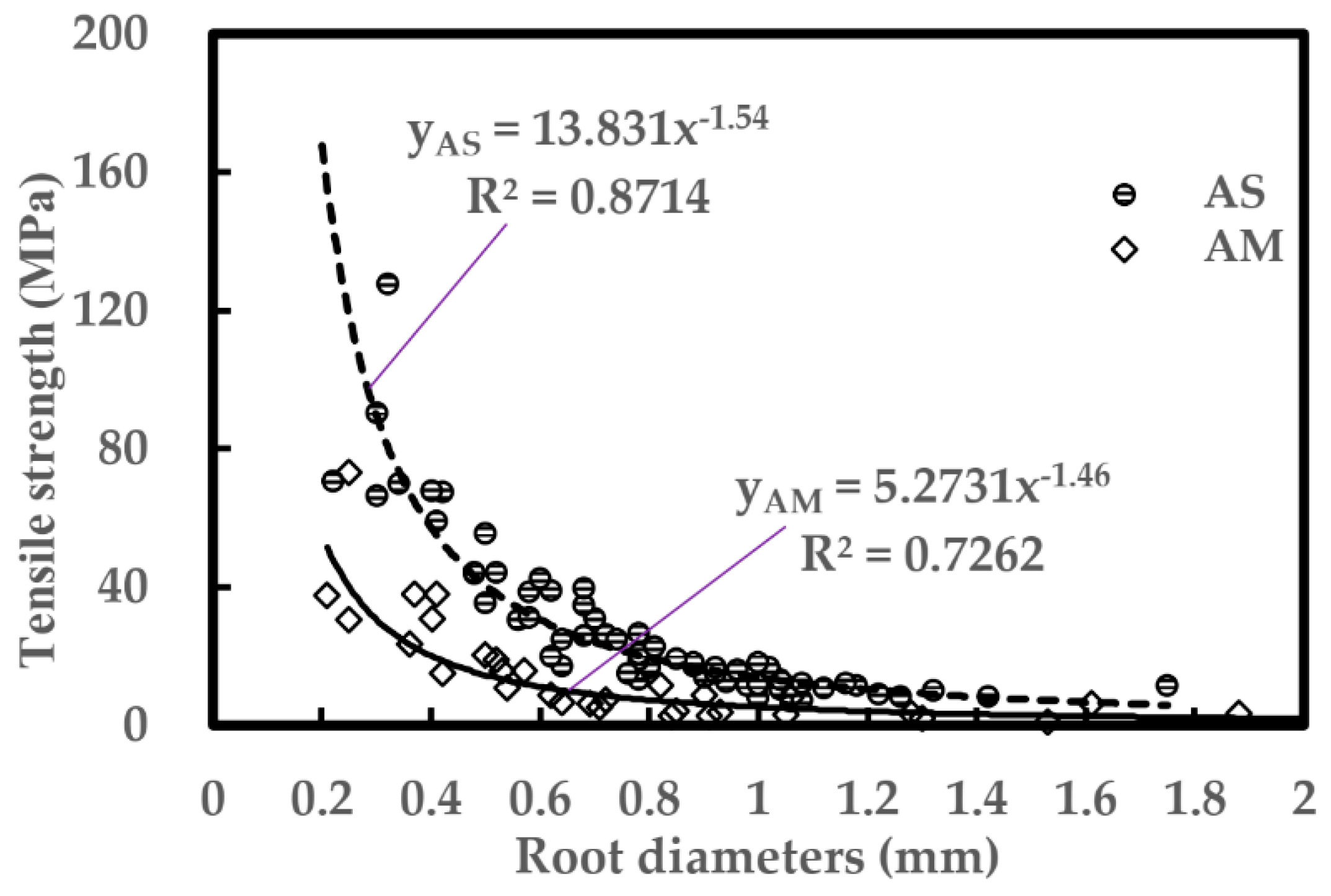
4.1. Effect of Degraded Riparian Vegetation on Tensile Strength of Individual Roots and the Soil-Root Mixture
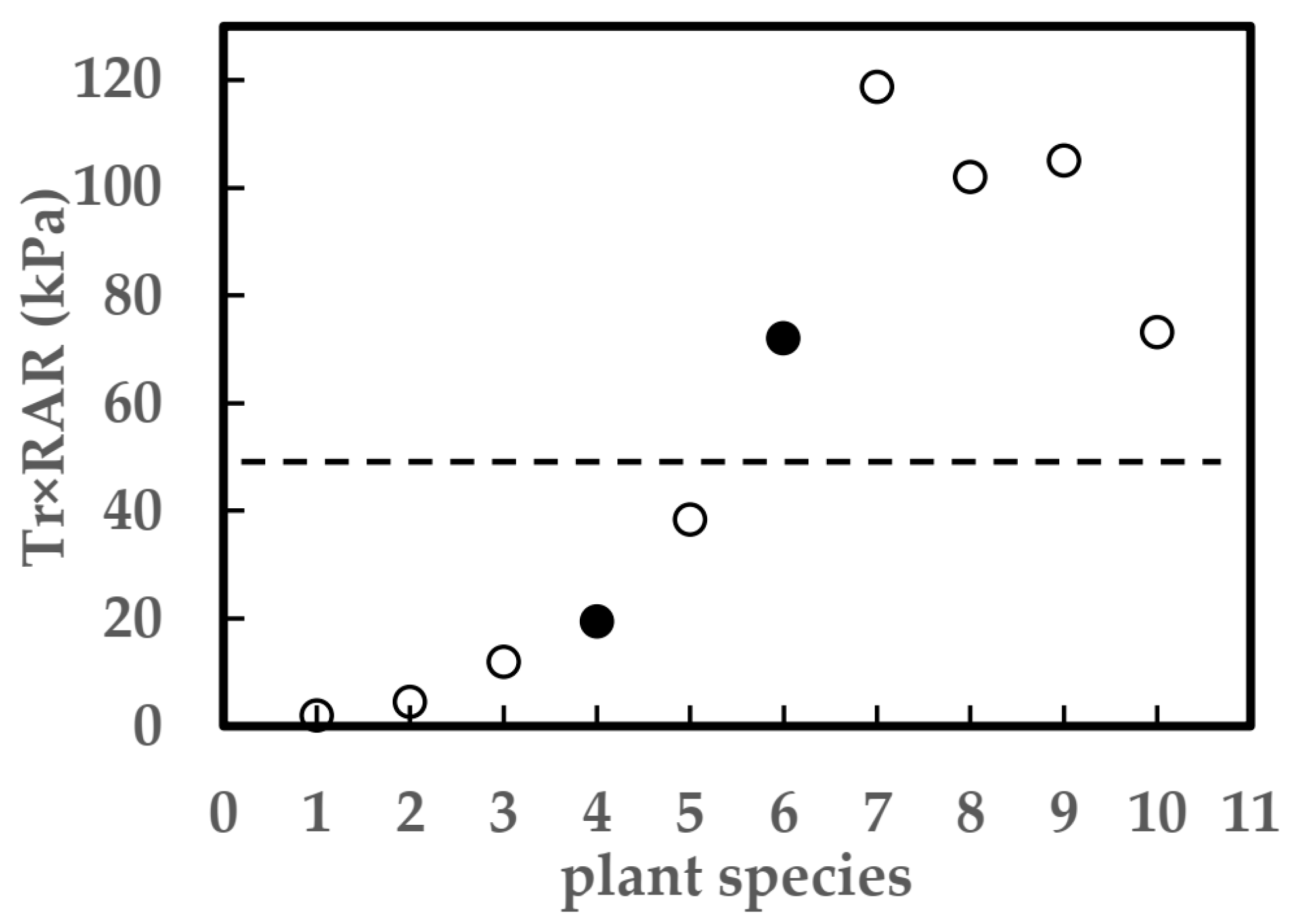
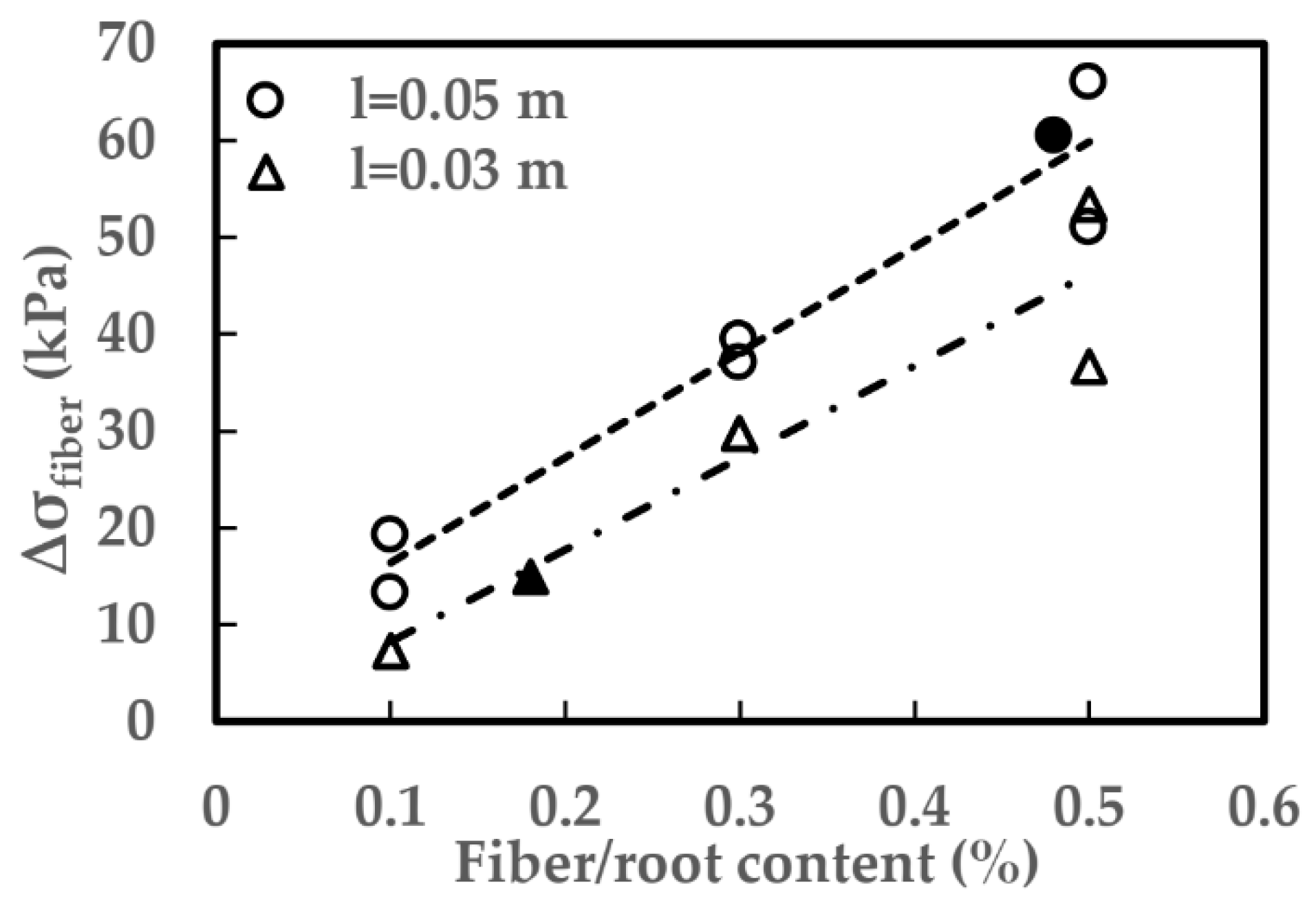
4.2. The Role of Root System in Preventing Development of Riverbank Cracks
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ziegler, S.; Leshchinsky, D.; Ling, H.L.; Perry, E.B. Effect of short polymeric fibers on crack development in clays. Soils Found. 1998, 38, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Li, G.X.; Chen, L. Study of centrifugal model tests on texsol and cohesive soil slopes. Chin. J. Geotech. Eng. 1998, 20, 12–15. (In Chinese) [Google Scholar]
- Tang, C.S.; Wang, D.Y.; Cui, Y.Y. Tensile Strength of Fiber reinforced soil. Mater. Civ. Eng. 2016, 28, 1–13. [Google Scholar] [CrossRef]
- Divya, P.V.; Viswanadham, B.V.S.; Gourc, J.P. Evaluation of tensile strength-strain characteristics of fiber reinforced soil through laboratory tests. J. Mater. Civ. Eng. 2014, 26, 14–23. [Google Scholar] [CrossRef]
- Simon, A.; Curini, A.; Darby, S.E.; Langendoen, E.J. Bank and near-bank processes in an incised channel. Geomorphology 2000, 35, 193–217. [Google Scholar] [CrossRef]
- Schmidt, K.M.; Roering, J.J.; Stock, J.D.; Dietrich, W.E.; Montgomery, D.R.; Schaub, T. The variability of root cohesion as an influence on shallow landslide susceptibility in the Oregon Coast Range. Can. Geotech. J. 2001, 38, 995–1024. [Google Scholar] [CrossRef]
- Roering, J.J.; Schmidt, K.M.; Stock, J.D.; Dietrich, W.E.; Montgomery, D.R. Shallow landsliding, root reinforcement, and the spatial distribution of trees in the Oregon Coast Range. Can. Geotech. J. 2003, 40, 237–253. [Google Scholar] [CrossRef] [Green Version]
