A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production




1
Center for Midstream Management and Science, Lamar University, Beaumont, TX 77710, USA
2
Environmental Science and Engineering Department, Indian Institute of Technology Bombay, Powai, Mumbai 400076, Maharashtra, India
3
Department of Civil and Environmental Engineering, Lamar University, Beaumont, TX 77710, USA
*
Author to whom correspondence should be addressed.
Water 2020, 12(9), 2351; https://0-doi-org.brum.beds.ac.uk/10.3390/w12092351
Submission received: 27 July 2020
/
Revised: 17 August 2020
/
Accepted: 19 August 2020
/
Published: 21 August 2020
(This article belongs to the Special Issue Advanced Technologies for Produced Water Management, Treatment, and Reuse)
Abstract
:Produced water (PW), the largest waste stream generated in oil and gas industries, has the potential to be a harmless product rather than being a waste. Biological processes using microorganisms have proven useful to remediate PW contaminated by petroleum hydrocarbons, complex organic chemicals, and solvents. In particular, the bioremediation of PW using algae is an eco-friendly and low-cost approach due to algae’s ability to utilize certain pollutants as nutrient sources. Therefore, the utilization of PW as an algal growth medium has a great potential to eliminate chemicals from the PW and minimize the large volumes of freshwater needed for cultivation. Although several reviews describing the bioremediation of PW have been published, to the best of our knowledge, no review has exclusively focused on the algae-based PW treatment. Therefore, the present review is dedicated to filling this gap by portraying the many different facets of the algae cultivation in PW. Several algal species that are known to thrive in a wide range of salinity and the critical steps for their cultivation in hypersaline PW have been identified. Overall, this comprehensive review highlights the PW bioremediation using algae and brings attention to utilizing PW to grow biomass that can be processed to generate biofuels and useful bioproducts.
Keywords:
salinity; hypersaline; pretreatment; bioremediation; microalgae; halophilic; adaptation; nutrient; post-processing; bioproduct1. Introduction
Oil and gas industries generate a significant amount of wastewater along with oil and gas, to the surface, lifting from the subsurface. This geologic water is known as produced water (PW). All over the world, approximately 250 million barrels of PW are generated per day from gas and oil industry and more than 40% of these are introduced into the environment posing a threat of severe environmental pollution [1,2,3,4]. PW has varying concentrations of contaminants such as hydrocarbons, heavy metals, and additives that, if not treated properly, can significantly contaminate the receiving water bodies, soil and air [5,6]. The composition of PW varies with the geologic age, depth, and geochemistry of the hydrocarbon-bearing region along with the chemical composition of crude oil and natural gas in the particular zone and the chemicals added during the exploration process. Traditionally, chemical, physical, and mechanical methods have been used to treat PW. Major physico–chemical methods include thermal technologies (multi-stage flash distillation, vapor compression distillation, freeze–thaw evaporation) [7], floatation [8] and density-based separation [2], adsorption technologies [9], membrane filtration technologies [10], chemical precipitation, chemical oxidation, and electrochemical methods [11]. These methods suffer from high cost and energy input (thermal methods), scale formation issues (membrane, thermal methods), extensive pre- and post-treatment requirements (thermal methods, membrane methods, adsorption), sludge disposal (chemical precipitation, electrochemical systems) and poor efficiency (membrane methods, electrochemical methods). Moreover, these methods are often complicated since they depend substantially on the certain characteristics of the effluent [2,6].
On the other hand, biological treatment methods are an economical and effective method that can remove harmful chemicals and pollutants from the environment [12]. It is well known that many microorganisms such as algae, bacteria, fungi, yeast etc. can grow in highly saline media, which qualify them for the bioremediation of effluents such as PW [13]. Therefore, the use of microorganisms, such as algae for the purification of PW, represents an important advantage in the bioremediation process. Algae is capable of removing pollutants such as nutrients, heavy metals, and dissolved and complex organic chemicals from various types of wastewater such as municipal wastewater [14,15,16], landfill leachate [17], anaerobic digestion effluent [18] and sewage waste [19]. Generally, algae remove toxic elements through mechanisms such as biosorption and bioaccumulation [20,21,22,23,24]. Negatively-charged extracellular polymeric substances (EPS) of microalgae are also known for metal biosorption [20,24,25]. Microalgae need some of the elements present in PW for their growth [4]. Therefore, the cultivation of microalgae in PW could significantly clean up this effluent. Moreover, the algal approach can also lead to a more sustainable approach in PW remediation since it incorporates resource recovery in the treatment model by producing biomass that can serve as raw materials for biofuel, bioactive compounds, and nutrients supplement production [26,27].
Although PW contains some toxic components that may inhibit algae growth, it also contains some nutrients such as nitrogen and phosphorus, in the form of ammonium and phosphate, respectively, that are required for biomass cultivation [28]. These vital nutrients are generally available in PW at an adequate level for algae growth and often at levels more than that in the commercial growth media [28,29,30,31]. Since the nutrient addition can represent ~50% of the algae cultivation costs [12], the presence of nitrogen and phosphorus improves the overall economy and return on investment. Therefore, the cultivation of microalgae in PW is an efficient means to remove nutrients from PW. Additionally, the algal biomass can be transformed into valuable products, such as biofuels (e.g., biodiesel and bioethanol).
Generally, brackish groundwater is widely used to cultivate many species of algae due to its wide range of total dissolved solids (TDS). Therefore, PW, often found to be in the brackish TDS range, can be a great resource for algae cultivation [32]. Hence, the proper utilization of PW could provide cost-effective advantages to current PW disposal methods by producing biofilm biomass and associated high-value products such as phycocyanin. Commonly, algal biofilms are used to produce various valuable products such as bioplastics, pharmaceutically active compounds, high-value chemicals, biofuel feedstock, and animal feed [33]. In addition, phycocyanin, a major blue phycobiliprotein pigment found in cyanobacteria, has numerous potential applications in biotechnology, medicine, cosmetics, and food applications [34,35]. For example, it is widely used as a label for fluorescence diagnostics and immunoassays, owing to its specific and intense fluorescent properties [36]. However, the production of this high-value bioproduct is currently dominated by the outdoor culture in open ponds and raceways, which require large volumes of growth medium and include expensive post-processing operations such as harvesting and drying [34].
Algae-based biofuel production has received considerable attention in recent years owing to its biodegradability and sustainability. The carbohydrate and oil contents of algae allow them to be considered as a potential feedstock to produce biofuels. For example, algae biomass could be converted to bio-crude by hydrothermal liquefaction (HTL) that mimics fossil fuel generation [37,38]. Microalgae produce mainly non-polar lipids such as triacylglycerols/triglycerides and polar lipids such as phospholipids [39]. These lipids are excellent sources for biofuels production. Generally, non-polar lipid forms such as mono-, di-, and triacylglycerols (MAGs, DAGs, and TAGs, respectively), and free fatty acids (FFAs) can be converted into biodiesel (i.e., fatty acid methyl esters—FAMEs) through transesterification process [39]. However, triacylglycerols (TAGs) are the most desirable form for conversion into biodiesel due to the high yield of FAME production (i.e., 3 moles of FAME per mole of TAG). Many microalgae are known to accumulate triacylglycerols under stressed conditions (e.g., high salinity, nutrient deficiency, excess light, etc.) as a storage of carbon and energy. While various types of wastewater have been widely used as media to cultivate algae, PW is not well explored in this context. While a limited number of studies utilized PW explicitly for algae cultivation, only a few determined the lipid production [11,40]. Therefore, a review of such exploration is needed to identify potential challenges and shed light on future perspectives.
Many studies have been conducted in recent years into using PW for algal-based biomass and biofuel production [41]. However, to the best of our knowledge, there have been no systematic reviews that holistically reported the cultivation of microalgae in PW and discussed the potential of bioremediation and biomass and biofuels production. Moreover, there is little information available regarding the availability, quality, toxicity, and overall feasibility of PW for the cultivation of algae. Therefore, the primary objectives of this review are to demonstrate the potential of algae to (i) produce biomass and biofuels utilizing the PW as the source of nutrients, and (ii) remediate contaminants from PW. In the beginning, a comprehensive literature search strategy has been followed to determine the current trend in this field and the scope of the review. Later, the review extensively summarizes recent findings related exclusively to PW and algae and discusses the different methodologies developed by researchers to exploit the algae–PW system. Finally, the review sheds light on the post-processing of both PW and algae, the factors affecting the large-scale development of this system, and associated challenges.
Furthermore, a special focus is given to the cultivation of algae in PW for biomass production. Interestingly, the current work is unique in identifying the major steps to promote the cultivation of algae in PW. We also collected the various algal species from the literature and provided a breakdown of freshwater and marine strains that have been deployed in PW systems. These strains can be further researched, and the breakdown can be used to identify other potential strains that can thrive in PW. Additionally, we provided an exclusive list of algal species that are known to thrive in a wide range of salinity; therefore, they can be explored in hypersaline PW treatment.
A comprehensive literature search strategy was done through the Web of Science, Google Scholar, ProQuest database and Produced Water Society (PWS) seminars. From the literature survey, we observed that although the algae-based PW treatment system or PW-based algae cultivation system is not new, there has not been much research effort spent on either system before 2014. Moreover, while the number of research articles published every year increased since then, the overall efforts made are still meager. In addition to review articles and patents, the increasing number of research articles and citations, as primary indicators of the growing research interest in this field, have inspired the current review.
2. Algae-Based PW Systems
2.1. Cultivation of Algae in PW for Biomass Production
The characteristics of PW depend on the geological formation of the reservoirs, the procedure used in the extraction, and the age of the oil well [42]. Generally, the complex chemical composition of PW makes the cultivation of algae challenging and often requires treatment beyond oil/water separation to make it suitable for algal growth. Nevertheless, PW has high levels of salinity and has nutrients such as nitrogen and potassium that can facilitate algae growth. Therefore, PW can be used to grow algae either as a standalone media or by mixing with other media. Table 1 presents a comprehensive list of parameters with a typical range to provide a general overview of PW characteristics.
Undoubtedly, the chemical and physical properties of PW show significant variation, as indicated by the wide range of the parameters listed above, and no two region’s PW can be alike. Therefore, multiple steps such as the pre-treatment of PW, selection of algal species based on the specific characteristics of PW and adaptation of algae in PW, and their combinations are often followed to successfully cultivate algae using PW.
2.1.1. Pre-Treatment of PW
Since PW contains various concentrations of free oil, dissolved solids, metals, organic contaminants, volatile organic compounds (VOCs), and precipitates of scale-forming minerals that can interfere with algae growth, pre-treatment of PW prior to media mixing is often required [40]. For instance, Al-Ghouti et al. filtered PW from a natural gas field in Qatar using a 0.45 μm millipore filter to remove most of the suspended solids and other significant pollutants [11]. The PW was then used to effectively grow five chlorophytes that effectively removed several heavy metals such as Al, Zn, and Fe. Similarly, Godfrey pre-treated PW by centrifugation, settling, and activated carbon filtration and observed significant increase in algal growth and lipid productivity [47]. In particular, the author centrifuged the PW to remove visible black hydrocarbons that settled in a black mass [48]. The study then reported that pre-treatment helped to increase the lipid accumulation in the algal biomass. However, since centrifugation in a large, commercial process is too costly activated carbon can be used as an alternative. Godfrey compared both methods of hydrocarbon removal and reported that activated carbon filtration was more effective than centrifugation in terms of hydrocarbon removal from PW. Therefore, activated carbon filtration can be a viable technique for hydrocarbon removal, especially in large-scale algal systems using PW [48].
Among other pre-treatment methods, Ranjbar et al. used sterilization of PW at 121 °C for 15 min before inoculating Dunaliella salina. In the absence of sterilization, complete inhibition of the growth by antagonistic microorganisms was recorded [39]. Therefore, sterilization should be a part of the pre-treatment of PW not only to inhibit the growth of microorganisms but also to inactivate inhibitory chemicals, decompose organic contaminants, and precipitate toxic metals [39]. However, since sterilization is not feasible in large-scale cultivation, algal growth can often be challenged. For instance, Winckelmann et al. used pre-cleaned PW to grow an indigenous isolate Scenedesmus sp. in open ponds under semi-continuous conditions [49]. Although the pre-cleaning process was not elaborately described, the group reported weed algae (cyanobacteria and diatoms) contamination of the produced biomass that led to a negative effect on photosynthetic efficiency of the cultures over time [49]. Another challenge of using PW could arise from precipitate formation. For example, Neal et al. cultivated Nannochloropsis salina in treated PW collected from Eldorado Biofuels, Jal, New Mexico, that was then filtered using a 0.22 μm filter [32]. Nevertheless, the group reported high optical density data without a scaled amount of chlorophyll. This indicated that some precipitates, which formed from high sulfate concentrations as were shown by the scanning electron microscope (SEM) image and energy-dispersive X-ray spectroscopy (EDAX) spectra, may have fallen out of the PW even after filtration. Such precipitates in PW can absorb the light that the algae need and thus inhibit the growth.
2.1.2. Selection of Algal Species Based on the Salinity of PW
Additional challenges of cultivating algae in PW comes from its high salinity. Generally, freshwater algae do not seem to grow well in PW with high salinity. The unpublished work from Badrinarayanan showed that freshwater algae Chlamydomonas reinhardtii grows in 20 times diluted PW i.e., at low salinity [50]. The work also reported that the growth rates get better when grown along with Tris-Acetate-Phosphate (TAP) medium. Although C. reinhardtii did not seem to grow well in this study, it must be noted that the PW used in the study had an exceptional level of salinity. Badrinarayanan pre-treated PW using centrifugation and vacuum filtration to remove the solids, yet the final, clear PW had a salinity of ~147,000 mg L−1. Among the published work, Johnson et al. recently identified Scenedesmus rotundus, another freshwater species, that caused bio-fouling of an oil field PW treatment facility. As the research group conducted a site audit, they reported a high bicarbonate content likely from oil biodegradation and a high phosphate influent from a storm drain. The group attributed these findings to the growth of the colony-forming S. rotundus, that initially manifested as a slime which then aggregated to form slime mats after flocculants were added, and finally led to the biofouling of the PW treatment system [40,51]. Initial analysis of the slime by light microscopy indicated the slime to be microbial in origin. Furthermore, the species level classification of algal sequences under the classification Plantae from the slime sample, indicated that 70% of plastid sequences belonged to the species S. rotundus. This alga is known to be found in a wide range of sub-tropical and desert locations, which indicates the climate (i.e., elongated periods of desiccation and tropical rainfall) experienced by this microalgal species at the PW treatment plant [52,53,54,55,56]. The salinity of the PW treatment plant at the time of sampling was between 5021 and 7294 mg L−1, which typically is in the range of moderately saline water. Other research has corroborated that Scenedesmus sp. can grow optimally at salinities between 0 and 10,000 mg L−1, with growth possible but inhibited between 10,000 and 30,000 mg L−1 [52]. These research findings indicate that some freshwater algae such as the C. reinhardtii, S. rotundus, and Scenedesmus sp. can tolerate moderate salinity.
