
Phenotypic and Gene Expression Profiles of Embryo Development of the Ascidian Ciona robusta Exposed to Dispersants



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials‘ and Methods
2.1. Dispersants Formulation and Preparation
2.2. Animal and Gametes Collection and In Vitro Fertilization
2.3. Embryotoxicity
2.4. RNA Extraction and cDNA Synthesis
2.5. Gene Expression by Real Time-Quantitative PCR (RT-qPCR)
2.6. Statistical Analysis
3. Results and Discussion
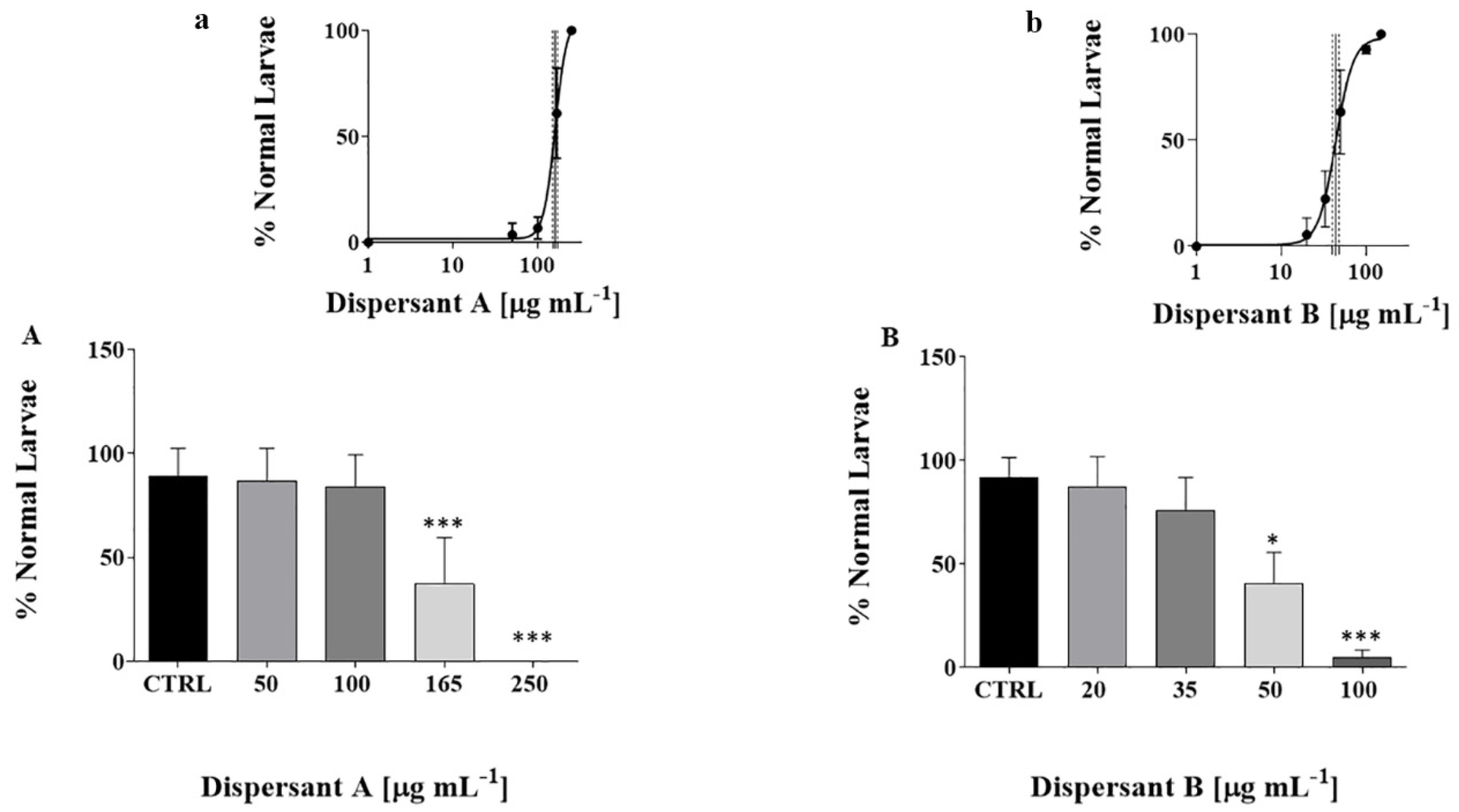
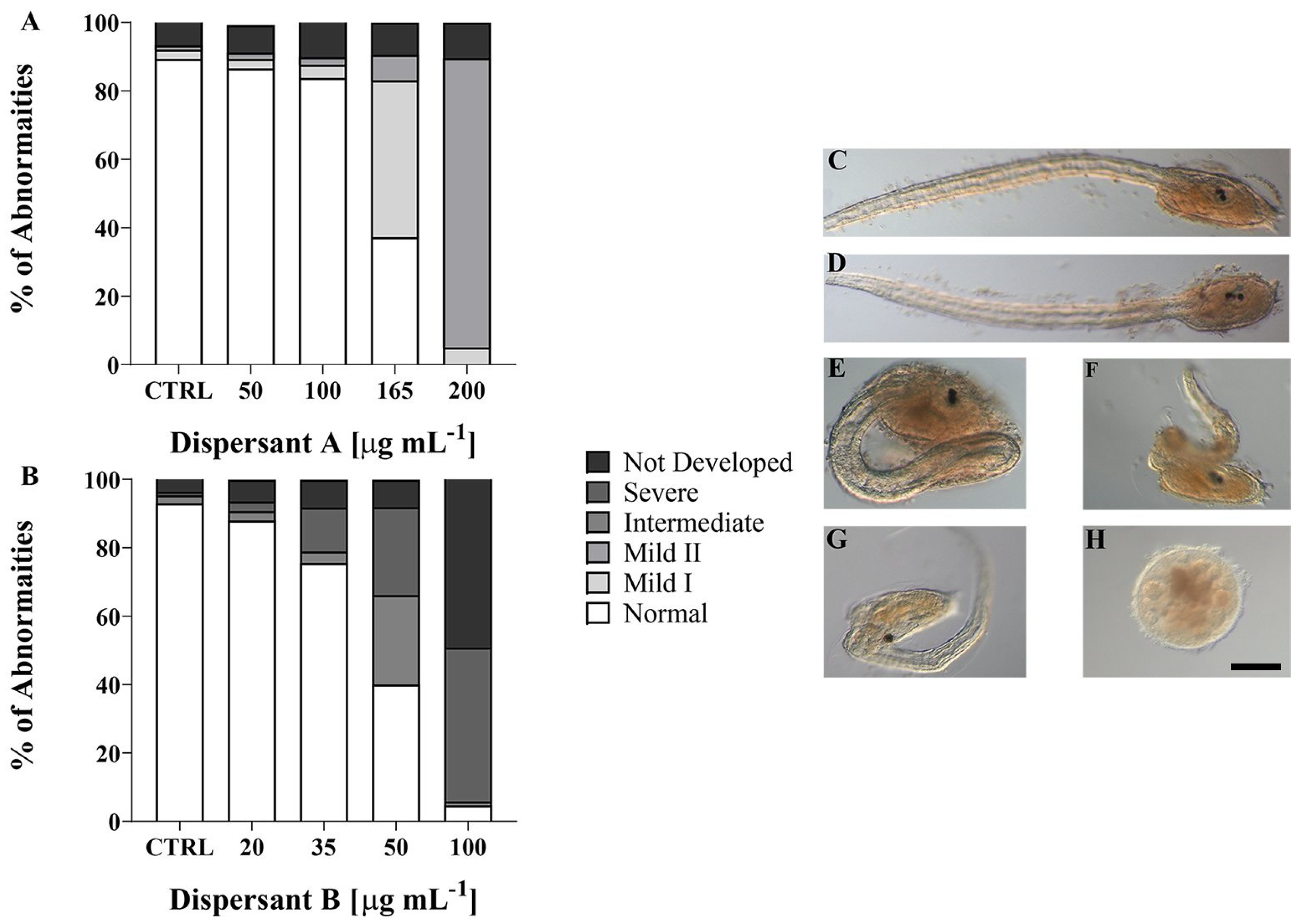
3.1. Larvae Phenotypes
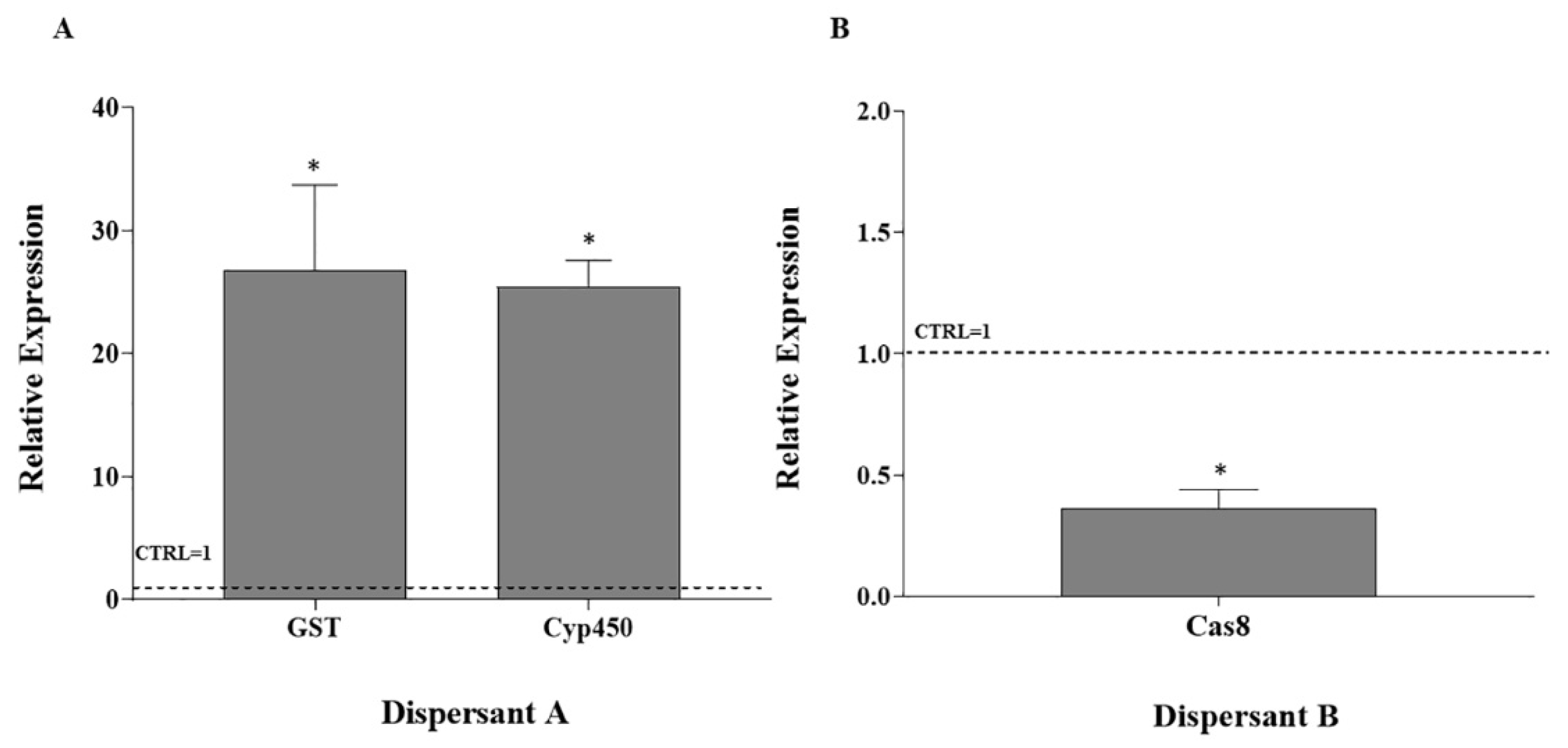
3.2. RT-qPCR on Stress Response Genes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, K.; Nedwed, T.; Prince, R.C.; Palandro, D. Lab Tests on the Biodegradation of Chemically Dispersed Oil Should Consider the Rapid Dilution That Occurs at Sea. Mar. Pollut. Bull. 2013, 73, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Lessard, R.R.; DeMarco, G. The Significance of Oil Spill Dispersants. Spill Sci. Technol. Bull. 2000, 6, 59–68. [Google Scholar] [CrossRef]
- National Research Council (USA). Committee on Effectiveness of Oil Spill Dispersants. In Using Oil Spill Dispersants on the Sea; National Academy Press: Cambridge, MA, USA, 1989; ISBN 9780309038829. [Google Scholar]
- Committee on Understanding Oil Spill Dispersants: Efficacy and Effects; Ocean Studies Board; Division on Earth and Life Studies; National Research Council. Oil Spill Dispersants: Efficacy and Effects; National Academies Press: Cambridge, MA, USA, 2005; ISBN 9780309095624. [Google Scholar]
- Singer, M.M.; Smalheer, D.L.; Tjeerdema, R.S.; Martin, M. Effects of Spiked Exposure to an Oil Dispersant on the Early Life Stages of Four Marine Species. Environ. Toxicol. Chem. 1991, 10, 1367–1374. [Google Scholar] [CrossRef]
- Singer, M.M.; George, S.; Jacobson, S.; Lee, I.; Weetman, L.L.; Tjeerdema, R.S.; Sowby, M.L. Comparison of Acute Aquatic Effects of the Oil Dispersant Corexit 9500 with Those of Other Corexit Series Dispersants. Ecotoxicol. Environ. Saf. 1996, 35, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Bejarano, A.C. Critical Review and Analysis of Aquatic Toxicity Data on Oil Spill Dispersants. Environ. Toxicol. Chem. 2018, 37, 2989–3001. [Google Scholar] [CrossRef] [Green Version]