- Hubble, T.C.T.; Docker, B.B.; Rutherford, I.D. The role of riparian trees in maintaining riverbank stability: A review of Australian experience and practice. Ecol. Eng. 2010, 36, 292–304. [Google Scholar] [CrossRef]
- Abernethy, B.; Rutherford, I.D. Where along a river’s length will vegetation most effectivelystabilise stream banks? Geomorphology 1998, 23, 55–75. [Google Scholar] [CrossRef]
- Thorne, C.R.; Tovey, N.K. Stability of composite river banks. Earth Surf. Process. Landf. 1981, 6, 469–484. [Google Scholar] [CrossRef]
- Abernethy, B.; Rutherford, I.D. The distribution and strength of riparian tree roots in relation to riverbank reinforcement. Hydrol. Process. 2001, 15, 63–79. [Google Scholar] [CrossRef]
- Simon, A.; Collison, A.J.C. Quantifying the mechanical and hydrologic effects of riparian vegetation on streambank stability. Earth Surf. Process. Landf. 2002, 27, 527–546. [Google Scholar] [CrossRef]
- Yu, G.A.; Li, Z.; Yang, H.; Lu, J.; Huang, H.Q.; Yi, Y. Effects of riparian plant roots on the unconsolidated bank stability of meandering channels in the Tarim River, China. Geomorphology 2020, 351, 106958. [Google Scholar] [CrossRef]
- Van De Wiel, M.J.; Darby, S.E. A new model to analyse the impact of woody riparian vegetation on the geotechnical stability of riverbanks. Earth Surf. Proc. Landf. 2007, 32, 2185–2198. [Google Scholar] [CrossRef]
- Parker, G.; Shimizu, Y.; Wilkerson, G.V.; Eke, E.C.; Abad, J.D.; Lauer, J.W.; Paola, C.; Dietrich, W.E.; Voller, V.R. A new framework for modeling the migration of meandering rivers. Earth Surf. Proc. Landf. 2011, 36, 70–86. [Google Scholar] [CrossRef]
- Zhu, H.L.; Hu, X.S.; Li, Z.W.; Song, L.; Li, K.; Li, X.L.; Li, G.R. The influences of riparian vegetation on bank failures of a small meadow-type meandering river. Water 2018, 10, 692. [Google Scholar] [CrossRef] [Green Version]