Alternatively, marine algal strains can withstand even higher salinity and thus, have a competitive advantage over their freshwater counterparts. For example, marine microalgae species Nannochloropsis sp. Showed a higher growth and biomass production rate than C. reinhardtii at the same dilution (20 times) of PW, as reported by Badrinarayanan. Another Nannochloropsis species; Nannochloropsis oculata, showed great potential for cultivation in culture media with 50% PW and produced significant biomass in rates similar to those achieved in traditional culture systems [42]. The group reported that the culture in 100% PW was also possible, although the biomass yield was low. A previous study has reported that Nannochloropsis sp. has a peak salinity tolerance of around 40 g TDS L−1 [57]. Another marine algae Dunaliella sp. is known to tolerate salt concentrations ranging from 5000 to 359,000 mg L−1 [39]. Hopkins et al. cultivated Dunaliella tertiolecta, another marine species, in PW with a wide range of salinities 30–210 g TDS L−1 and reported similar biomass productivity (i.e., biomass accumulation rate) for salinities between 30 and 120 g TDS L−1 while salinity-induced growth inhibition occurred at ≥180 g TDS L−1. Generally, the genus Dunaliella is very well known for its ability to thrive in saline to extreme hypersaline environments. This species has also been reported to grow in brackish coal gas seam and saline hydraulic fracturing flowback water [58,59].
Hopkins et al. further correlated the growth with nutrient removal (phosphate and nitrate) at various salinity of the PW media. At lower salinity conditions (30–210 g TDS L−1), nitrate was removed by days eight to nine, whereas at the higher salinity conditions, this did not occur until day 11. In either case, phosphate was removed one or two days earlier. The authors also reported that the most extreme hypersaline conditions (210 g TDS L−1) required the highest phosphate and nitrate nutrient amounts, which is likely due to greater nutrient and energy requirements for growth at this extreme ionic strength [60]. Nevertheless, these experimental findings indicate D. tertiolecta and other species in the genus as potential candidates that can tolerate the challenging ionic strengths typical of PW.
Very recently, Hopkins et al. discovered two freshwater species Cyanobacterium aponinum and Parachlorella kessleri dominated polyculture in PW originating from a production facility in the Permian Basin of southwestern New Mexico [41]. This polyculture demonstrated great potential for growth in a wide range of PW salinities such as 15, 30, and 60 g TDS L−1. Although the growth reduced at salinities of 75 and 90 g TDS L−1 and little to no biomass was produced in salinities ≥ 105 g TDS L−1, however, dilution to 60 g TDS L−1 was able to restart the growth. Therefore, the researchers concluded that both C. aponinum and P. kessleri were able to grow in highly saline PW, with an upper halotolerance of ≥60 g TDS L−1. More interestingly, when the authors compared the growth rates in PW to the f/2 medium, the polyculture displayed higher initial growth rates in all PW conditions (≤ 60 g TDS L−1) than that in the f/2 medium. The authors attributed this finding to the 10 times higher alkalinity due to higher levels of bicarbonate (~20 mM) in PW based media, compared to that (~2 mM) of the seawater-based f/2 medium. Table 2 presents detailed information on the algal species that have been deployed in PW for bioremediation and biomass generation purposes at various cultivation and experimental conditions.
Figure 1a shows the number of algal species per phylum that have been explored for their ability to thrive in PW and Figure 1b presents the breakdown of freshwater and marine algal strains among the explored algae. It is no wonder that Chlorophyta and Cyanobacteria are the most explored phyla. It is evidenced that marine strains have been less explored than the freshwater strains. However, our current discussion indicates that exploring marine strains is worth more than freshwater strains due to the hypersalinity of PW. Therefore, Table 2 and Figure 1 combined can be used to identify algal species that have already been known to thrive in PW and conduct their feasibility for large-scale PW systems and explore more species, especially halotolerant marine strains.
2.1.3. Adaptation of Algae in PW
In addition to the pre-treatment of PW and choosing a halotolerant species, it is equally crucial to allow algae to adapt in PW for achieving optimal growth. For instance, Ammar et al. cultivated marine species N. oculata and Isochrysis galbana in a mixture of PW and modified BG-11 in three adaptation stages: 10%, 25%, and 50% PW media. In every stage, the group repeated the cultivation experiment twice to allow microalgae to adapt with the new PW environment. They reported that by increasing the PW loading, the period of the lag phase was prolonged for both N. oculata and I. galbana. This indicated that the microalgae took a longer time to adapt to the new conditions when the PW concentration was gradually increased. As the authors compared the growth of the microalgae in PW with the control (0% PW), they observed that both strains showed a noticeable improvement in the second experiment of each adaptation stage. This finding indicated that both N. oculata and I. galbana were able to adapt to new and low-nutrition conditions. However, the final biomass yields decreased with increasing PW loading. The biomass concentration dropped by ~70% for both strains when the effluent loading increased from 10% to 50%. This suggests inhibition of microalgal growth in high PW loadings. In addition, at higher PW concentrations, microalgae flocculated in the flask bottom, which is due to the presence of metal ions that neutralized the cell surface. Nevertheless, both microalgae adapted substantially in PW. N. oculata yielded a higher biomass than I. galbana in low PW loadings, whereas I. galbana showed an improvement in biomass yield in high PW loading (50%) due to the high salinity of PW media which is favorable for its growth. In a similar study, Godfrey successfully grew Synechocystis sp. in PW by periodically adding concentrated sodium nitrate to the culture. It was observed that Synechocystis sp. grew easily in diluted PW if given enough nitrate. These studies, therefore, indicate a possible adaptive advantage of preadapted cells in PW.
In a very recent study, Calderón-Delgado explored the effect of PW on the cell diameter of Chlorella vulgaris. A significant negative correlation was reported as they observed a reduction in cell diameter in PW experiments with respect to the control experiment [61]. The diameter reduced more as the PW concentration increased, which is probably due to an adaptive mechanism for the number of xenobiotics present in PW. The small cell diameter of C. vulgaris is an advantage to reduce the levels of xenobiotics and increase biotransformation. Generally, smaller cells have a bigger specific surface area that allows the algae to absorb xenobiotics effectively [66,67]. This adaptive response to the toxicity can also be correlated with the chlorophyll content by a lower volume due to the degradation of pigment.
2.1.4. Addition of Media Mixes and Nutrients to PW
Although adding standard media components and nutrients to PW has become a very common practice, there are challenges associated with such additions [40]. Such additions can often add excess trace metals and anions that may result in the precipitation of multiple constituents in various forms. For example, Ca2+ mixed with high SO42− water and subsequent evaporation leads to precipitation of gypsum [40]. Moreover, significant losses of mineral constituents occurred when the same researchers modeled a medium mixing process to create a typical saline f/2- Si medium (35,000 mg TDS L−1) using standard media mixes of chemically pure compounds and brackish PW from the Jal, New Mexico, test well [40]. The group reported the formation of fluorapatite (Ca5(PO4)3F), calcite (CaCO3), huntite (CaMg3(CO3)4) etc. when insoluble amounts of calcium and phosphorous were added to the culture media. These minerals consisted of 99% of the mass of precipitated minerals, while the precipitation occurred as a function of temperature (typical ranges for southern New Mexico and west Texas). In contrast, nutrient losses could have been reduced if the mix was adjusted to accommodate the existing elements in solution. In fact, the media can be optimized to suppress precipitation by selectively removing excess divalent ions such as Ca2+, SO42− or that are beyond the need for algae growth. This reduces the loss of critical constituents, including Co, Fe, K, Mg, Mn etc. Therefore, chemical balancing of the media mix can prevent precipitation and minimize nutrient costs.
As previously discussed, the pre-treatment of the PW can potentially reduce or remove some of the contaminants and allow better growth conditions for algae. For example, the removal of suspended solids and colloidal materials can allow better sunlight penetration, and consequently, more algal biomass production [4]. For instance, Das et al. investigated the growth viability of three freshwater strains (Chlorella sp., Scenedesmus sp., and Monoraphidium sp.) and three marine microalgal strains (Tetraselmis sp., Picochlorum sp., Lyptolyngbya sp.) in the pre-treated PW, presented in Table 2. The group pre-treated the PW by adding NaOH to adjust the pH from 4.2 to 7.1 and centrifuging to remove suspended and colloidal materials [4]. The pre-treated PW was used as a growth media in two conditions (i) with N and P supplementation and (ii) without additional nutrients. Out of these microalgae, only Chlorella sp. and Scenedesmus sp. were able to grow in both conditions. Although the pre-treated PW lacked phosphorus, the authors reported that the growth of Chlorella sp. and Scenedesmus sp. was supported by the residual phosphorus in the inoculum. Additionally, being a mixotrophic strain, Chlorella sp. could utilize suitable organic compounds from the PW. When N and P were supplemented in the PW, the biomass density of Chlorella sp. increased by 32% after 15 days. The other strains lost the green pigments (i.e., chlorophylls) and their biomass productivity was decreased. Additionally, none of these strains could grow even after supplementing nutrients, which indicates the toxicity of the pre-treated PW. Chlorella sp. had the maximum biomass density on both occasions. In a similar study, Godfrey et al. reported that substitution of growth media with PW supplemented with N and P can be more economical owing to comparable lipid and biomass productivities at a lower cost and without the requirement of freshwater [47].
Therefore, the growth dynamics and cell density of algae can be significantly influenced by the PW inclusion, depending on the availability of nutrients such as N and P in the ultimate culture media. Moreover, some manipulation of these key nutrients can alter the metabolic and developmental pathways in algae. For instance, the addition of supplemental nutrients in all cases resulted in higher biomass densities and a longer exponential growth phase, as reported by Ranjbar et al. [39]. On the other hand, other research demonstrated a faster log phase and doubling time by increasing the PW supplementation. For example, Talebi et al. showed that biomass production and lipid content of D. salina increased by approximately 120% and 65%, respectively, compared to those in seawater (control), and after the addition of the PW [46]. As a result, approximately two to four times higher lipid productivity values were achieved using the cultures enriched by different ratios of seawater and PW (1:1 to 3:1). The group also reported that the algal cells grown in the 1: 1 medium reached the pre-stationary phase earlier than that in seawater, where the cells were still in their early stage of development. In addition, the biomass productivity (BP), lipid content (LC), and volumetric lipid productivity (LP) increased approximately 120%, 65%, and 263%, respectively, when compared with that of the seawater. Moreover, additional dilutions of the PW, i.e., 2:1 and 3:1 (seawater and PW), negatively affected the BP compared to the 1: 1 medium. The algal cells grown in the 1: 1 medium also grew larger (presumably due to moderate β-carotene accumulation) than those in the other medium. As the authors analyzed the bioremediation capability of D. salina, it turned out that the NO3 and PO4 reduced by ~66% and ~41%, respectively, which indicated that the algal cells efficiently took up the required nutrients (N and P) present in the PW. Overall, the group attributed these findings to the availability of required N and P in the 1: 1 medium, therefore, no further nutrient supplementation was necessary to improve biomass production and consequently the increase in BP and LP values.
Among the large-scale studies, recent research by Wood et al. showed that the Rotating Algal Biofilm Reactor (RABR) growth platform, coupled with utilizing PW as a growth medium could reduce the costs of large-scale phycocyanin production [65]. The authors used undiluted PW to grow cyanobacteria that they named Logan Lagoons Cyanobacteria Two (LLC2). Phycocyanin accumulated in the cyanobacterial biofilms reached the maximum value during the stationary period of LLC2 growth. However, the authors reported that phycocyanin productivity was lower in the undiluted PW than that of the prepared growth medium. Therefore, supplemental use of the prepared medium can be necessary to ensure nutrient availability.
2.1.5. Dilution of PW for Optimal Growth of Algae
The concentration PW in the medium can determine the growth and biomass productivity of algae. Many studies, as presented in Table 2, have used various dilutions of PW as standard media for the successful cultivation of algae. For example, Calderón-Delgado challenged C. vulgaris to 0%, 25%, 50%, 75%, and 100% of PW concentrations adding inorganic culture medium [61]. Their results showed continuous growth and cellular division with a constant rate until 72 h, before a high variation, when compared with the control between lower and higher concentrations, was observed. At 120 h, cell density of 50% PW and 75% PW was considerably lower than that of the control group. In contrast, experiments with 100% PW and 25% PW showed the highest cell density. The authors attributed this result, at the lowest concentration, to a hormetic effect, which is a phenomenon favored by a low-dose stimulation over any biological response [68,69]. The authors reported the treatments with 50% PW and 75% PW as self-limiting systems where the waste products, in the presence of toxic compounds, inhibited the growth by depositing and accumulating in the cellular membrane, reducing the exchange between the cell and the outside environment [70]. Nevertheless, it is well known that PW with a high nutrient concentration can induce eutrophication in an ecosystem through fast adaptation such as physiological acclimation, modification of genetic expression etc. [71]. Moreover, PW can transport bacteria capable of producing compounds that can be used by algae making a symbiotic relationship [72].
Based on the above literature survey and current discussion, four critical steps have been identified to promote the cultivation of algae in PW. These steps are presented in Figure 2. Typically, pre-treatments that remove organic constituents, and not the salinity, should be the most economical. Therefore, aligning the available salinity of the PW with the optimal salinity for a cultivated alga of interest can save considerable treatment costs for algal media preparation. Nevertheless, a clear understanding of PW chemistry is always critical to success in determining cultivation goals [40].
2.2. Algal-Based Biofuel Production Using PW
The complex chemical nature of PW is often found to create stress on the algae. For example, Neal et al. reported several conditions, such as bacterial contamination, precipitate formation, and the wrong type of nutrient that may cause the stress [32]. However, the lower growth may not be as important to overall biofuel production if higher lipid yields result from such PW-induced stress. Additionally, PW can be deficient in nitrogen, as reported by Ranjbar et al. [39], which is essential for increasing the algal growth. Therefore, the addition of supplemental nitrogen can be done at low concentrations to stimulate lipid accumulation in algae [39]. In addition, the dynamics of nutrient uptake and depletion of nitrogen can play a vital role in deciding the production of lipids. For instance, Ranjbar et al. inoculated D. salina in PW for 29 days and recorded a higher cellular lipid content in the presence of supplemental nutrients than that in their absence [39]. This finding contradicts the fact of stimulation in D. salina’s lipid production under nitrogen limiting conditions. However, the nitrogen analysis in the growth medium disclosed that the nitrogen in both cultures, with and without supplemental nutrients, was nearly exhausted at the beginning of the stationary phase after 22 and 15 days, respectively.