- International Petroleum Industry Environmental Conservation Association-International Association of Oil & Gas Producers (IPIECA-OGP). Regulatory Approval of Dispersants and Authorization for Their Use; IPIECA-OGP: London, UK, 2014. [Google Scholar]
- European Maritime Safety Agency. In Overview of National Dispersant Testing and Approval Policies in the EU; European Maritime Safety Agency: Lisbon, Portugal, 2016.
- Oil Spill Prevention, Administration and Response (OSPAR). Guidelines for Completing the Harmonised Offshore Chemical Notification Format (HOCNF); OSPAR Agreement: 2012-05; OSPAR Commission: London, UK, 2015. [Google Scholar]
- Wise, J.; Wise, J.P., Sr. A Review of the Toxicity of Chemical Dispersants. Rev. Environ. Health 2011, 26, 281–300. [Google Scholar] [CrossRef]
- Vignier, J.; Donaghy, L.; Soudant, P.; Chu, F.L.E.; Morris, J.M.; Carney, M.W.; Lay, C.; Krasnec, M.; Robert, R.; Volety, A.K. Impacts of Deepwater Horizon Oil and Associated Dispersant on Early Development of the Eastern Oyster Crassostrea Virginica. Mar. Pollut. Bull. 2015, 100, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Adeyemo, O.K.; Kroll, K.J.; Denslow, N.D. Developmental Abnormalities and Differential Expression of Genes Induced in Oil and Dispersant Exposed Menidia Beryllina Embryos. Aquat. Toxicol. 2015, 168, 60–71. [Google Scholar] [CrossRef]
- Beirão, J.; Baillon, L.; Litt, M.A.; Langlois, V.S.; Purchase, C.F. Impact of Crude Oil and the Dispersant CorexitTM EC9500A on Capelin (Mallotus Villosus) Embryo Development. Mar. Environ. Res. 2019, 147, 90–100. [Google Scholar] [CrossRef]
- DeMiguel-Jiménez, L.; Etxebarria, N.; Lekube, X.; Izagirre, U.; Marigómez, I. Influence of Dispersant Application on the Toxicity to Sea Urchin Embryos of Crude and Bunker Oils Representative of Prospective Oil Spill Threats in Arctic and Sub-Arctic Seas. Mar. Pollut. Bull. 2021, 172, 112922. [Google Scholar] [CrossRef]
- Barron, M.G.; Bejarano, A.C.; Conmy, R.N.; Sundaravadivelu, D.; Meyer, P. Toxicity of Oil Spill Response Agents and Crude Oils to Five Aquatic Test Species. Mar. Pollut. Bull. 2020, 153, 110954. [Google Scholar] [CrossRef] [PubMed]