- Abernethy, B.; Rutherford, I.D. The effect of riparian tree roots on the mass-stability of riverbanks. Earth Surf. Process. Landf. 2000, 25, 921–937. [Google Scholar] [CrossRef]
- Samadi, A.; Amiri-Tokaldany, E.; Davoudi, M.H.; Darby, S.E. Experimental and numerical investigation of the stability of overhanging riverbanks. Geomorphology 2013, 184, 1–19. [Google Scholar] [CrossRef]
- Lawton, E.C.; Khire, M.V.; Fox, N.S. Reinforcement of Soils by Multi-oriented Geo-synthetic Inclusions. J. Geotech. Eng. 1993, 119, 257–275. [Google Scholar] [CrossRef]
- Plé, O.; Lê, T.N.H. Effect of polypropylene fiber reinforcement on the mechanical behavior of silty clay. Geotext. Geomembr. 2012, 32, 111–116. [Google Scholar] [CrossRef]
- Mesbah, A.; Morel, J.C.; Walker, P.; Ghavami, K. Development of a direct tensile test for compacted earthblocks reinforced with natural fibers. J. Mater. Civ. Eng. 2004, 16, 95–98. [Google Scholar] [CrossRef]
- Onur Akaya, A.; Özera, T.; Foxb, G.A.; Wilson, G.V. Application of fibrous streambank protection against groundwater seepage erosion. J. Hydrol. 2018, 565, 27–38. [Google Scholar] [CrossRef]
- Das, N.; Singh, S.K. Geotechnical behaviour of lateritic soil reinforced with brown waste and synthetic fiber. Int. J. Geotech. Eng. 2019, 13, 287–297. [Google Scholar] [CrossRef]
- Meriem, C.; Houda, G.; Mehrez, J. Tensile behaviour analysis of compacted clayey soil reinforced with natural and synthetic fibers: Effect of initial compaction conditions. Eur. J. Environ. Civ. Eng. 2020, 24, 354–380. [Google Scholar]
- Wu, T.H.; McKinnell, W.P., III; Swanston, D.N. Strength of tree roots and landslides on Prince of Wales Island, Alaska. Can. Geotech. J. 1979, 16, 19–33. [Google Scholar] [CrossRef]
- Waldron, L.J.; Dakessian, S. Soil reinforcement by roots: Calculation of increased soil shear resistance from root properties. Soil Sci. 1981, 132, 427–435. [Google Scholar] [CrossRef]