Godfrey successfully grew several strains of algae in PW, with a few strains producing significant amounts of non-polar or neutral lipids such as TAGs [47]. The work supplemented PW with sodium nitrate and potassium phosphate, since both N and P were much lower in the PW than the amount needed for growth. Among the strains tested, Amphora coffeaformis yielded high TAG content. In an effort to optimizing the amount of these nutrients for maximal growth and lipid production, N and P were added, ranging from no addition to that of the Broad Seawater (BS) recipe which contains 150 mg L−1 sodium nitrate and 250 mg L−1 potassium phosphate, respectively. The results from these experiments showed that the highest lipid production occurred at no addition of phosphate. In contrast, algal growth was highest at the maximum phosphate concentration. However, no significant difference was observed in biomass production at all other concentrations. Therefore, Godfrey concluded that the addition of phosphate stimulated no advantage in the biofuel objective and A. coffeaformis could produce sufficient biomass and lipids in the PW. In a further effort to optimize the nitrate concentration, sodium nitrate was supplemented at 0, 25, 75, 150, and 300 mg L-1. The highest neutral lipid formation was achieved when 75 mg L−1 sodium nitrate was supplemented, while the highest dry weight was achieved at 150 mg L−1 sodium nitrate. It was evidenced that adding < 75 mg L−1 nitrate was insufficient to trigger lipid formation. On the other hand, at 150 mg L−1 nitrate addition, biomass production was the highest and a reasonable concentration of neutral lipids was produced. A very similar result was obtained for Chaetoceros gracilis as Godfrey conducted a similar optimization experiment for C. gracilis. Therefore, Godfrey selected 150 mg L−1 nitrate concentration as the optimal amount to add to PW as growth medium.
Although the highest lipid production occurred with no addition of phosphate, the study further reported that phosphate addition had a distinct effect on A. coffeaformis and C. gracilis. Both TAG content and dry weight in the former were higher when no phosphate was added, whereas phosphate addition increased both lipid production and growth for the latter [47]. More interestingly, C. gracilis was observed to achieve higher optical densities in PW than BS media. Furthermore, the dry weight of biomass for any given phosphate concentration was higher than that in the earlier studies. These results indicate that PW can outperform BS in growth and lipid production for C. gracilis. Although phosphate addition can increase both lipid and growth productivities, enough growth can be achieved without phosphate addition, if production costs are required to be kept marginal [47].
Generally, PW is a matter of great concern due to the presence of high amounts of NaCl [5]. Moreover, the salt content in PW can often influence the biofuel productivity of algae, be that either freshwater or marine. For example, Scenedesmus obliquus, a freshwater green alga, showed an increase in lipid content after the addition of excess NaCl to some extent [73], however, this was negatively influenced by a further increase in NaCl level [74,75]. Therefore, while it is crucial to understand how salt stress can impact the lipid productivity in PW, the hypersaline salinity levels—in general—enhance algal lipid yields for the transesterification to biodiesel [41]. For example, a very recent study that reported the halotolerance of C. aponinum and P. kessleri in PW, indicated that lipid production by these algae varies with salinity. Although it is well known that lipid content increases with nutrient depletion, the post-nutrient-depletion lipid enrichment was displayed at ≥60 g TDS L−1 PW. Although, lipid fractions were greater in higher salinity conditions, such as 75 and 90 g TDS L−1, polyculture’s tolerance for high growth, at or near 60 g TDS L−1, needs to be considered. Nevertheless, polyculture cultivation in a hypersaline PW based media with depleted nutrients can be an efficient approach to augment the lipid content. The polyculture exhibited the highest lipid content after phosphorus or nitrogen depletion of the 60 g TDS L−1 PW media. The authors stepped forward and found that P. kessleri has a greater capacity to tolerate and utilize ammonium and the ability to outcompete C. aponinum, which was inhibited by higher ammonium and phosphate levels between day two and day seven of the experiment. Although the authors hypothesized that the higher alkalinity (bicarbonate concentration) in PW could shift the balance between P. kessleri and C. aponinum, further investigations in terms of fuel conversion yields of such polyculture systems are needed [41]. Additionally, the study showed that lower initial concentrations of ammonium and phosphate resulted in the highest growth rates, and the lipid productivity could be maximized by ensuring the limitation of only one of these nutrients after peak biomass density is achieved.
In a similar study by the same group, lipid production by D. tertiolecta displayed a diverse pattern in various salinity conditions [60]. In the 30 and 60 g TDS L−1 experiments, early lipid concentration and lipid content was high, then decreased, again increased after day 15 as the culture reached the stationary phase and then continued until the end of the experiment where it reached a maximum value of 0.070 g L−1. In the 120 and 210 g TDS L−1 experiments, lipid concentration did not exhibit any early bump, rather, it increased at a somewhat constant rate until the culture entered stationary phase on day 17. Although nutrient starvation of alga has been reported for inducing lipid production, there has been a lack of published results testing the effect for D. tertiolecta at salinities higher than seawater (about 35 g TDS L−1) [60]. In their 30 and 60 TDS L−1 experiments, Hopkins et al. showed that the total lipid content decreased shortly after nutrient depletion but increased later until reaching a maximum concentration on day 21. On the other hand, in the 120 and 210 g TDS L−1 experiments, the lipid content of biomass exhibited an increase after nitrate depletion but then either decreased or leveled off after reaching the maximum biomass concentration around day 16. In this context, it is likely that maintaining nutrient concentrations in the media could yield higher lipid productivity by maximizing biomass growth. In a similar study, Calderón-Delgado observed a decrease in total phenols concentrations and total hydrocarbons in C. vulgaris exposed to 75% PW and 100% PW [61].
2.3. Bioremediation of PW Using Algae
Remediation of PW containing heavy metals poses a great challenge, mainly because of the costly treatment methods [76]. Algal-based bioremediation of the PW is regarded as a promising method in terms of both efficiency and capital/operational costs [46]. The bioremediation of heavy metals is achieved through the binding of the metals to the organic ligands present in algal biomass representing the adsorption capacity of the culture [77,78]. It is worth noting that the metal removal potential of algal cells is directly related to the biomass concentration. Therefore, higher absorption capacities could be achieved by increasing biomass concentration [79,80]. This is attributed to the fact that more algal cells reveal more free-binding sites to absorb more metal ions. Various types of binding groups on the cell surface, such as amino, carbohydrate, carboxyl, hydroxyl, phosphoryl, sulphuryl etc. determine the final metal ions biosorption capacity of the cell [81,82]. Nevertheless, the number of sites on the algal cells, the chemical state of these sites and the accessibility of binding groups for metal ions altogether act as determining factors for the ultimate volume of absorbed heavy metals [83].
Generally, the metal absorption capacity of algae can vary depending on both the species and the strains. In their comprehensive study, Godfrey who utilized PW as a medium to grow various microalgae including C. gracilis, A. coffeiformis, P. tricornutum and several other strains isolated from the Great Salt Lake, reported that all organisms successfully reduced the concentration of heavy metals such as Ba, Ca, Fe, Mn, Si etc. and other elements that are potentially more harmful to the environment such as As, P and Sr. In fact, the diatoms A. coffeaformis and C. gracilis absorbed more of these elements from PW than the other algal strains and their growth reduced the concentrations of Ca, Mn and P in the PW that thereby met the water quality standards in the state of Utah, USA. In a recent study, Al-Ghouti et al. used filtered PW from a natural gas field in Qatar to grow five different chlorophytes (Dictyosphaerium sp., Scenedesmus sp., Neochloris sp., Chlorella sp. and Monoraphidium sp.) and investigated their capabilities to remove heavy metals (Table 2) [11]. The group reported achieving 100% removal efficiency of Al, Fe and Zn. Among the species, Dictyosphaerium sp. had better growth than the others and was able to recover more elements. It is worth noting that previous research showed that Dictyosphaerium sp. can grow in metal-rich water [84,85].
In addition, the initial concentration of metal ions in the PW can determine the final biosorption ability of algae [46]. While metal biosorption can initially increase with increasing metal concentration up to a certain level, however, any further increase in the concentration may deteriorate the adsorption capacity [46,86]. Ranjbar et al. inoculated D. salina in sterile PW and measured concentrations of 10 metals (Al, Ba, Cr, Cu, Fe, Mg, Mn, Ni, Sr, Zn) in the culture medium. After 29 days of incubation, seven of these metals were found to be reduced by a significant percentage upon the growth of D. salina cells [39]. In addition, Talebi et al. used the same algae and was able to reduce more than 84% of the Ni ions (5–100 mg L−1) from the PW in only six hours [46]. However, increasing the concentration led to a decreased bioremediation capacity of D. salina, observed as the researchers doubled the concentration of the Ni ions from 100 mg L−1 to 200 mg L−1. The active cells of D. salina absorbed free ions vigorously using their free binding sites that saturated as the ion concentration increased. The same study reported that D. salina removed 78% and 91% of free Zn ions at 2 to 5 mg L−1, respectively, during the six-hour cultivation period. However, higher initial Zn concentrations such as 10 and 20 mg L−1 negatively affected the Zn removal capacity of the algal cells as Zn harmfully affected the photosynthetic apparatus [87,88].
Therefore, the PW loading is crucial to alleviate the negative effects of some toxic components present in PW and to facilitate the bioremediation process by microalgae. Moreover, it has been reported that many microalgae are capable of decomposing organic compounds in the PW at appropriate loading. For instance, Ammar et al. who cultivated marine microalgae N. oculata and I. galbana in 10%, 25%, and 50% PW loading, reported that both the species showed significant chemical oxygen demand (COD) and oil removal efficiencies [45]. However, the group observed differences in the removal efficiencies that were based on the type of microalgae species and PW loadings. Pre-adaptive N. oculata showed better oil removal in low PW loading (10% and 25%), whereas I. galbana showed better oil removal in 50% PW loading. In fact, N. oculata and I. galbana reduced 66.5% and 68% of the oil, respectively, in the 50% PW medium in 21 days. However, in 10% PW, N. oculata and I. galbana were able to reduce 89% and 82% of the oil, respectively. As for the COD removal, N. oculata and I. galbana, respectively, removed 54%–90% and 56%–83% of COD. As carbon and the nutrients decreased after increasing the PW loading, microalgae were forced to consume organic compounds in the PW as the sole carbon source, especially when the pre-adaptation method was incorporated. Therefore, the COD removal efficiency decreased with increasing PW loading. This is caused by the high salinity of the PW which evidently reduced the biodegradation ability of the microalgae [89]. Nevertheless, it has been clearly observed that salt-adapted microalgae can resist high salinity and degrade the pollutants that are present in PW.
In their US patent, Mendes et al. evaluated the behavior of microalgae in PW containing high concentrations of ammonia [62]. This Brazilian research group prepared high-ammonia PW by deliberately adding ammonia in PW in a concentration that was two orders of magnitude higher than that is permitted by the Brazilian law for discharge into the environment. They inoculated two different species of microalgae, Phormidium sp. and Cyanobium sp. separately in a final medium that contained 95% (by volume) high-ammonia PW and 5% H2 culture medium. The group carried out bioassays for seven days and reported bioassimilation that removed more than 110% of ammonia than that in the control sample which contained no algae. In a similar experiment the authors inoculated Cyanobium sp. in high-phosphate PW and reported an increase in chlorophyll concentration and a decrease in phosphate concentration that, respectively, indicated the growth of the microalgae and the uptake of phosphate. Therefore, these findings indicate the enormous potential of microalgae for technological applications of this nature.
Among the large-scale experiments, Das et al. grew Chlorella sp. in 1 L photobioreactor (PBR) to study the contaminant removal efficiencies and reported that after 15 days of growth Chlorella sp. could remove 73% of the total organic carbon (TOC) and 92% of the total nitrogen (TN) from the pre-treated PW [4]. The TN removal rate was low for the first three days as the microbial nitrogen requirement in the lag phase was low. In contrast, the rate was higher for the next eight days until Chlorella sp. reached the stationary phase. Since the pre-treated PW was not sterilized before inoculating Chlorella sp., the degradation of the organics could have occurred by bacteria and made the nitrogen compounds available for microalgal consumption. Overall, Chlorella sp. and/or the consortium achieved a very high nitrogen removal efficiency. Nevertheless, since PW lacks N and P in most of the cases, Das et al. suggest adding these elements, as required, for maintaining high biomass density and consequently, a more efficient bioremediation process [4].
In addition to microalgae, aerobic bacteria can detoxify PW [90]. Therefore, microalgae-bacterium consortium can be considered as a viable treatment option when it comes to bioremediation. However, higher biomass density of microalgae-bacterium consortium is required for efficient detoxification [4]. In this context, Das et al. showed a symbiotic relationship between Chlorella sp. and aerobic bacteria that could have developed in the PW. The consortium successfully reduced the contaminants in addition to detoxifying the PW. As discussed previously in the current review, Eldorado Biofuels is growing a mix of freshwater algae (Scenedesmus and Tetracystis) referred to as Jalgae™, in a 10%–50% PW solution mixed with freshwater [32]. As such, a symbiotic relationship can reduce the cost of both the acquisition and disposal of PW.
2.4. Post-Processing of Algae and PW
It is evident that combining the algal-based treatment of PW for heavy metal and nutrients removal with biofuel and/or biomass production is a promising platform with great economic aspects [91]. However, biofuel and biomass should be considered as by-products and not as mainstream products. Therefore, both need to undergo post-processing for further utilization. Thus, the selection of the downstream processes can greatly influence the energy requirements, operating costs, economic viability, and efficiency of algal-based treatment systems [92].
In this context, Brasil et al. proposed a PW treatment facility with combined microalgae cultivation and processing to allow the opportunity for process integration with a reduction in logistics costs [3]. After cultivation, the algal biomass can be harvested and converted into biofuels and bioproducts. However, the actual scenario for microalgae biorefineries integration to the biofuels and petrochemical industries can pose some major challenges. Harvesting itself is generally a multi-stage process that is responsible for up to 30% of the total production costs [3]. It generally involves one or more steps of solid-liquid phase separation, such as flocculation, filtration, flotation, settling and/or centrifugation [93]. Although, centrifuges are a proven technology that have been used as a means for harvesting algae for many years, the high energy requirements and the costs to centrifuge large amounts of water reduce the economic viability of this process [92]. Therefore, low-cost harvesting techniques with high processivity is recommended to dewater algal biomass before biomass conversion and/or lipid extraction. Harvesting methods based on the use of chemical flocculants and dissolved air flotation are low-cost options that have been successfully applied either to wastewater treatment or to cultured microalgae [94,95]. Especially promising are methods based on the use of non-toxic flocculants such as mixed calcium and magnesium hydroxides [96]. In terms of cost-effectiveness, methods based on membrane filtration, ultrasound, and electrocoagulation harvesting are considered to have high potential. In fact, electrocoagulation is already an established technology with existing applications to remove charged ionic contaminants from wastewater [92,97]. Optimistically, recent advances in microalgae large-scale production can provide technologies necessary to integrate such low-cost methods in the current PW treatment system.