- Pineda, M.C.; McQuaid, C.D.; Turon, X.; López-Legentil, S.; Ordóñez, V.; Rius, M. Tough Adults, Frail Babies: An Analysis of Stress Sensitivity across Early Life-History Stages of Widely Introduced Marine Invertebrates. PLoS ONE 2012, 7, e46672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehal, P.; Satou, Y.; Campbell, R.K.; Chapman, J.; Degnan, B.; De Tomaso, A.; Davidson, B.; Di Gregorio, A.; Gelpke, M.; Goodstein, D.M.; et al. The Draft Genome of Ciona Intestinalis: Insights into Chordate and Vertebrate Origins. Science 2002, 298, 2157–2167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zega, G.; De Bernardi, F.; Groppelli, S.; Pennati, R. Effects of the Azole Fungicide Imazalil on the Development of the Ascidian Ciona Intestinalis (Chordata, Tunicata): Morphological and Molecular Characterization of the Induced Phenotype. Aquat. Toxicol. 2009, 91, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Mizotani, Y.; Itoh, S.; Hotta, K.; Tashiro, E.; Oka, K.; Imoto, M. Evaluation of Drug Toxicity Profiles Based on the Phenotypes of Ascidian Ciona Intestinalis. Biochem. Biophys. Res. Commun. 2015, 463, 656–660. [Google Scholar] [CrossRef]
- Satou, Y.; Imai, K.S. Gene Regulatory Systems That Control Gene Expression in the Ciona Embryo. Proc. Japan Acad. Ser. B 2015, 91, 33–51. [Google Scholar] [CrossRef] [Green Version]
- Bellas, J.; Vázquez, E.; Beiras, R. Toxicity of Hg, Cu, Cd, and Cr on Early Developmental Stages of Ciona Intestinalis (Chordata, Ascidiacea) with Potential Application in Marine Water Quality Assessment. Water Res. 2001, 35, 2905–2912. [Google Scholar] [CrossRef]
- Bellas, J.; Beiras, R.; Mariño-Balsa, J.C.; Fernández, N. Toxicity of Organic Compounds to Marine Invertebrate Embryos and Larvae: A Comparison between the Sea Urchin Embryogenesis Bioassay and Alternative Test Species. Ecotoxicology 2005, 14, 337–353. [Google Scholar] [CrossRef]
- Bellas, J. Comparative Toxicity of Alternative Antifouling Biocides on Embryos and Larvae of Marine Invertebrates. Sci. Total Environ. 2006, 367, 573–585. [Google Scholar] [CrossRef]