- Pollen, N.; Simon, A. Estimating the mechanical effects of riparian vegetation on stream bank stability using a fiber bundle model. Water Resour. Res. 2005, 41, 1–11. [Google Scholar] [CrossRef]
- Schwarz, M.; Lehmann, P.; Or, D. Quantifying lateral root reinforcement in steep slopes: From a bundle of roots to tree stands. Earth Surf. Proc. Landf. 2010, 35, 354–367. [Google Scholar] [CrossRef]
- Li, Y.Z.; Fu, J.T.; Yu, D.M.; Hu, X.S.; Zhu, H.L.; Li, G.Y.; Hu, X.T. Mechanical effects of halophytes roots and optimal root content for slope protection in cold and arid environment. Chin. J. Rock Mech. Eng. 2015, 34, 1370–1383. (In Chinese) [Google Scholar]
- Liu, Y.B.; Hu, X.S.; Yu, D.M.; Li, S.X.; Yang, Y.Q. Microstructural features and friction characteristics of the interface of shrub roots and soil in loess area of Xining Basin. Chin. J. Rock Mech. Eng. 2018, 37, 1270–1280. (In Chinese) [Google Scholar]
- Li, Z.W.; Yu, G.A.; Xu, M.Z.; Hu, X.Y.; Yang, H.M.; Hu, S.X. Progress in studies on river morphodynamics in Qinghai-Tibet Plateau. Adv. Water Sci. 2016, 27, 617–628. [Google Scholar]
- Li, J.; Zhang, F.W.; Lin, L.; Li, H.Q.; Du, Y.G.; Li, Y.K.; Cao, G.M. Response of the plant community and soil water status to alpine Kobresia meadow degradation gradients on the Qinghai–Tibetan Plateau, China. Ecol. Res. 2015, 30, 589–596. [Google Scholar] [CrossRef]
- Wang, G.X.; Wang, Y.B.; Li, Y.S.; Cheng, H.Y. Influences of alpine ecosystem responses to climatic change on soil properties on the Qinghai-Tibet Plateau, China. Catena 2007, 70, 506–514. [Google Scholar] [CrossRef]
- Wang, G.X.; Liu, G.S.; Li, C.J. Effects of changes in alpine grassland vegetation cover on hillslope hydrological processes in a permafrost watershed. J. Hydrol. 2012, 444–445, 22–33. [Google Scholar]