The harvested algal biomass can be converted into fuel through various techniques such as anaerobic digestion [98], bioethanol fermentation [99], transesterification [100], pyrolysis [101], and hydrothermal liquefaction [102]. However, the conversion of whole wet algal biomass into oil through hydrothermal liquefaction and catalytic hydrothermal gasification (HTL–CHG) might be the most cost-effective downstream processing route, as reported by several studies in recent times [92,103,104]. For example, Richardson and coworkers reported 69% higher revenues generated using HTL–CHG than that of using wet solvent extraction [92]. Moreover, the combination of electrocoagulation harvesting and HTL–CHG has been reported to lead to a 90% lower lipid total cost compared to centrifugation followed by wet solvent extraction [92,103].
In addition to the traditional harvesting and conversion techniques, the development of novel methods based on experimental outcomes can often lead to cost-effective post-processing. For example, a recent study by Wood et al. showed that the purity of phycocyanin (measured spectrophotometrically as the ratio of absorbance values at 620 nm and 280 nm) increased as the cellular phycocyanin content increased when they inoculated LLC2 cyanobacteria in a RABR growth platform that utilized PW as a growth medium [65]. Additionally, phycocyanin accumulated in the cyanobacterial biofilms reached the maximum value during the stationary period of the LLC2’s growth. Therefore, harvesting the biofilm produced in RABR during the stationary period can lead to a lower downstream phycocyanin purification processing cost. In a similar study, Godfrey reported that the combined production of phycocyanobilin and biodiesel using cyanobacteria Synechocystis sp. could be highly commercially feasible owing to the extreme availability of the PW [47]. Moreover, since the phycocyanobilin extraction utilizes water solvents and biodiesel extraction utilizes organic solvents, the two extraction processes can be performed on the same biomass material.
Generally, the higher the biomass concentration in the PW, the lower will be the cost of biomass harvesting [105]. However, since N and P are often supplemented in PW there may be additional cost to this [4]. Hence, recovering a fraction of the biomass bound N and P as byproducts and adding in the subsequent batch of the process can optimize the cost [106]. An additional step is separating the contaminated biomass from the purified PW by separation processes, such as floatation after the algae have concentrated metal compounds [62]. Thus, supplementing the PW with N and P can improve the overall process economics. Alternatively, the biomass residues can be used instead of fertilizer and digested for biogas, which can be used as a CO2 and energy source [107]. As for large-scale development, the usability of algae residues on site should be taken into consideration to determine the economic viability of a PW based algal pond [49].
In addition, the commercialization of the PW based algal cultivation system must address the downstream processing of saline water disposal [47]. Although some marine microalgal species such as N. oculata and I. galbana have an excellent ability to endure extreme conditions such as salinity, the bioremediation of PW using microalgae should not be considered realistic until efforts to remove salinity are made, as commented recently by Godfrey [47]. The author reported that the TDS, sodium, and chlorine concentrations were hardly affected by microalgal growth and remained above the water quality standards. It is the high salinity of PW that brings the TDS orders of magnitude higher than permitted at any location, and microalgae have no promise to uptake large amounts of salt. Additionally, while microalgae successfully removed nutrients like N and P, and reduced Ca and Mn to below acceptable concentration ranges, most other elements were in an acceptable range prior to the cultivation [47]. Therefore, the disposal of saline water should be taken seriously in the post-processing phase.
3. The Large-Scale Development of Algae-Based PW System
3.1. Factors Affecting the Large-Scale Development
The use of ponds to cultivate algae for large-scale production of biofuels and biomass is not a new idea, but the use of PW in algal ponds has hardly been studied. Among the few large-scale studies, Graham et al. reported some key factors that are relevant to algae cultivation using PW in ponds [40]. The factors include dry climate, availability of large and inexpensive areas, and solar irradiance. These factors can be considered major and are presented in Figure 3. There have been some studies that considered these factors hence can provide further insight. For example, Winckelmann et al. reported the growth of indigenous algae in open ponds in an arid region of southwestern United States where freshwater sources are scarce. It is worth mentioning that the use of PW can offset the demand for freshwater in algae cultivation. In this context, marine algae species such as Nannochloropsis sp. can be exploited to treat PW in addition to biofuel production [32,50]. Neal et al. reported a 44.38% cost saving by using nutrients present in the PW over fresh water to mix the f/2-Si media. However, there are some challenges associated with open outdoor ponds, especially regarding solar irradiance.
In addition to the factors discussed above, it is also critical to ensure the availability of PW for large-scale development of PW-based algal ponds. Therefore, regions that generate large quantities of PW should be considered [60]. For example, the Permian Basin of southeast New Mexico can be a feasible option. Especially, the utilization of PW in these areas for algae production would not conflict with scarce freshwater for agricultural and municipal use. The average TDS of PW in the Permian Basin is 154–225 g TDS L−1 [108]. Since the salinity of algae media used in outdoor cultivation would increase through evaporation, therefore, halotolerant species that can thrive in such a challenging environment would be ideal to efficiently utilize residual nutrients [109]. Typically, evaporation is the primary way of losing water in algae cultivation while the secondary losses occur during harvesting, as water is removed with the algae and media [110,111]. Therefore, dilution of saline media with less saline brackish sources can be done to maintain correct media salinity levels under the correct conditions. PW in several basins also contains dissolved inorganic carbon, mostly in the form of bicarbonate, at levels higher than seawater. The salinity of PW worldwide differs from near fresh (1000 mg TDS L−1) to highly saline (>200,000 mg TDS L−1) depending on the source formation and region [40]. Salinities near seawater concentrations or lower are the best for algae growth. However, PW volume and location data are needed to evaluate the feasibility of where algae cultivation could be co-located [40]. This may lead to higher biomass productivity, which can be more pronounced in hypersaline conditions where inorganic carbon is less available than in PW-based media [60]. Moreover, it has been reported that algal species such as D. tertiolecta increased lipid productivity when cultivated using 100% CO2 along with NaOH [112]. Therefore, a bicarbonate-rich medium can increase biomass productivity and biofuel production in hypersaline PW which would improve the overall economics of PW-based algae cultivation systems. However, any bicarbonate induced lipid enrichment is unlikely to occur under nutrient-depleted conditions. Hence, increasing carbon availability (through carbon dioxide aeration or bicarbonate addition while maintaining a seawater pH) can be a better strategy than nutrient starvation to increase lipid productivity in D. tertiolecta using PW. In addition, further research needs to be conducted with other halophiles under Dunaliella genus such as D. salina that are proposed to capture inorganic carbon at high ionic strength.
In a similar context, some other potential areas in the United States include the southeastern portion of New Mexico and parts of west Texas (Permian Basin), where solar irradiance and land are available and where freshwater availability is low [40]. In addition, the availability of PW in all these regions is high since oil and gas extraction is done both by conventional and unconventional extraction methods. On the other hand, northeastern New Mexico and southwestern Colorado have higher elevations and less availability of flat land for cultivation. However, lack of oil and gas production in the west of the Permian Basin region requires additional facility to avail PW for algae production in South–central New Mexico. Therefore, pipelines can be used to transfer PW from the Permian Basin to these areas for large facilities, whereas truck transportation can be a viable option for smaller facilities [40]. Iran, for instance, among other oil-producing countries, generates a huge amount of PW. This west Asian country has a long coastline and numerous sunny days. A recent study estimated that by using the available technology/experts on algal-based PW treatment, the country can produce 794.75 × 103 tons of biodiesel monthly that could meet ~26.5% of its diesel demand [46].
3.2. Challenges and Future Prospectives
The productivity of large-scale platforms such as ponds can often be challenged by non-axenic culture conditions. For example, Winckelmann et al. reported weed algae contamination as they grew Scenedesmus sp. MKB in PW in their open pond system [49]. In addition, the physiological stress caused by the compounds present in PW hinders the biomass production [42]. Factors such as the presence of toxic metals, excessive nutrients, and other organic compounds contribute to this hindrance. Generally, microorganisms such as algae have passed through a process of natural selection and are naturally adapted to the hostile environment offered by PW [62]. However, physiological adjustments are often necessary for a strain of alga to adapt to new growing conditions as the characteristics of PW vary.
For example, Mendes et al. reported the growth of genera Phormidium sp. and Cyanobium sp. in PW which surprisingly had comparable growth to those obtained in a conventional culture medium, indicating that they had fully adapted to the medium [62]. Therefore, successive transfers of algae in the PW, and several generations after the initial inoculums can lead to efficient acclimatization of the algal cells [113]. One of the most comprehensive studies to date was done as a thesis by Godfrey, who utilized PW as a medium to grow various microalgae including C. gracilis, A. coffeiformis, P. tricornutum, and several other strains isolated from Great Salt Lake (Table 2). N and P were supplemented in the PW media along with CO2 bubbling. The C. gracilis strain, initially tested in shaker flasks and tube cultures, was later scaled-up to 5 L indoor and 50 L outdoor bag reactors [47]. In each case, algae growth and biofuel production using PW media were reported to be equal to, or higher than, laboratory media. Additionally, C. gracilis was cultivated in a 220 L raceway. Although the biomass productivity was lower than that in closed tube systems, but is quite satisfactory for an open raceway that lacks sterility, as opposed to the laboratory. Godfrey, therefore, concluded that algal growth could indeed be done in larger-scale systems using PW [47].
Although the sterility is compromised in open pond systems, the high salinity, free ammonia, and pH of PW would be highly advantageous in lowering predatory organisms such as rotifers in outdoor ponds and would allow algae to thrive in ammonium-rich PW [60]. For instance, PW with high ammonium and phosphate has been reported to lead to conditions that favored the development of large-scale production of biofuels using oleaginous algal species such as P. Kessleri [41]. Additionally, some microalgae can develop strategies to control and prevent pollution in the unsterile condition. For instance, N. oculata and I. galbana have been reported to follow such a strategy known as allelopathy where they release biochemicals, known as allelopathic compounds, that have a toxic effect on other organisms such as the anaerobic bacteria [114]. Non-axenic harsh conditions, such as high salinity and low nutrients or unbalanced N:P, help their growth especially in the absence of other microorganisms. This fact has a close connection with competition for limiting nutrient resources, as reported by Graneli [115]. Therefore, further understanding of this strategy can be an interesting and promising way to develop algae-based PW systems [116].
In the light of current discussion, Table 3 presents an exclusive list of algal species that are tolerant of a relatively-wide range of salinity. Although these species have been extensively reported to thrive in brackish, saline and hypersaline conditions, most of them are not yet explored in the context of PW systems. These species belong to various phyla including Bacillariophyta, Cyanobacteria, Euglenozoa, Haptophyta, Heterokonta and Ochrophyta, however, most of them are primarily associated with marine habitats and many of them are known as halophiles. Therefore, they are high-potential candidates for being deployed in PW of various salinity ranges.
In the context of the large-scale production of biofuels there are some financial challenges due to the high cost of nutrients such as N and P, used in culture media, that are essential to produce algal biomass [3]. A couple of Brazilian companies, named Petrobras and Embrapa, are leading programs to characterize and domesticate productive native algal strains. While Petrobras primarily focuses on biodiesel production using marine microalgae, Embrapa aims at characterizing continental microalgae and establishing a long-term research program for bioproducts and biofuel production. Since 2012, Petrobras has been making investments focused on microalgal-based treatment of PW and CO2 emissions capture with its pre-commercial oil exploration operations in the northeastern coastal region of Brazil [3].
In a recent study, Winckelmann et al. cultivated indigenous isolate Scenedesmus sp. MKB in open ponds (4 m long, 0.8 m wide, and 0.38 m deep) on non-arable land in the Al-Wusta region of the Sultanate of Oman [49]. Although evaporated water was replaced with fresh treated PW, the changes in salinity from this process were not reported. Therefore, the concerns regarding salt stress due to evaporation remained unwarranted [40,49]. The group used pre-cleaned PW as a growth medium and conducted experiments in 600 L volume. Although Scenedesmus sp. successfully grew in PW ponds, the authors declared that an economic feasibility was not possible. However, they indicated that the nutrient costs, nutrient recycling, the usability of algae residues on-site, the work hours etc., need to be considered to determine the economic viability. Nevertheless, the study revealed that effective salt management could be achieved from fresh treated and even saline PW used as media makeup, even at the highest evaporation rates, for halotolerant algae.
Several researchers including Park and coworkers recently reported that coupled microalgae cultivation and wastewater bioremediation could (i) eliminate the cost of nutrient and freshwater implementation; (ii) minimize unit cost energy by 20%–25%; (iii) meet capital costs, operation and maintenance costs [144,145,146]. Additionally, a symbiotic relationship between algae can also help reduce the cost of both the acquisition and disposal of PW.
Although the algae-integrated biorefineries are promising, the economic viability of large-scale algal biomass cultivation is yet to be achieved. To date, the costs for microalgae biofuel production are still at least two-fold higher than its fossil-based counterparts [3]. Moreover, low-value products such as bulk chemicals and biomaterials have not yet achieved economic viability. Therefore, continued improvements in the areas of algal cultivation, harvesting and processing are needed. In addition, the feasibility of using PW as a potential source of cultivation media should be evaluated with regards to the current biorefinery combinations, environmental risks, and resources management. Moreover, region-specific studies need to be conducted to evaluate the effects of different climatic conditions.
4. Conclusions
Treated PW can mitigate the scarcity of water and can enhance oil and gas recovery from oil and gas fields. The algal approach can remediate PW of its toxic contaminants and provide biofuels, nutrients supplements, and bioactive compounds upon further processing. Moreover, PW offers some key advantages to water availability and culture optimization. In light of these facts, the present review has extensively described the potential of cultivating algae using PW as a replacement for micronutrients and freshwater. Although the available studies show that salinity and nutrients in PW support algal growth, pre-treatment and dilution of PW as well as the addition of nutrients are often required. These critical steps, as discussed in detail in the current study, are crucial to promote the cultivation of algae in PW. Additionally, our work emphasizes the cultivation of halotolerant and marine algal species in PW systems. Nevertheless, there is much to discover and investigate in order to address some significant challenges, such as the upstream and downstream processing of both algae and PW. For example, additional studies should be conducted to examine the performance of pre-adapted algae in PW in terms of fatty acid profile, lipid production, and biomass generation. Since such studies are clearly important, a conclusive statement on the feasibility of a large-scale algal-based PW treatment is not possible at this stage. Nevertheless, we conclude that algae have the potential to be involved in the biological segment of PW treatment and, therefore, demand further research. The successful development of this promising field would not only offer PW disposal for oil and gas operators, but also create a considerable opportunity for algae growers to save about half of the fertilizer costs.