- Beiras, R.; Bellas, J. Inhibition of Embryo Development of the Mytilus Galloprovincialis Marine Mussel by Organic Pollutants; Assessment of Risk for Its Extensive Culture in the Galician Rias. Aquaculture 2008, 277, 208–212. [Google Scholar] [CrossRef]
- Eliso, M.C.; Bergami, E.; Manfra, L.; Spagnuolo, A.; Corsi, I. Toxicity of Nanoplastics during the Embryogenesis of the Ascidian Ciona Robusta (Phylum Chordata). Nanotoxicology 2020, 14, 1415–1431. [Google Scholar] [CrossRef] [PubMed]
- Eliso, M.C.; Manfra, L.; Savorelli, F.; Tornambè, A.; Spagnuolo, A. New Approaches on the Use of Tunicates (Ciona Robusta) for Toxicity Assessments. Environ. Sci. Pollut. Res. Int. 2020, 27, 32132–32138. [Google Scholar] [CrossRef] [PubMed]
- Manfra, L.; Tornambè, A.; Guyomarch, J.; Le Guerrogue, P.; Kerambrun, L.; Rotini, A.; Savorelli, F.; Onorati, F.; Magaletti, E. Dispersant Approval Procedures in France and Italy: A Comparative Ecotoxicity Study. Ecotoxicol. Environ. Saf. 2017, 143, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Durier, G.; Nadalini, J.-B.; Saint-Louis, R.; Genard, B.; Comeau, L.A.; Tremblay, R. Sensitivity to Oil Dispersants: Effects on the Valve Movements of the Blue Mussel Mytilus Edulis and the Giant Scallop Placopecten Magellanicus, in Sub-Arctic Conditions. Aquat. Toxicol. 2021, 234, 105797. [Google Scholar] [CrossRef]
- Scarlett, A.; Galloway, T.S.; Canty, M.; Smith, E.L.; Nilsson, J.; Rowland, S.J. Comparative Toxicity of Two Oil Dispersants, Superdispersant-25 and Corexit 9527, to a Range of Coastal Species. Environ. Toxicol. Chem. 2005, 24, 1219–1227. [Google Scholar] [CrossRef]
- Lambert, C.C.; Brandt, C.L. The effect of light on the spawning of Ciona Intestinalis. Biol. Bull. 1967, 132, 222–228. [Google Scholar] [CrossRef]
- Bellas, J.; Beiras, R.; Vázquez, E. A Standardisation of Ciona Intestinalis (Chordata, Ascidiacea) Embryo-Larval Bioassay for Ecotoxicological Studies. Water Res. 2003, 37, 4613–4622. [Google Scholar] [CrossRef]