- Zeng, C.; Zhang, F.; Wang, Q.J.; Chen, Y.Y.; Joswiak, D.R. Impact of alpine meadow degradation on soil hydraulic properties over the Qinghai-Tibetan Plateau. J. Hydrol. 2013, 478, 148–156. [Google Scholar] [CrossRef]
- Pan, T.; Hou, S.; Wu, S.H.; Liu, Y.J.; Liu, Y.H.; Zou, X.T.; Herzberger, A.; Liu, J.G. Variation of soil hydraulic properties with alpine grassland degradation in the eastern Tibetan Plateau. Hydrol. Earth Syst. Sci. 2017, 21, 2249–2261. [Google Scholar] [CrossRef] [Green Version]
- Krzeminska, D.; Kerkhof, T.; Skaalsveen, K.; Stolte, J. Effect of riparian vegetation on stream bank stability in small agricultural catchments. Catena 2019, 172, 87–96. [Google Scholar] [CrossRef]
- Duan, S.Q. Runoff spatial difference of small-scale in Huangnan, Qinghai province and its cause. Adv. Water Sci. 2016, 27, 11–21. [Google Scholar]
- Yu, Z.X. Investigation of variety resources of Oula sheep at Henan county in Qinghai province. Anim. Husb. Feed Sci. 2009, 30, 120–124. (In Chinese) [Google Scholar]
- Ren, G.H.; Deng, B.; Hou, Y. Changes of community characteristics in the degradation process of the alpine swamp wetland in the Yellow River Source area. Pratacult. Sci. 2015, 32, 1222–1229. (In Chinese) [Google Scholar]
- Xie, B.S.; Zhu, H.L.; Li, B.F.; Hu, X.S. Study on relationship between vegetation spatial distribution and soil properties in the meander riverside in source region of the Yellow River. J. Sediment Res. 2019, 44, 66–73. (In Chinese) [Google Scholar]
- Schenk, H.J.; Jackson, R.B. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. J. Ecol. 2002, 90, 480–494. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Construction. GB/T 50123–1999. Standard for Soil Test Method; Ministry of Construction: Beijing, China, 2010.