Author Contributions
Conceptualization, A.R. and T.S.; methodology, A.R.; software, A.R. and S.P.; validation, A.R., S.A. and T.S.; formal analysis, A.R.; investigation, T.S.; resources, A.R. and T.S.; data curation, A.R. and S.A.; writing—original draft preparation, A.R and S.A.; writing—review and editing, A.R., S.A., T.N., S.P. and T.S.; visualization, A.R.; supervision, T.S. and T.N.; project administration, T.S.; funding acquisition, T.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Center for Midstream Management and Science (CMMS)—Lamar University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bakke, T.; Klungsøyr, J.; Sanni, S. Environmental impacts of produced water and drilling waste discharges from the Norwegian offshore petroleum industry. Mar. Environ. Res. 2013, 92, 154–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igunnu, E.T.; Chen, G.Z. Produced water treatment technologies. Int. J. Low Carbon Technol. 2014, 9, 157–177. [Google Scholar] [CrossRef] [Green Version]
- Brasil, B.; Silva, F.C.P.; Siqueira, F.G. Microalgae biorefineries: The Brazilian scenario in perspective. New BioTechnol. 2017, 39, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; AbdulQuadir, M.; Thaher, M.; Khan, S.; Chaudhary, A.K.; Alghasal, G.; Al-Jabri, H.M.S. Microalgal bioremediation of petroleum-derived low salinity and low pH produced water. J. Appl. Phycol. 2019, 31, 435–444. [Google Scholar] [CrossRef]
- Campos, J.C.; Borges, R.M.H.; Filho, A.O.; Nobrega, R.; Sant’Anna, G.L., Jr. Oilfield wastewater treatment by combined microfiltration and biological processes. Water Res. 2002, 36, 95–104. [Google Scholar] [CrossRef]
- Fakhru’l-Razi, A.; Pendashteh, A.; Abdullah, L.C.; Biak, D.R.A.; Madaeni, S.S.; Abidin, Z.Z. Review of technologies for oil and gas produced water treatment. J. Hazard Mater. 2009, 170, 530–551. [Google Scholar] [CrossRef]
- Chen, L.; Xu, Q.; Gossage, J.L.; Lou, H.H. Simulation and economic evaluation of a coupled thermal vapor compression desalination process for produced water management. J. Nat. Gas Sci. Eng. 2016, 36, 442–453. [Google Scholar] [CrossRef]
- Bayati, F.; Shayegan, J.; Noorjahan, A. Treatment of oilfield produced water by dissolved air precipitation/solvent sublation. J. Petrol Sci. Eng. 2011, 80, 26–31. [Google Scholar] [CrossRef]
- Fathy, M.; El-Sayed, M.; Ramzi, M.; Abdelraheem, O.H. Adsorption separation of condensate oil from produced water using ACTF prepared of oil palm leaves by batch and fixed bed techniques. Egypt J. Petrol 2018, 27, 319–326. [Google Scholar] [CrossRef]
- Jiang, Q.; Rentschler, J.; Perrone, R.; Liu, K. Application of ceramic membrane and ion-exchange for the treatment of the flowback water from Marcellus shale gas production. J. Membrane Sci. 2013, 431, 55–61. [Google Scholar] [CrossRef]
- Al-Ghouti, M.A.; Al-Kaabi, M.A.; Ashfaq, M.Y.; Da’na, D.A. Produced water characteristics, treatment and reuse: A review. J. Water Process. Eng. 2019, 28, 222–239. [Google Scholar] [CrossRef]
- Lu, M.; Zhang, Z.; Yu, W.; Zhu, W. Biological treatment of oilfield-produced water: A field pilot study. Int. Biodeter. Biodegr. 2009, 63, 316–321. [Google Scholar] [CrossRef]
- Chavan, A.; Mukherji, S. Treatment of hydrocarbon-rich wastewater using oil degrading bacteria and phototrophic microorganisms in rotating biological contactor: Effect of N:P ratio. J. Hazard Mater. 2008, 154, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Selvaratnam, T.; Pegallapati, A.K.; Montelya, F.; Rodriguez, G.; Nirmalakhandan, N.; Van Voorhies, W.; Lammers, P.J. Evaluation of a thermo-tolerant acidophilic alga, Galdieria sulphuraria, for nutrient removal from urban wastewaters. Bioresour. Technol. 2014, 156, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Luo, S.; Fan, X.; Yang, Z.; Guo, R. Biomass and lipid production of marine microalgae using municipal wastewater and high concentration of CO2. Appl. Energy 2011, 88, 3336–3341. [Google Scholar] [CrossRef]
- Henkanatte-Gedera, S.M.; Selvaratnam, T.; Caskan, N.; Nirmalakhandan, N.; Van Voorhies, W.; Lammers, P.J. Algal-based, single-step treatment of urban wastewaters. Bioresour. Technol. 2015, 189, 273–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, T.; Rahman, A.; Pan, S.; Dixon, K.; Petri, B.; Selvaratnam, T. A review of landfill leachate treatment by microalgae: Current status and future directions. Processes 2020, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Cai, T.; Ge, X.; Park, S.Y.; Li, Y. Comparison of Synechocystis sp. PCC6803 and Nannochloropsis salina for lipid production using artificial seawater and nutrients from anaerobic digestion effluent. Bioresour. Technol. 2013, 144, 255–260. [Google Scholar] [CrossRef]
- Hidaka, T.; Inoue, K.; Suzuki, Y.; Tsumori, J. Growth and anaerobic digestion characteristics of microalgae cultivated using various types of sewage. Bioresour. Technol. 2014, 170, 83–89. [Google Scholar] [CrossRef]
- Rahman, A.; Kumar, S.; Bafana, A.; Lin, J.; Dahoumane, S.A.; Jeffryes, C. A mechanistic view of the light-induced synthesis of silver nanoparticles using extracellular polymeric substances of Chlamydomonas reinhardtii. Molecules 2019, 24, 3506. [Google Scholar] [CrossRef] [Green Version]
- Klimmek, S.; Stan, H.-J.; Wilke, A.; Bunke, G.; Buchholz, R. Comparative analysis of the biosorption of cadmium, lead, nickel, and zinc by algae. Environ. Sci. Technol. 2001, 35, 4283–4288. [Google Scholar] [CrossRef] [PubMed]
- Krejci, M.R.; Finney, L.; Vogt, S.; Joester, D. Selective sequestration of strontium in desmid green algae by biogenic co-precipitation with barite. Chemsuschem 2011, 4, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.K.; Gaur, J.P. Use of algae for removing heavy metal ions from wastewater: Progress and prospects. Cr. Rev. Biotechnol. 2005, 25, 113–152. [Google Scholar] [CrossRef]
- Rahman, A.; Lin, J.; Jaramillo, F.E.; Bazylinski, D.A.; Jeffryes, C.; Dahoumane, S.A. In vivo biosynthesis of inorganic nanomaterials using eukaryotes—A Review. Molecules 2020, 25, 3246. [Google Scholar] [CrossRef]
- Mishra, A.; Jha, B. Isolation and characterization of extracellular polymeric substances from micro-algae Dunaliella salina under salt stress. Bioresour. Technol. 2009, 100, 3382–3386. [Google Scholar] [CrossRef]
- Selvaratnam, T.; Pegallapati, A.; Montelya, F.; Rodriguez, G.; Nirmalakhandan, N.; Lammers, P.J.; van Voorhies, W. Feasibility of algal systems for sustainable wastewater treatment. Renew. Energy 2015, 82, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Lammers, P.J.; Huesemann, M.; Boeing, W.; Anderson, D.B.; Arnold, R.G.; Bai, X.; Bhole, M.; Brhanavan, Y.; Brown, L.; Brown, J.; et al. Review of the cultivation program within the National Alliance for Advanced Biofuels and Bioproducts. Algal. Res. 2017, 22, 166–186. [Google Scholar] [CrossRef]
- Harkness, J.S.; Dwyer, G.S.; Warner, N.R.; Parker, K.M.; Mitch, W.A.; Vengosh, A. Iodide, bromide, and ammonium in hydraulic fracturing and oil and gas wastewaters: Environmental implications. Environ. Sci. Technol. 2015, 49, 1955–1963. [Google Scholar] [CrossRef] [PubMed]
- Fakhru’l-Razi, A.; Pendashteh, A.; Abidin, Z.Z.; Abdullah, L.C.; Biak, D.R.A.; Madaeni, S.S. Application of membrane-coupled sequencing batch reactor for oilfield produced water recycle and beneficial re-use. Bioresour. Technol. 2010, 101, 6942–6949. [Google Scholar] [CrossRef]
- Riley, S.M.; Oliveira, J.M.S.; Regnery, J.; Cath, T.Y. Hybrid membrane bio-systems for sustainable treatment of oil and gas produced water and fracturing flowback water. Sep. Purif. Technol. 2016, 171, 297–311. [Google Scholar] [CrossRef] [Green Version]
- Pendashteh, A.R.; Abdullah, L.C.; Fakhru’l-Razi, A.; Madaeni, S.S.; Zainal Abidin, Z.; Awang Biak, D.R. Evaluation of membrane bioreactor for hypersaline oily wastewater treatment. Process. Saf. Environ. 2012, 90, 45–55. [Google Scholar] [CrossRef]
- Neal, J.N.; Sullivan, E.J.; Dean, C.A.; Steichen, S.A. Recycling Produced Water for Algal Cultivation for Biofuels; Los Alamos National Lab. (LANL): Los Alamos, NM, USA, 2012. [Google Scholar]
- Markou, G.; Georgakakis, D. Cultivation of filamentous cyanobacteria (blue-green algae) in agro-industrial wastes and wastewaters: A review. Appl. Energy 2011, 88, 3389–3401. [Google Scholar] [CrossRef]
- Eriksen, N.T. Production of phycocyanin—A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sekar, S.; Chandramohan, M. Phycobiliproteins as a commodity: Trends in applied research, patents and commercialization. J. Appl. Phycol. 2008, 20, 113–136. [Google Scholar] [CrossRef]
- Chu, W.-L. Biotechnological applications of microalgae. Int. J. Sci. Med. Edu. 2012, 6, S24–S37. [Google Scholar]
- Reddy, H.K.; Muppaneni, T.; Ponnusamy, S.; Sudasinghe, N.; Pegallapati, A.; Selvaratnam, T.; Seger, M.; Dungan, B.; Nirmalakhandan, N.; Schaub, T.; et al. Temperature effect on hydrothermal liquefaction of Nannochloropsis gaditana and Chlorella sp. Appl. Energy 2016, 165, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Selvaratnam, T.; Reddy, H.; Muppaneni, T.; Holguin, F.O.; Nirmalakhandan, N.; Lammers, P.J.; Deng, S. Optimizing energy yields from nutrient recycling using sequential hydrothermal liquefaction with Galdieria sulphuraria. Algal. Res. 2015, 12, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Ranjbar, S.; Quaranta, J.D.; Tehrani, R.; Van Aken, B. Algae-Based Treatment of Hydraulic Fracturing Produced Water: Metal Removal and Biodiesel Production by the Halophilic Microalgae Dunaliella salina. In Bioremediation and Sustainable Environmental Technologies. In Proceedings of the Third International Symposium on Bioremediation and Sustainable Environmental Technologies, Miami, FL, USA, 18–21 May 2015. [Google Scholar]
- Sullivan Graham, E.J.; Dean, C.A.; Yoshida, T.M.; Twary, S.N.; Teshima, M.; Alvarez, M.A.; Zidenga, T.; Heikoop, J.M.; Perkins, G.B.; Rahn, T.A.; et al. Oil and gas produced water as a growth medium for microalgae cultivation: A review and feasibility analysis. Algal. Res. 2017, 24, 492–504. [Google Scholar] [CrossRef]
- Hopkins, T.C.; Graham, E.J.S.; Schwilling, J.; Ingram, S.; Gómez, S.M.; Schuler, A.J. Effects of salinity and nitrogen source on growth and lipid production for a wild algal polyculture in produced water media. Algal. Res. 2019, 38, 101406. [Google Scholar] [CrossRef]
- Arriada, A.A.; Abreu, P.C. Nannochloropsis oculata growth in produced water: An alternative for massive microalgae biomass production. Braz. J. Petrol Gas 2014, 8, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Almaraz, N.; Regnery, J.; Vanzin, G.F.; Riley, S.M.; Ahoor, D.C.; Cath, T.Y. Emergence and fate of volatile iodinated organic compounds during biological treatment of oil and gas produced water. Sci. Total Environ. 2020, 699, 134202. [Google Scholar] [CrossRef]
- Nie, H.; Nie, M.; Diwu, Z.; Wang, L.; Yan, H.; Lin, Y.; Zhang, B.; Wang, Y. Biological treatment of high salinity and low pH produced water in oilfield with immobilized cells of P. aeruginosa NY3 in a pilot-scale. J. Hazard Mater. 2020, 381, 121232. [Google Scholar] [CrossRef] [PubMed]
- Ammar, S.H.; Khadim, H.J.; Mohamed, A.I. Cultivation of Nannochloropsis oculata and Isochrysis galbana microalgae in produced water for bioremediation and biomass production. Environ. Technol. Innov. 2018, 10, 132–142. [Google Scholar] [CrossRef]
- Talebi, A.F.; Dastgheib, S.M.M.; Tirandaz, H.; Ghafari, A.; Alaie, E.; Tabatabaei, M. Enhanced algal-based treatment of petroleum produced water and biodiesel production. RSC Adv. 2016, 6, 47001–47009. [Google Scholar] [CrossRef]
- Godfrey, V. Production of Biodiesel from Oleaginous Organisms Using Underutilized Wastewaters. Master’s Thesis, Utah State University, Logan, UT, USA, 2012. [Google Scholar]
- “Production of Biodiesel from Oleaginous Organisms Using Underutilized” by Valerie Godfrey. Available online: https://digitalcommons.usu.edu/etd/1337/ (accessed on 23 July 2020).