- Fujikawa, T.; Munakata, T.; Kondo, S.-I.; Satoh, N.; Wada, S. Stress Response in the Ascidian Ciona Intestinalis: Transcriptional Profiling of Genes for the Heat Shock Protein 70 Chaperone System under Heat Stress and Endoplasmic Reticulum Stress. Cell Stress Chaperones 2010, 15, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Colvin, K.A.; Lewis, C.; Galloway, T.S. Current Issues Confounding the Rapid Toxicological Assessment of Oil Spills. Chemosphere 2020, 245, 125585. [Google Scholar] [CrossRef]
- International Maritime Organization. Manual on Oil Pollution, Combating Oil Spills. In Chemical Dispersion; Section IV; International Maritime Organization: London, UK, 2011; Chapter 7. [Google Scholar]
- Fingas, M. Visual Appearance of Oil on the Sea. J. Mar. Sci. Eng. 2021, 9, 97. [Google Scholar] [CrossRef]
- Thompson, H.; Shimeld, S.M. Transmission and Scanning Electron Microscopy of the Accessory Cells and Chorion During Development of Ciona Intestinalis Type B Embryos and the Impact of Their Removal on Cell Morphology. Zoolog. Sci. 2015, 32, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Edmunds, R.C.; Gill, J.A.; Baldwin, D.H.; Linbo, T.L.; French, B.L.; Brown, T.L.; Esbaugh, A.J.; Mager, E.M.; Stieglitz, J.; Hoenig, R.; et al. Corresponding Morphological and Molecular Indicators of Crude Oil Toxicity to the Developing Hearts of Mahi Mahi. Sci. Rep. 2015, 5, 17326. [Google Scholar] [CrossRef] [PubMed]
- Esbaugh, A.J.; Mager, E.M.; Stieglitz, J.D.; Hoenig, R.; Brown, T.L.; French, B.L.; Linbo, T.L.; Lay, C.; Forth, H.; Scholz, N.L.; et al. The Effects of Weathering and Chemical Dispersion on Deepwater Horizon Crude Oil Toxicity to Mahi-Mahi (Coryphaena Hippurus) Early Life Stages. Sci. Total Environ. 2016, 543, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Greer, J.B.; Pasparakis, C.; Stieglitz, J.D.; Benetti, D.; Grosell, M.; Schlenk, D. Effects of Corexit 9500A and Corexit-Crude Oil Mixtures on Transcriptomic Pathways and Developmental Toxicity in Early Life Stage Mahi-Mahi (Coryphaena Hippurus). Aquat. Toxicol. 2019, 212, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Parsi, K. Interaction of Detergent Sclerosants with Cell Membranes. Phlebology 2015, 30, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, A.F.; Meyer, D.N.; Petriv, A.-M.V.; Soto, A.L.; Shields, J.N.; Akemann, C.; Baker, B.B.; Tsou, W.-L.; Zhang, Y.; Baker, T.R. Nanoplastics Impact the Zebrafish (Danio Rerio) Transcriptome: Associated Developmental and Neurobehavioral Consequences. Environ. Pollut. 2020, 266, 115090. [Google Scholar] [CrossRef]