- Xia, J.Q.; Zong, Q.L.; Xu, Q.X.; Deng, C.Y. Soil properties and erosion mechanisms of composite riverbanks in Lower Jingjiang Reach. Adv. Water Sci. 2013, 24, 810–820. (In Chinese) [Google Scholar]
- Ajaz, A. Stress-Strain Behaviour of Compacted Clays in Tension and Compression. Ph.D. Thesis, Cambridge University, Cambridge, MA, USA, 1973. [Google Scholar]
- Baets, S.D.; Poesen, J.; Reubens, B.; Wemans, K.; Baerdemaeker, J.D.; Muys, B. Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant Soil 2008, 305, 207–226. [Google Scholar] [CrossRef]
- Mattia, C.; Bischetti, G.B.; Gentile, F. Biotechnical characteristics of root systems of typical Mediterranean species. Plant Soil 2005, 278, 23–32. [Google Scholar] [CrossRef]
- Bischetti, G.B.; Chiaradia, E.A.; Simonato, T.; Speziali, B.; Vitali, B.; Vullo, P.; Zocco, A. Root strength and root area ratio of forest species in Lombardy (Northern Italy). Plant Soil 2005, 278, 11–22. [Google Scholar] [CrossRef]
- Docker, B.B.; Hubble, T.C.T. Quantifying root-reinforcement of river bank soils by four Australian tree species. Geomorphology 2008, 100, 401–418. [Google Scholar] [CrossRef]
- Lawrence, C.J.; Rickson, R.J.; Clark, J.E. The effect of grass roots on the shear strength of colluvial soils in Nepal. In Advances in Hillslope Processes; Anderson, M.G., Brooks, S.M., Eds.; John Wiley and Sons: Chichester, UK, 1996; Volume 2, pp. 857–868. [Google Scholar]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Schwarz, M.; Cohen, D.; Or, D. Soil-root mechanical interactions during pullout and failure of root bundles. Geophys. Res. 2010, 115, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, M.; Preti, F.; Giadrossich, F.; Lehmann, P.; Or, D. Quantifying the role of vegetation in slope stability: A case study in Tuscany (Italy). Ecol. Eng. 2010, 36, 285–291. [Google Scholar] [CrossRef]
- Zhu, J.Q.; Wang, Y.Q.; Wang, Y.J.; Zhang, H.L.; Li, Y.P.; Li, Y. An analysis on soil physical enhancement effects of root system of PinusTabulaeformis and Acer Truncatum based on two models. Bull. Soil Water Conserv. 2015, 35, 277–282. (In Chinese) [Google Scholar]
- Li, X.L.; Perry, G.L.W.; Brierley, G.; Sun, H.Q.; Li, C.H.; Lu, G.X. Quantitative assessment of degradation classifications for degraded alpine meadows (heitutan), Sanjiangyuan, Western China. Land Degrad. Dev. 2014, 25, 417–427. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Wang, G.X.; Hu, H.C.; Wang, Y.B. The variation of soil temperature and water content of seasonal frozen soil with different vegetation coverage in the headwater region of the Yellow River, China. Environ. Geol. 2008, 54, 1755–1762. [Google Scholar] [CrossRef]
- Niu, Y.J.; Zhou, J.W.; Yang, S.W.; Chu, B.; Zhu, H.M.; Zhang, B.; Fang, Q.G.; Tang, Z.S.; Hua, L.M. Plant diversity is closely related to the density of zokor mounds in three alpine rangelands on the Tibetan Plateau. PeerJ 2019, 7, e6921. [Google Scholar] [CrossRef]
- Li, G.R.; Li, X.L.; Chen, W.T.; Li, J.F.; Zhu, H.L.; Hu, X.S.; Zhou, H.K.; Sun, H.Q. Effects of degradation severity on the physical, chemical and mechanical properties of topsoil in alpine meadow on the Qinghai-Tibet Plateau, west China. Catena 2020, 187, 104370. [Google Scholar] [CrossRef]
- Tang, C.S.; Li, J.; Wang, D.Y.; Shi, B. Investigation on the interfacial mechanical behavior of wave-shaped fiber reinforced soil by pullout test. Geotext. Geomembr. 2016, 44, 872–883. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, X.M.; Tian, J.Y. The effectiveness of plant roots in improving soil anti-scourability on the loess plateau. Sci. Bull. 1991, 12, 935–938. (In Chinese) [Google Scholar]