- Winckelmann, D.; Bleeke, F.; Thomas, B.; Elle, C.; Klöck, G. Open pond cultures of indigenous algae grown on non-arable land in an arid desert using wastewater. Int. Aquat. Res. 2015, 7, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Badrinarayanan, I. Using Produced Water to Grow Microalgae; Russel School of Chemical Engineering the University of Tulsa: Tulsa, OK, USA, 2017; p. 20. [Google Scholar]
- Johnson, R.J.; Jurawan, I.; Frenzel, M.; Price, A.C. The identification and mechanism of a Scenedesmus spp. causing bio-fouling of an oil field produced water treatment plant. Int. Biodeter. Biodegr. 2016, 108, 207–213. [Google Scholar] [CrossRef]
- Peng, J.; Yin, K.; Yuan, J.-P.; Cao, G.-X.; Xue, M.; Wu, C.-F.; Wang, J.-H. Characterization of a newly isolated green microalga Scenedesmus sp. as a potential source of biodiesel. Afr. J. Biotechnol. 2012, 11, 16083–16094. [Google Scholar]
- Büdel, B.; Darienko, T.; Deutschewitz, K.; Dojani, S.; Friedl, T.; Mohr, K.I.; Salisch, M.; Reisser, W.; Weber, B. Southern African biological soil crusts are ubiquitous and highly diverse in drylands, being restricted by rainfall frequency. Microbial. Ecol. 2009, 57, 229–247. [Google Scholar] [CrossRef]
- Eliaš, M.; Němcová, Y.; Škaloud, P.; Neustupa, J.; Kaufnerova, V.; Šejnohová, L. Hylodesmus singaporensis gen. et sp. nov., a new autosporic subaerial green alga (Scenedesmaceae, Chlorophyta) from Singapore. Int. J. Syst. Evol. Micr. 2010, 60, 1224–1235. [Google Scholar] [CrossRef] [Green Version]
- Lewis, L.A.; Flechtner, V.R. Green algae (Chlorophyta) of desert microbiotic crusts: Diversity of North American taxa. Taxon 2002, 51, 443–451. [Google Scholar] [CrossRef]
- Lewis, L.A.; Flechtner, V.R. Cryptic species of Scenedesmus (chlorophyta) from desert soil communities of Western North America. J. Phycol. 2004, 40, 1127–1137. [Google Scholar] [CrossRef]
- Bartley, M.L.; Boeing, W.J.; Corcoran, A.A.; Holguin, F.O.; Schaub, T. Effects of salinity on growth and lipid accumulation of biofuel microalga Nannochloropsis salina and invading organisms. Biomass Bioenergy 2013, 54, 83–88. [Google Scholar] [CrossRef]
- Racharaks, R.; Ge, X.; Li, Y. Cultivation of marine microalgae using shale gas flowback water and anaerobic digestion effluent as the cultivation medium. Bioresour. Technol. 2015, 191, 146–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aravinthan, V.; Harrington, D. Coal seam gas water as a medium to grow Dunalliella tertiolecta microalgae for lipid extraction. Desalin. Water Treat 2014, 52, 947–958. [Google Scholar] [CrossRef]
- Hopkins, T.C.; Graham, E.J.S.; Schuler, A.J. Biomass and lipid productivity of Dunaliella tertiolecta in a produced water-based medium over a range of salinities. J. Appl. Phycol. 2019, 31, 3349–3358. [Google Scholar] [CrossRef]
- Calderón-Delgado, I.C.; Mora-Solarte, D.A.; Velasco-Santamaría, Y.M. Physiological and enzymatic responses of Chlorella vulgaris exposed to produced water and its potential for bioremediation. Environ. Monit. Assess 2019, 191, 399. [Google Scholar] [CrossRef]
- Mendes, L.B.B.; Cunha, P.C.R.; D’oca, M.G.M.; Abreu, P.C.; Primel, E.G. Method for Removing Pollutants from Produced Water. U.S. Patent US 7,955,505, 7 June 2011. [Google Scholar]
- Mohd Abdul Hakim, M.A.Q. A Potential Application of Microalgae in Produced Water Treatment. Master’s Thesis, Qatar University, Doha, Qatar, 2016. [Google Scholar]
- Andrade, B.G.; Andrade, V.T.; Costa, B.R.; Campos, J.C.; Dezotti, M. Distillation of oil field produced water for reuse on irrigation water: Evaluation of pollutants removal and ecotoxicity. J. Water Reuse Desal. 2011, 1, 224–236. [Google Scholar] [CrossRef]
- Wood, J.L.; Miller, C.D.; Sims, R.C.; Takemoto, J.Y. Biomass and phycocyanin production from cyanobacteria dominated biofilm reactors cultured using oilfield and natural gas extraction produced water. Algal Res. 2015, 11, 165–168. [Google Scholar] [CrossRef]
- Vento, S.D.; Dachs, J. Prediction of uptake dynamics of persistent organic pollutants by bacteria and phytoplankton. Environ. Toxicol. Chem. 2002, 21, 2099–2107. [Google Scholar] [CrossRef]
- Fan, C.-W.; Reinfelder, J.R. Phenanthrene accumulation kinetics in marine diatoms. Environ. Sci. Technol. 2003, 37, 3405–3412. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Blain, R.B. Hormesis and plant biology. Environ. Pollut. 2009, 157, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Spoljaric, D.; Cipak, A.; Horvatic, J.; Andrisic, L.; Waeg, G.; Zarkovic, N.; Jaganjac, M. Endogenous 4-hydroxy-2-nonenal in microalga Chlorella kessleri acts as a bioactive indicator of pollution with common herbicides and growth regulating factor of hormesis. Aquat. Toxicol. 2011, 105, 552–558. [Google Scholar] [CrossRef]
- Franqueira, D.; Orosa, M.; Torres, E.; Herrero, C.; Cid, A. Potential use of flow cytometry in toxicity studies with microalgae. Sci. Total Environ. 2000, 247, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Witt, G. Occurrence and transport of polycyclic aromatic hydrocarbons in the water bodies of the Baltic Sea. Mar. Chem. 2002, 79, 49–66. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Algae–bacteria interactions: Evolution, ecology and emerging applications. BioTechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Gorain, P.C.; Bagchi, S.K.; Mallick, N. Effects of calcium, magnesium and sodium chloride in enhancing lipid accumulation in two green microalgae. Environ. Technol. 2013, 34, 1887–1894. [Google Scholar] [CrossRef]
- Affenzeller, M.J.; Darehshouri, A.; Andosch, A.; Lutz, C.; Lutz-Meindl, U. Salt stress-induced cell death in the unicellular green alga Micrasterias denticulata. J. Exp. Bot. 2009, 60, 939–954. [Google Scholar] [CrossRef] [Green Version]
- Salama, E.-S.; Kim, H.-C.; Abou-Shanab, R.A.I.; Ji, M.-K.; Oh, Y.-K.; Kim, S.-H.; Jeon, B.-H. Biomass, lipid content, and fatty acid composition of freshwater Chlamydomonas mexicana and Scenedesmus obliquus grown under salt stress. Bioprocess Biosyst. Eng. 2013, 36, 827–833. [Google Scholar] [CrossRef]
- O’Connell, D.W.; Birkinshaw, C.; O’Dwyer, T.F. Heavy metal adsorbents prepared from the modification of cellulose: A review. Bioresour. Technol. 2008, 99, 6709–6724. [Google Scholar] [CrossRef]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.; Jeffryes, C. Biosynthetic conversion of Ag+ to highly stable Ag0 nanoparticles by wild type and cell wall deficient strains of Chlamydomonas reinhardtii. Molecules 2018, 24, 98. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, C. Biosorbents for heavy metals removal and their future. BioTechnol. Adv 2009, 27, 195–226. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Kumar, S.; Bafana, A.; Dahoumane, S.A.; Jeffryes, C. Individual and combined effects of extracellular polymeric substances and whole cell components of Chlamydomonas reinhardtii on silver nanoparticle synthesis and stability. Molecules 2019, 24, 956. [Google Scholar] [CrossRef] [Green Version]
- Finocchio, E.; Lodi, A.; Solisio, C.; Converti, A. Chromium (VI) removal by methylated biomass of Spirulina platensis: The effect of methylation process. Chem. Eng. J. 2010, 156, 264–269. [Google Scholar] [CrossRef]
- Rahman, A. Biosynthetic conversion of Ag+ to colloidally stable Ag0 nanoparticles by Chlamydomonas reinhardtii: A Mechanistic View of the Light-Induced Synthesis Process and the Impact of Extracellular Polymeric Substances on the Stability of the Nanoparticles. Ph.D. Thesis, Lamar University, Beaumont, TX, USA, 2019. [Google Scholar]
- Rahman, A.; Kumar, S.; Nawaz, T. Chapter 17—Biosynthesis of nanomaterials using algae. In Microalgae Cultivation for Biofuels Production; Yousuf, A., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 265–279. ISBN 978-0-12-817536-1. [Google Scholar]
- Volesky, B. Biosorption and me. Water Res. 2007, 41, 4017–4029. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.; Carmen Bartolomé, M.; Sánchez-Fortún, S. Influence of pH on the survival of Dictyosphaerium chlorelloides populations living in aquatic environments highly contaminated with chromium. Ecotox. Environ. Safe 2013, 98, 82–87. [Google Scholar] [CrossRef] [PubMed]
- López-Rodas, V.; Marvá, F.; Rouco, M.; Costas, E.; Flores-Moya, A. Adaptation of the chlorophycean Dictyosphaerium chlorelloides to stressful acidic, mine metal-rich waters as result of pre-selective mutations. Chemosphere 2008, 72, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.S.; Rodrigues, M.S.; de Carvalho, J.C.M.; Lodi, A.; Finocchio, E.; Perego, P.; Converti, A. Adsorption of Ni2+, Zn2+ and Pb2+ onto dry biomass of Arthrospira (Spirulina) platensis and Chlorella vulgaris. I. single metal systems. Chem. Eng. J. 2011, 173, 326–333. [Google Scholar] [CrossRef]
- Ojeda-Barrios, D.; Abadía, J.; Lombardini, L.; Abadía, A.; Vázquez, S. Zinc deficiency in field-grown pecan trees: Changes in leaf nutrient concentrations and structure. J. Sci. Food Agric. 2012, 92, 1672–1678. [Google Scholar] [CrossRef]
- Öncel, I.; Keleş, Y.; Üstün, A.S. Interactive effects of temperature and heavy metal stress on the growth and some biochemical compounds in wheat seedlings. Environ. Pollut. 2000, 107, 315–320. [Google Scholar] [CrossRef]
- Lefebvre, O.; Moletta, R. Treatment of organic pollution in industrial saline wastewater: A literature review. Water Res. 2006, 40, 3671–3682. [Google Scholar] [CrossRef]
- Mahdavi, H.; Prasad, V.; Liu, Y.; Ulrich, A.C. In situ biodegradation of naphthenic acids in oil sands tailings pond water using indigenous algae–bacteria consortium. Bioresour. Technol. 2015, 187, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Guo, X.; Su, G.; Danquah, M.K.; Zhang, S.; Lu, Y.; Sun, Y.; Lin, L. Bioprocess considerations for microalgal-based wastewater treatment and biomass production. Renew. Sustain. Energy Rev. 2015, 42, 1385–1392. [Google Scholar] [CrossRef]
- Richardson, J.W.; Johnson, M.D.; Lacey, R.; Oyler, J.; Capareda, S. Harvesting and extraction technology contributions to algae biofuels economic viability. Algal. Res. 2014, 5, 70–78. [Google Scholar] [CrossRef]
- Brennan, L.; Owende, P. Biofuels from microalgae: Towards meeting advanced fuel standards. In Advanced Biofuels and Bioproducts; Springer: New York, NY, USA, 2013; pp. 553–599. [Google Scholar]
- Coons, J.E.; Kalb, D.M.; Dale, T.; Marrone, B.L. Getting to low-cost algal biofuels: A monograph on conventional and cutting-edge harvesting and extraction technologies. Algal. Res. 2014, 6, 250–270. [Google Scholar] [CrossRef] [Green Version]
- Roselet, F.; Vandamme, D.; Roselet, M.; Muylaert, K.; Abreu, P.C. Screening of commercial natural and synthetic cationic polymers for flocculation of freshwater and marine microalgae and effects of molecular weight and charge density. Algal. Res. 2015, 10, 183–188. [Google Scholar] [CrossRef]
- Schlesinger, A.; Eisenstadt, D.; Bar-Gil, A.; Carmely, H.; Einbinder, S.; Gressel, J. Inexpensive non-toxic flocculation of microalgae contradicts theories; overcoming a major hurdle to bulk algal production. BioTechnol. Adv. 2012, 30, 1023–1030. [Google Scholar] [CrossRef]
- Emamjomeh, M.M.; Sivakumar, M. Review of pollutants removed by electrocoagulation and electrocoagulation/flotation processes. J. Environ. Manag. 2009, 90, 1663–1679. [Google Scholar] [CrossRef]
- Lakaniemi, A.-M.; Tuovinen, O.H.; Puhakka, J.A. Anaerobic conversion of microalgal biomass to sustainable energy carriers—A review. Bioresour. Technol. 2013, 135, 222–231. [Google Scholar] [CrossRef]