- Aguirre-Ramírez, M.; Silva-Jiménez, H.; Banat, I.M.; Díaz De Rienzo, M.A. Surfactants: Physicochemical Interactions with Biological Macromolecules. Biotechnol. Lett. 2021, 43, 523–535. [Google Scholar] [CrossRef]
- Van der Ven, L.T.M.; Schoonen, W.G.; Groot, R.M.; den Ouden, F.; Heusinkveld, H.J.; Zwart, E.P.; Hodemaekers, H.M.; Rorije, E.; de Knecht, J. The Effects of Aliphatic Alcohols and Related Acid Metabolites in Zebrafish Embryos-Correlations with Rat Developmental Toxicity and with Effects in Advanced Life Stages in Fish. Toxicol. Appl. Pharmacol. 2020, 407, 115249. [Google Scholar] [CrossRef]
- Nagatomo, K.-I.; Ishibashi, T.; Satou, Y.; Satoh, N.; Fujiwara, S. Retinoic Acid Affects Gene Expression and Morphogenesis without Upregulating the Retinoic Acid Receptor in the Ascidian Ciona Intestinalis. Mech. Dev. 2003, 120, 363–372. [Google Scholar] [CrossRef]
- Hylland, K.; Burgeot, T.; Martínez-Gómez, C.; Lang, T.; Robinson, C.D.; Svavarsson, J.; Thain, J.E.; Vethaak, A.D.; Gubbins, M.J. How Can We Quantify Impacts of Contaminants in Marine Ecosystems? Mar. Environ. Res. 2017, 124, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Nebert, D.W.; Dalton, T.P. The Role of Cytochrome P450 Enzymes in Endogenous Signalling Pathways and Environmental Carcinogenesis. Nat. Rev. Cancer 2006, 6, 947–960. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H. Activation and Detoxification Enzymes: Functions and Implications; Springer: Berlin/Heidelberg, Germany, 2011; ISBN 9781461410492. [Google Scholar]
- Portnoy, D.S.; Fields, A.T.; Greer, J.B.; Schlenk, D. Genetics and Oil: Transcriptomics, Epigenetics, and Population Genomics as Tools to Understand Animal Responses to Exposure Across Different Time Scales. In Deep Oil Spills: Facts, Fate, and Effects; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Murawski, S.A.; Ainsworth, C.H.; Gilbert, S.; Hollander, D.J.; Paris, C.B.; Schlüter, M.; Wetzel, D.L. (Eds.) Deep Oil Spills: Facts, Fate, and Effects; Springer International Publishing: Cham, Germany, 2020; pp. 515–532. ISBN 9783030116057. [Google Scholar]