- Zhou, Y.; Watts, D.; Cheng, X.P.; Li, Y.H.; Luo, H.S.; Xiu, Q. The traction effect of lateral roots of Pinus yunnanensis on soil reinforcement: A direct in situ test. Plant Soil 1997, 190, 77–86. [Google Scholar] [CrossRef]
- Li, G.Y.; Hu, X.T.; Li, X.L.; Yu, D.M.; Fu, J.T.; Zhu, H.L.; Hu, X.S. Mechanical Effects of Alpine Grassland Plants in Slope Protection in Maqin County of the Source Area of the Yellow River. Mt. Res. Dev. 2014, 32, 550–560. (In Chinese) [Google Scholar]
- Comino, E.; Druetta, A. The effect of Poaceae roots on the shear strength of soils in the Italian alpine environment. Soil Tillage Res. 2010, 106, 194–201. [Google Scholar] [CrossRef]
- Bessadok, A.; Roudesli, S.; Marais, S.; Follain, N.; Lebrun, L. Alfa fibers for unsaturated polyester composites reinforcement: Effects of chemical treatments on mechanical and permeation properties. Compos. Part A 2009, 40, 184–195. [Google Scholar] [CrossRef]
- Tran, H.N.; Shinji, O.; Nguyen, H.T.; Satoshi, K. Effect of alkali treatment on interfacial and mechanical properties of coir fiber reinforced poly(butylene succinate) biodegradable composites. Compos. Part B 2011, 42, 1648–1656. [Google Scholar]
- Parisi, F.; Asprone, D.; Fenu, L.; Prota, A. Experimental characterization of Italian composite adobe bricks reinforced with straw fibers. Compos. Struct. 2015, 122, 300–307. [Google Scholar] [CrossRef]
- Khiem, Q.T.; Tomoaki, S.; Hiroshi, T. Improvement of mechanical behavior of cemented soil reinforced with waste cornsilk fibers. Constr. Build. Mater. 2018, 178, 204–210. [Google Scholar]
- Tang, C.S.; Shi, B.; Cai, Y.; Gao, W.; Chen, F.J. Experimental study on polypropylene fiber improving soft soils. Rock Soil Mech. 2007, 28, 1796–1800. (In Chinese) [Google Scholar]
- Zhu, C.H.; Liu, J.M.; Yan, B.W.; Ju, J.L. Experimental study on relationship between tensile and shear strength of unsaturation clay earth material. Chin. J. Rock Mech. Eng. 2008, 27 (Suppl. S2), 3453–3458. (In Chinese) [Google Scholar]
- Li, B.F.; Zhu, H.L.; Xie, B.S.; Luo, L.Y.; Li, G.R.; Hu, X.S. Study on tensile properties of root-soil composite of alpine meadow plants in the riparian zone of the Yellow River source region. Chin. J. Rock Mech. Eng. 2020, 39, 424–432. (In Chinese) [Google Scholar]
- Leung, A.K.; Garg, A.; Ng, C.W.W. Effects of plant roots on soil–water retention and induced suction in vegetated soil. Eng. Geol. 2015, 193, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Ng, C.W.W.; Leung, A.K.; Woon, K.X. Effects of soil density on grass-induced suction distributions in compacted soil subjected to rainfall. Can. Geotech. J. 2014, 51, 311–321. [Google Scholar] [CrossRef]
- Gabrm, A.; Akran, M.; Taylor, H.M. Effect of simulated roots on the permeability of silty soil. Geotech. Test. J. 1995, 18, 112–115. [Google Scholar]
- Huat, B.B.K.; Alif, H.J.; Low, T.H. Water infiltration characteristics of unsaturated soil slope and its effect on suction and stability. Geotech. Geol. Eng. 2006, 24, 1293–1306. [Google Scholar] [CrossRef]
- Ng, C.W.W. Atmosphere-plant-soil interactions: Theories and mechanisms. Chin. J. Geotech. Eng. 2017, 39, 1–47. (In Chinese) [Google Scholar]
- Pollen, N.; Simon, A. Hydrologic and hydraulic effects of riparian root networks on streambank stability: Is mechanical root-reinforcement the whole story? Geomorphology 2010, 116, 353–362. [Google Scholar] [CrossRef]
- Morris, P.H.; Graham, J.; Wiliams, D.J. Cracking in drying soils. Can. Geotech. J. 1992, 29, 263–277. [Google Scholar] [CrossRef]