- Lee, O.K.; Kim, A.L.; Seong, D.H.; Lee, C.G.; Jung, Y.T.; Lee, J.W.; Lee, E.Y. Chemo-enzymatic saccharification and bioethanol fermentation of lipid-extracted residual biomass of the microalga, Dunaliella tertiolecta. Bioresour. Technol. 2013, 132, 197–201. [Google Scholar] [CrossRef]
- Tang, H.; Abunasser, N.; Garcia, M.E.D.; Chen, M.; Simon Ng, K.Y.; Salley, S.O. Potential of microalgae oil from Dunaliella tertiolecta as a feedstock for biodiesel. Appl. Energy 2011, 88, 3324–3330. [Google Scholar] [CrossRef]
- Kim, S.-S.; Ly, H.V.; Kim, J.; Lee, E.Y.; Woo, H.C. Pyrolysis of microalgae residual biomass derived from Dunaliella tertiolecta after lipid extraction and carbohydrate saccharification. Chem. Eng. J. 2015, 263, 194–199. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, N.; Wu, Y.; Wu, K.; Wu, X.; Liu, J.; Yang, M. Distributions of organic compounds to the products from hydrothermal liquefaction of microalgae. Environ. Prog. Sustain. 2017, 36, 259–268. [Google Scholar] [CrossRef]
- Richardson, J.W.; Johnson, M.D. Financial feasibility analysis of NAABB developed technologies. Algal. Res. 2015, 10, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Frank, E.D.; Elgowainy, A.; Han, J.; Wang, Z. Life cycle comparison of hydrothermal liquefaction and lipid extraction pathways to renewable diesel from algae. Mitig. Adapt. Strat. Gl. 2013, 18, 137–158. [Google Scholar] [CrossRef] [Green Version]
- Uduman, N.; Qi, Y.; Danquah, M.K.; Forde, G.M.; Hoadley, A. Dewatering of microalgal cultures: A major bottleneck to algae-based fuels. J. Renew. Sustain. Energy 2010, 2, 012701. [Google Scholar] [CrossRef]
- Hu, Y.; Feng, S.; Yuan, Z.; Xu, C.C.; Bassi, A. Investigation of aqueous phase recycling for improving bio-crude oil yield in hydrothermal liquefaction of algae. Bioresour. Technol. 2017, 239, 151–159. [Google Scholar] [CrossRef]
- Chisti, Y. Constraints to commercialization of algal fuels. J. Biotechnol. 2013, 167, 201–214. [Google Scholar] [CrossRef]
- Chaudhary, B.K.; Willman, S.E.; Researcher, P.-M.; Carroll, K.C. Spatial Variability and Geochemistry of Produced Water in Southeastern New Mexico, USA; New Mexico Water Resources Research Institute: Las Cruces, NM, USA, 2016; p. 33. [Google Scholar]
- Ishika, T.; Moheimani, N.R.; Bahri, P.A. Sustainable saline microalgae co-cultivation for biofuel production: A critical review. Renew. Sust. Energy Rev. 2017, 78, 356–368. [Google Scholar] [CrossRef]
- Guieysse, B.; Béchet, Q.; Shilton, A. Variability and uncertainty in water demand and water footprint assessments of fresh algae cultivation based on case studies from five climatic regions. Bioresour. Technol. 2013, 128, 317–323. [Google Scholar] [CrossRef]
- Stephens, E.; Ross, I.L.; King, Z.; Mussgnug, J.H.; Kruse, O.; Posten, C.; Borowitzka, M.A.; Hankamer, B. An economic and technical evaluation of microalgal biofuels. New Biotechnol. 2010, 28, 126–128. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Guria, C.; Pathak, A.K. Potential CO2 fixation and optimal Dunaliella tertiolecta cultivation: Influence of fertilizer, wavelength of light-emitting diodes, salinity and carbon supply strategy. J. CO2 Util. 2017, 22, 164–177. [Google Scholar] [CrossRef]
- Andersen, R.A. Algal Culturing Techniques; Academic Press: Cambridge, MA, USA, 2005; ISBN 978-0-12-088426-1. [Google Scholar]
- Mendes, L.B.B.; Vermelho, A.B. Allelopathy as a potential strategy to improve microalgae cultivation. BioTechnol. Biofuels 2013, 6, 1–14. [Google Scholar]
- Graneli, E.; Weberg, M.; Salomon, P.S. Harmful algal blooms of allelopathic microalgal species: The role of eutrophication. Harmful Algae 2008, 8, 94–102. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; Khairy, H.M. A trial for biological control of a pathogenic fungus (Fusarium solani) by some marine microorganisms. Environ. Sci. 2009, 8, 434–440. [Google Scholar]
- Lutzu, G.A.; Dunford, N.T. Algal treatment of wastewater generated during oil and gas production using hydraulic fracturing technology. Environ. Technol. 2019, 40, 1027–1034. [Google Scholar] [CrossRef]
- Taikhao, S.; Incharoensakdi, A.; Phunpruch, S. Dark fermentative hydrogen production by the unicellular halotolerant cyanobacterium Aphanothece halophytica grown in seawater. J. Appl. Phycol. 2015, 27, 187–196. [Google Scholar] [CrossRef]
- Papry, R.I.; Ishii, K.; Mamun, M.A.A.; Miah, S.; Naito, K.; Mashio, A.S.; Maki, T.; Hasegawa, H. Arsenic biotransformation potential of six marine diatom species: Effect of temperature and salinity. Sci. Rep.-UK 2019, 9, 10226. [Google Scholar] [CrossRef] [Green Version]
- Shikata, T.; Matsubara, T.; Yoshida, M.; Sakamoto, S.; Yamaguchi, M. Effects of temperature, salinity, and photosynthetic photon flux density on the growth of the harmful diatom Asteroplanus karianus in the Ariake Sea, Japan. Fisheries Sci. 2015, 81, 1063–1069. [Google Scholar] [CrossRef]
- Paul, J.T.; Ramaiah, N.; Sardessai, S. Nutrient regimes and their effect on distribution of phytoplankton in the Bay of Bengal. Mar. Environ. Res. 2008, 66, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Saros, J.E. Changes in the growth rates of saline-lake diatoms in response to variation in salinity, brine type and nitrogen form. J. Plankton Res. 2000, 22, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Pahl, S.L.; Lewis, D.M.; Chen, F.; King, K.D. Heterotrophic growth and nutritional aspects of the diatom Cyclotella cryptica (Bacillariophyceae): Effect of some environmental factors. J. Biosci. Bioeng. 2010, 109, 235–239. [Google Scholar] [CrossRef]
- Hevia-Orube, J.; Orive, E.; David, H.; Díez, A.; Laza-Martínez, A.; Miguel, I.; Seoane, S. Molecular and morphological analyses of solitary forms of brackish Thalassiosiroid diatoms (Coscinodiscophyceae), with emphasis on their phenotypic plasticity. Eur J. Phycol. 2016, 51, 11–30. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, Y.; Nagumo, T.; Kato, M. A New Variety of Cyclotella atomus from Tokyo Bay, Japan; C. atomus var. marina var. nov. Bull. Nat. Sci. Mus. 2004, 30, 5–11. [Google Scholar]
- Wen, Z.; Mian-ping, Z.; Xian-zhong, X.; Xi-Fang, L.; Gan-lin, G.; Zhi-hui, H. Biological and ecological features of saline lakes in northern Tibet, China. Hydrobiologia 2005, 541, 189–203. [Google Scholar] [CrossRef]
- Rynearson, T.A.; Newton, J.A.; Armbrust, E.V. Spring bloom development, genetic variation, and population succession in the planktonic diatom Ditylum brightwellii. Limnol. Oceanogr. 2006, 51, 1249–1261. [Google Scholar] [CrossRef]
- Papry, R.I.; Omori, Y.; Fujisawa, S.; Al Mamun, M.A.; Miah, S.; Mashio, A.S.; Maki, T.; Hasegawa, H. Arsenic biotransformation potential of marine phytoplankton under a salinity gradient. Algal. Res. 2020, 47, 101842. [Google Scholar] [CrossRef]
- Xu, N.; Pang, S.; Shan, T.; Liu, F.; Zhao, X.; Gao, S. Molecular identification and culture trials of Eutreptiella gymnastica (Eutreptiales, Euglenophyceae). Chin. J. Oceanol. Limn. 2012, 30, 446–455. [Google Scholar] [CrossRef]
- Jeong, H.J.; Kim, T.H.; Yoo, Y.D.; Yoon, E.Y.; Kim, J.S.; Seong, K.A.; Kim, K.Y.; Park, J.Y. Grazing impact of heterotrophic dinoflagellates and ciliates on common red-tide euglenophyte Eutreptiella gymnastica in Masan Bay, Korea. Harmful Algae 2011, 10, 576–588. [Google Scholar] [CrossRef]
- Carić, M.; Jasprica, N.; Čalić, M.; Batistić, M. Phytoplankton response to high salinity and nutrient limitation in the eastern Adriatic marine lakes. Sci. Mar. 2011, 75, 493–505. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Lambert, W.; Giang, S.; Goericke, R.; Palenik, B. Microalgal assemblages in a poikilohaline pond. J. Phycol. 2014, 50, 303–309. [Google Scholar] [CrossRef]
- Wang, S.; Shi, X.; Palenik, B. Characterization of Picochlorum sp. use of wastewater generated from hydrothermal liquefaction as a nitrogen source. Algal. Res. 2016, 13, 311–317. [Google Scholar] [CrossRef] [Green Version]
- Foflonker, F.; Ananyev, G.; Qiu, H.; Morrison, A.; Palenik, B.; Dismukes, G.C.; Bhattacharya, D. The unexpected extremophile: Tolerance to fluctuating salinity in the green alga Picochlorum. Algal. Res. 2016, 16, 465–472. [Google Scholar] [CrossRef] [Green Version]
- Foflonker, F.; Mollegard, D.; Ong, M.; Yoon, H.S.; Bhattacharya, D. Genomic analysis of Picochlorum species reveals how microalgae may adapt to variable environments. Mol. Biol. Evol. 2018, 35, 2702–2711. [Google Scholar] [CrossRef] [Green Version]
- Pálmai, T.; Szabó, B.; Kotut, K.; Krienitz, L.; Padisák, J. Ecophysiology of a successful phytoplankton competitor in the African flamingo lakes: The green alga Picocystis salinarum (Picocystophyceae). J. Appl. Phycol. 2020, 32, 1813–1825. [Google Scholar] [CrossRef] [Green Version]
- Fanjing, K.; Qinxian, J.; Jia, E.; Mianping, Z. Characterization of a eukaryotic picoplankton alga, strain DGN-Z1, isolated from a soda lake in inner Mongolia, China. Nat. Resour. Env. Iss. 2009, 15, 38. [Google Scholar]
- Krienitz, L.; Bock, C.; Kotut, K.; Luo, W. Picocystis salinarum (Chlorophyta) in saline lakes and hot springs of East Africa. Phycologia 2012, 51, 22–32. [Google Scholar] [CrossRef]
- Brooks, B.W.; James, S.V.; Valenti, T.W., Jr.; Urena-Boeck, F.; Serrano, C.; Berninger, J.P.; Schwierzke, L.; Mydlarz, L.D.; Grover, J.P.; Roelke, D.L. Comparative toxicity of Prymnesium parvum in inland waters. J. Am. Water Resour As. 2010, 46, 45–62. [Google Scholar] [CrossRef]
- Granéli, E.; Salomon, P.S. Factors influencing allelopathy and toxicity in Prymnesium parvum. J. Am. Water Resour As. 2010, 46, 108–120. [Google Scholar] [CrossRef]
- Balzano, S.; Sarno, D.; Kooistra, W.H.C.F. Effects of salinity on the growth rate and morphology of ten Skeletonema strains. J. Plankton Res. 2011, 33, 937–945. [Google Scholar] [CrossRef] [Green Version]
- Demir, N. Changes in the phytoplankton community of a coastal, hyposaline lake in western Anatolia, Turkey. Limnology 2007, 8, 337–342. [Google Scholar] [CrossRef]
- Costa-Böddeker, S.; Thuyên, L.X.; Schwarz, A.; Huy, H.Đ.; Schwalb, A. Diatom assemblages in surface sediments along nutrient and salinity gradients of Thi Vai Estuary and Can Gio Mangrove Forest, Southern Vietnam. Estuar. Coast 2017, 40, 479–492. [Google Scholar] [CrossRef]
- Craggs, R.J.; Heubeck, S.; Lundquist, T.J.; Benemann, J.R. Algal biofuels from wastewater treatment high rate algal ponds. Water Sci. Technol. 2011, 63, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.K.; Craggs, R.J.; Shilton, A.N. Wastewater treatment high rate algal ponds for biofuel production. Bioresour. Technol. 2011, 102, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Vendramel, S.; Bassin, J.P.; Dezotti, M.; Sant’Anna, G.L., Jr. Treatment of petroleum refinery wastewater containing heavily polluting substances in an aerobic submerged fixed-bed reactor. Environ. Technol. 2015, 36, 2052–2059. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Exploration of algae in algae-based PW systems (data until July 2020) (a) number of algal species explored per phylum; (b) breakdown of freshwater, marine and freshwater/marine strains.
Figure 1.
Exploration of algae in algae-based PW systems (data until July 2020) (a) number of algal species explored per phylum; (b) breakdown of freshwater, marine and freshwater/marine strains.

Figure 2.
Major steps to promote the cultivation of algae in PW.

Figure 3.
The major factors that affect the large-scale development of algal-based PW system.