- Kumagai, T.; Suzuki, H.; Sasaki, T.; Sakaguchi, S.; Miyairi, S.; Yamazoe, Y.; Nagata, K. Polycyclic Aromatic Hydrocarbons Activate CYP3A4 Gene Transcription through Human Pregnane X Receptor. Drug Metab. Pharmacokinet. 2012, 27, 200–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenny, M.J.; Karchner, S.I.; Franks, D.G.; Woodin, B.R.; Stegeman, J.J.; Hahn, M.E. Distinct Roles of Two Zebrafish AHR Repressors (AHRRa and AHRRb) in Embryonic Development and Regulating the Response to 2,3,7,8-Tetrachlorodibenzo-P-Dioxin. Toxicol. Sci. 2009, 110, 426–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, A.; Dubansky, B.; Bodinier, C.; Garcia, T.I.; Miles, S.; Pilley, C.; Raghunathan, V.; Roach, J.L.; Walker, N.; Walter, R.B.; et al. Genomic and Physiological Footprint of the Deepwater Horizon Oil Spill on Resident Marsh Fishes. Proc. Natl. Acad. Sci. USA 2012, 109, 20298–20302. [Google Scholar] [CrossRef] [Green Version]
- Tummers, B.; Green, D.R. Caspase-8: Regulating Life and Death. Immunol. Rev. 2017, 277, 76–89. [Google Scholar] [CrossRef] [Green Version]
- Pomierny, B.; Krzyżanowska, W.; Smaga, I.; Pomierny-Chamioło, L.; Stankowicz, P.; Budziszewska, B. Ethylene Glycol Ethers Induce Oxidative Stress in the Rat Brain. Neurotox. Res. 2014, 26, 422–429. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eliso, M.C.; Corsi, I.; Manfra, L.; Spagnuolo, A. Phenotypic and Gene Expression Profiles of Embryo Development of the Ascidian Ciona robusta Exposed to Dispersants. Water 2022, 14, 1539. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101539
Eliso MC, Corsi I, Manfra L, Spagnuolo A. Phenotypic and Gene Expression Profiles of Embryo Development of the Ascidian Ciona robusta Exposed to Dispersants. Water. 2022; 14(10):1539. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101539
Chicago/Turabian StyleEliso, Maria Concetta, Ilaria Corsi, Loredana Manfra, and Antonietta Spagnuolo. 2022. "Phenotypic and Gene Expression Profiles of Embryo Development of the Ascidian Ciona robusta Exposed to Dispersants" Water 14, no. 10: 1539. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101539