- Albrecht, B.A.; Benson, C.H. Effect of desiccation on compacted natural clays. J. Geotech. Geoenviron. 2001, 127, 67–75. [Google Scholar] [CrossRef]
- Peron, H.; Hueckel, T.; Laloui, L.; Hu, L.B. Fundamentals of desiccation cracking of fine-grained soils: Experimental characterization and mechanisms identification. Can. Geotech. J. 2009, 46, 1177–1201. [Google Scholar] [CrossRef]
- Tang, C.S.; Shi, B.; Liu, C.; Zhao, L.Z.; Wan, B.J. Influencing factors of geometrical structure of surface shrinkage cracks in clayey soils. Eng. Geol. 2008, 101, 204–217. [Google Scholar] [CrossRef]
- Miller, C.J.; Rifai, S. Fiber reinforcement for waste containment soil liners. J. Environ. Eng. 2004, 130, 891–895. [Google Scholar] [CrossRef]
- Muller, S.; Dutoit, T.; Alard, D.; Grévilliot, F. Restoration and rehabilitation of species-rich grassland ecosystems in France: A review. Restor. Ecol. 1998, 6, 94–101. [Google Scholar] [CrossRef]
- Carrick, P.J.; Krüger, R. Restoring degraded landscapes in lowland Namaqualand: Lessons from the mining experience and from regional ecological dynamics. J. Arid Environ. 2007, 70, 767–781. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Type | Coverage (Mean ± SD *, %) | Number of Species (Mean ± SD) | Under-Ground Biomass (g m−2) | Dominant Species | Number of Surveyed Sites |
|---|---|---|---|---|---|
| AS | 96.3 ± 2.7 a | 3.5 ± 2.7 b | 378 ± 74 a | Blysmus sinocompressus | 40 |
| AM | 74.5 ± 11.6 b | 14.8 ± 5.1 a | 193 ± 56 b | Kobresia pygmaea | 44 |
| Elymus nutans | |||||
| Potentilla saundersiana |
| Tested Site | Sampling Depth (m) | Soil Type | Bulk Density ρ (kg·m−3) | Moisture Content ω (%) | RAR (%) |
|---|---|---|---|---|---|
| 1(AS) | 0.10–0.20 | Silt | 1560 | 41.71 | 0.22 |
| 0.30–0.40 | Silty sand | 1730 | 10.29 | - | |
| 0.10–0.20 | Silt | 1550 | 40.92 | 0.23 | |
| 0.30–0.40 | Silty sand | 1740 | 10.97 | - | |
| 2(AM) | 0.10–0.20 | Silt | 1260 | 40.42 | 0.12 |
| 0.40–0.50 | Silty sand | 1710 | 24.25 | - | |
| 0.10–0.20 | Silt | 1290 | 39.36 | 0.11 | |
| 0.40–0.50 | Silty sand | 1650 | 22.24 | - |
| Tested Site | Tensile Strength of Root Tr (kPa) | Thickness of Slump Block d1 (m) | Crack Depth dt (m) | Width of Slump Block bc (m) | Volume of Slump Block V (m3) | Tensile Strength Based on Formula (3) σt (kPa) |
|---|---|---|---|---|---|---|
| 1 | 31,670 | 0.25 | 0.035 | 0.70 | 0.175 | 66.86 |
| 30,950 | 0.26 | 0.037 | 0.72 | 0.187 | 67.93 | |
| mean | 31,310 | 0.255 | 0.036 | 0.71 | 0.181 | 67.395 |
| 2 | 15,420 | 0.35 | 0.048 | 0.52 | 0.182 | 21.29 |
| 16,890 | 0.35 | 0.050 | 0.53 | 0.186 | 22.63 | |
| mean | 16,155 | 0.35 | 0.049 | 0.525 | 0.184 | 21.96 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, H.; Gao, P.; Li, Z.; Fu, J.; Li, G.; Liu, Y.; Li, X.; Hu, X. Impacts of the Degraded Alpine Swamp Meadow on Tensile Strength of Riverbank: A Case Study of the Upper Yellow River. Water 2020, 12, 2348. https://0-doi-org.brum.beds.ac.uk/10.3390/w12092348
Zhu H, Gao P, Li Z, Fu J, Li G, Liu Y, Li X, Hu X. Impacts of the Degraded Alpine Swamp Meadow on Tensile Strength of Riverbank: A Case Study of the Upper Yellow River. Water. 2020; 12(9):2348. https://0-doi-org.brum.beds.ac.uk/10.3390/w12092348
Chicago/Turabian StyleZhu, Haili, Peng Gao, Zhiwei Li, Jiangtao Fu, Guorong Li, Yabin Liu, Xilai Li, and Xiasong Hu. 2020. "Impacts of the Degraded Alpine Swamp Meadow on Tensile Strength of Riverbank: A Case Study of the Upper Yellow River" Water 12, no. 9: 2348. https://0-doi-org.brum.beds.ac.uk/10.3390/w12092348