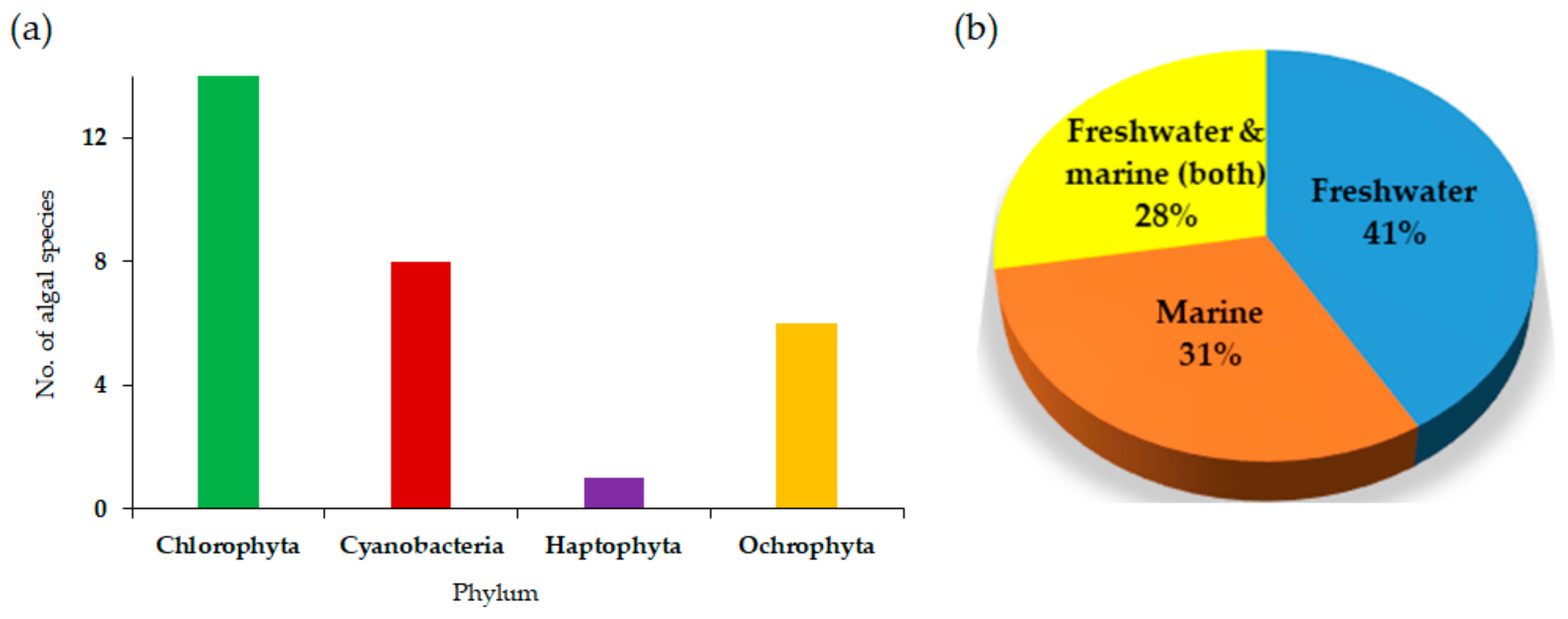{kind=link}
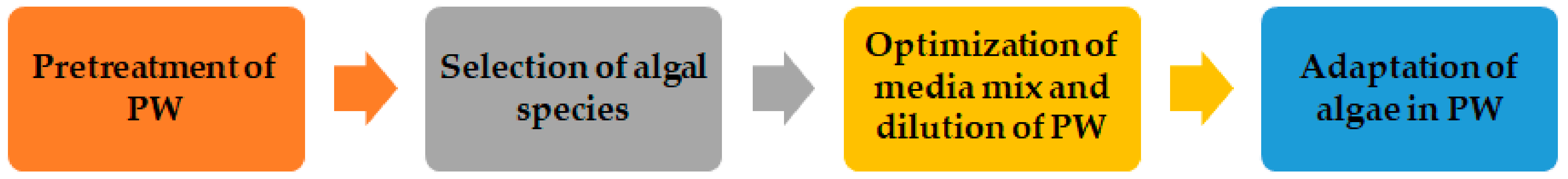{kind=link}
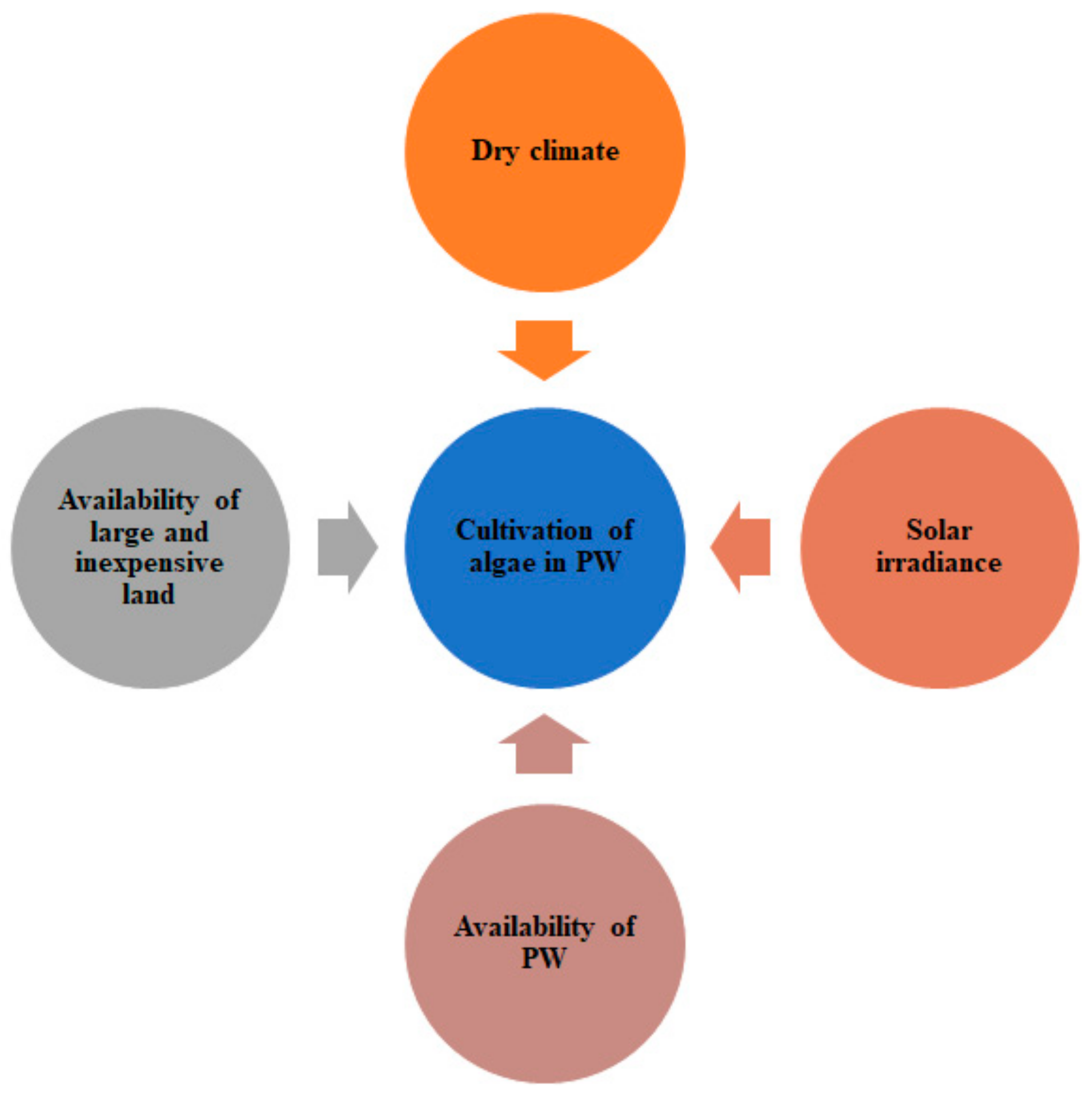{kind=link}
Table 1.
A comprehensive list of produced water (PW) parameters with typical range.
| Parameter Group | Parameter | Unit | Typical Range | Ref |
|---|---|---|---|---|
| General water quality | pH | S.U. | 6.0–8.5 | [39,40,43,44,45] |
| Density | Kg/m3 | ~1021 | [45] | |
| Salinity | mg/L NaCl | 52,700–633,000 | [39] | |
| Electrical conductivity (EC) | mS/cm | 30–600 | [39,40,43,45] | |
| Turbidity | NTU | ~206 | [45] | |
| Total suspended solids (TSS) | mg L−1 | 20–254 | [39,45,46] | |
| Total dissolved solids (TDS) | mg L−1 | 2650–138,500 | [39,40,44,45,46] | |
| Suspended solids (SS) | mg L−1 | ~550 | [44] | |
| Oil and grease | mg L−1 | 73–540 | [44] | |
| Dissolved organic carbon (DOC) | mg L−1 | 270–470 | [43] | |
| Total organic carbon (TOC) | mg L−1 | 55–63 | [39,44,46] | |
| Total nitrogen (TN) | mg L−1 | 30–34 | [43] | |
| Total phosphorus (TP) | mg L−1 | 0.08–0.2 | [44,46] | |
| Biological oxygen demand (BOD5) | mg L−1 | 5–86 | [44,45] | |
| Chemical oxygen demand (COD) | mg L−1 | 177–1300 | [39,44] | |
| Sodium Adsorption ratio (SAR) | mg L−1 | ~182 | [46] | |
| Polyatomic ions/radicals | Ammonium (NH4+) | mg L−1 | 8–20 | [40,44,46] |
| Bicarbonate (HCO3−) | mg L−1 | 71–263 | [44,45] | |
| Nitrate | mg L−1 | ~622 | [43,46] | |
| Nitrite | mg L−1 | 8.2–9.2 | [43] | |
| Phosphate (PO4) | mg L−1 | 2–300 | [39,46] | |
| Halides | Fluorine | mg L−1 | 0.3–204 | [40] |
| Bromide | mg L−1 | 126–134 | [43] | |
| Chloride | mg L−1 | 750–11,500 | [40,43,44,45,46] | |
| Iodide | mg L−1 | 46–48 | [43] | |
| Cations | Aluminium | mg L−1 | 0.01–1.5 | [39,40] |
| Barium | mg L−1 | 0.1–172 | [39,40] | |
| Boron | mg L−1 | 5–11 | [40] | |
| Calcium | mg L−1 | 4–7800 | [39,40,43,44,46] | |
| Iron | mg L−1 | 0.1–40 | [39,40,43,45,46] | |
| Lithium | mg L−1 | 0.1–507 | [39,40,46] | |
| Magnesium | mg L−1 | 2–700 | [39,40,43,45,46] | |
| Manganese | mg L−1 | 0.1–5.8 | [39,40,43,45] | |
| Potassium | mg L−1 | 170–390 | [39,44,46] | |
| Sodium | mg L−1 | 850–20,000 | [39,43,46] | |
| Heavy metals | Chromium | mg L−1 | 0.1–0.2 | [39,45] |
| Copper | mg L−1 | 0.06–4.2 | [39,40] | |
| Lead | mg L−1 | ~0.03 | [39] | |
| Nickel | mg L−1 | 0.06–0.2 | [39,45] | |
| Strontium | mg L−1 | 360–470 | [39] | |
| Zinc | mg L−1 | 0.003–1 | [39,40,45] |
Table 2.
Algae deployed in PW.
| Phylum | Name | Type of Algae | Assessment | PW Loading | Medium | Cultivation Type | Potential for Bioremediation | Ref | |
|---|---|---|---|---|---|---|---|---|---|
| Biomass Productivity | Bioremediation Potential | ||||||||
| Chlorophyta | Chlamydomonas reinhardtii | freshwater | ✓ | 0%, 20%, 40%, 60%, 80%, 100% | TAP | [50] | |||
| Chlorella sp. | freshwater | ✓ | ✓ | 0%, 25%, 50%, 100% PW loading [61] | H2 medium [62], BG-11 [4], f/2 [61], Broad seawater medium [47] | 1 L Photobioreactor (PBR), 450 mL Tube reactors [48] | TOC, TN, Al, Zn, Fe | [4,11,47,61,62,63] | |
| Dictyosphaerium sp. | freshwater/marine | ✓ | ✓ | 50%, 60%, 75%, 100% | Al, Zn, Fe | [11,63] | |||
| Dunaliella salina | marine | ✓ | ✓ | artificial seawater [39], Seawater [46] | 125 mL conical flask | Zn, Ni | [39,46] | ||
| Dunaliella tertiolecta | marine | ✓ | f/2 | [60] | |||||
| Monoraphidium sp. | freshwater | ✓ | ✓ | 50%, 60%, 75%,100% [63] | H2 medium [62], BG-11 [4] | Al, Zn, Fe | [4,11,62,63] | ||
| Neochloris oleoabundans | freshwater/marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | Al, Zn, Fe | [11,47] | |||
| Parachlorella kessleri | freshwater | ✓ | f/2 | [41] | |||||
| Picochlorum sp. | marine | ✓ | f/2 | [4] | |||||
| Pseudokirchneriella subcapitata | freshwater | ✓ | [64] | ||||||
| Scenedesmus sp. | freshwater | ✓ | ✓ | 50%, 60%, 75%, 100% [63] | Wuxal (W), BG-11 [4] | Open pond-600 L [49] | Al, Zn, Fe | [4,11,49,63] | |
| Scenedesmus rotundus | freshwater | ✓ | [51] | ||||||
| Tetracystis sp. | freshwater | ✓ | [32] | ||||||
| Tetraselmis sp. | marine | ✓ | f/2 | [4] | |||||
| Ochrophyta | Amphora coffeaformis | freshwater/marine | ✓ | ✓ | H2 medium [62], Broad seawater medium [47] | 450 mL Tube reactors [48] | Ba, Ca, Fe, Si, Mn | [47,62] | |
| Chaetoceros gracilis | marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | Ba, Ca, Fe, Si, Mn | [47] | |||
| Chaetoceros muelleri | marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | [47] | ||||
| Nannochloropsis oculata | marine | ✓ | ✓ | 10–50% [45], 0%, 50%, 100% [42] | BG11 [45], f/2 [42] | Chemical oxygen demand (COD) and Oil content | [42,45] | ||
| Nannochloropsis salina | freshwater/marine | ✓ | f/2-Si (minus silica) | [32] | |||||
| Phaeodactylum tricornutum | freshwater/marine | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | Ba, Ca, Fe, Si, Mn | [47] | |||
| Cyanobacteria | Cyanobacterium aponinum | freshwater/marine | ✓ | f/2 | [41] | ||||
| Cyanobium sp. | freshwater | ✓ | ✓ | H2 medium | phosphate | [62] | |||
| Logan Lagoons Cyanobacteria 2 (LLC2) | freshwater/marine | ✓ | [65] | ||||||
| Lyptolyngbya sp. | marine | ✓ | f/2 | [4] | |||||
| Phormidium sp. | freshwater | ✓ | ✓ | H2 medium | ammonia, phosphate | [62] | |||
| Pseudoanabaena sp. | freshwater | ✓ | H2 medium | [62] | |||||
| Synechocystis sp. PCC 6803 | freshwater | ✓ | Broad seawater medium [47] | 450 mL Tube reactors | [47] | ||||
| Synechococcus elongatus | freshwater | ✓ | 0%, 20%, 40%, 60%, 80%, 100% | Broad seawater medium [47] | 450 mL Tube reactors | [47] | |||
| Haptophyta | Isochrysis galbana | marine | ✓ | ✓ | 0%, 25%, 50%, 100% PW loading [61] | BG-11 | COD and Oil content | [45] | |
Table 3.
A list of algal species that can thrive in wide range of salinity (salinity presented in per mille (‰) unit).
Table 3.
A list of algal species that can thrive in wide range of salinity (salinity presented in per mille (‰) unit).
| Species (In Alphabetical Order) | Phylum | Salinity Range (‰) | Ref |
|---|---|---|---|
| Aphanothece sp. | Cyanobacteria | 0–175 | [117,118] |
| Asteroplanus karianus | Bacillariophyta | 10–35 | [119,120] |
| Chaetoceros sp. (e.g., Chaetoceros didymus, Chaetoceros elmorei) | Bacillariophyta | 10–35 | [119,121,122] |
| Cyanobacterium aponinum | Cyanobacteria | 15–60 | [41] |
| Cyclotella sp. (e.g., Cyclotella cryptica, Cyclotella marina, Cyclotella quillensis) | Bacillariophyta | 3–90 | [123,124,125] |
| Cymbella pusilla | Bacillariophyta | 3–200 | [122,126] |
| Ditylum brightwellii | Heterokonta | 10–35 | [119,127] |
| Dunaliella sp. (e.g., Dunaliella tertiolecta, Dunaliella salina) | Chlorophyta | 5–360 | [39,60] |
| Eutreptiella gymnestica | Euglenozoa | 5–30 | [128,129,130] |
| Nannochloropsis sp. (e.g., Nannochloropsis oculata) | Ochrophyta | 40 | [42,57,60] |
| Nitzschia longissima | Bacillariophyta | 10–35 | [119,131] |
| Oltmannsiellopsis viridis | Chlorophyta | 5–30 | [128] |
| Parachlorella kessleri | Chlorophyta | 15–60 | [41] |
| Picochlorum sp. (e.g., Picochlorum oklahomensis) | Chlorophyta | 35–150 | [132,133,134,135] |
| Picocystis salinarum | Chlorophyta | 15–300 | [136,137,138] |
| Prymnesium parvum | Haptophyta | 3–30 | [128,139,140] |
| Skeletonema sp. (e.g., Skeletonema costatum, Skeletonema subsalsum) | Bacillariophyta | 0–35 | [119,141] |
| Thalassionema nitzschioides | Bacillariophyta | 10–35 | [119,142,143] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rahman, A.; Agrawal, S.; Nawaz, T.; Pan, S.; Selvaratnam, T. A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production. Water 2020, 12, 2351. https://0-doi-org.brum.beds.ac.uk/10.3390/w12092351
AMA Style
Rahman A, Agrawal S, Nawaz T, Pan S, Selvaratnam T. A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production. Water. 2020; 12(9):2351. https://0-doi-org.brum.beds.ac.uk/10.3390/w12092351
Chicago/Turabian StyleRahman, Ashiqur, Saumya Agrawal, Tabish Nawaz, Shanglei Pan, and Thinesh Selvaratnam. 2020. "A Review of Algae-Based Produced Water Treatment for Biomass and Biofuel Production" Water 12, no. 9: 2351. https://0-doi-org.brum.beds.ac.uk/10.3390/w12092351
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.