
Spartina alterniflora Invaded Coastal Wetlands by Raising Soil Sulfur Contents: A Meta-Analysis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
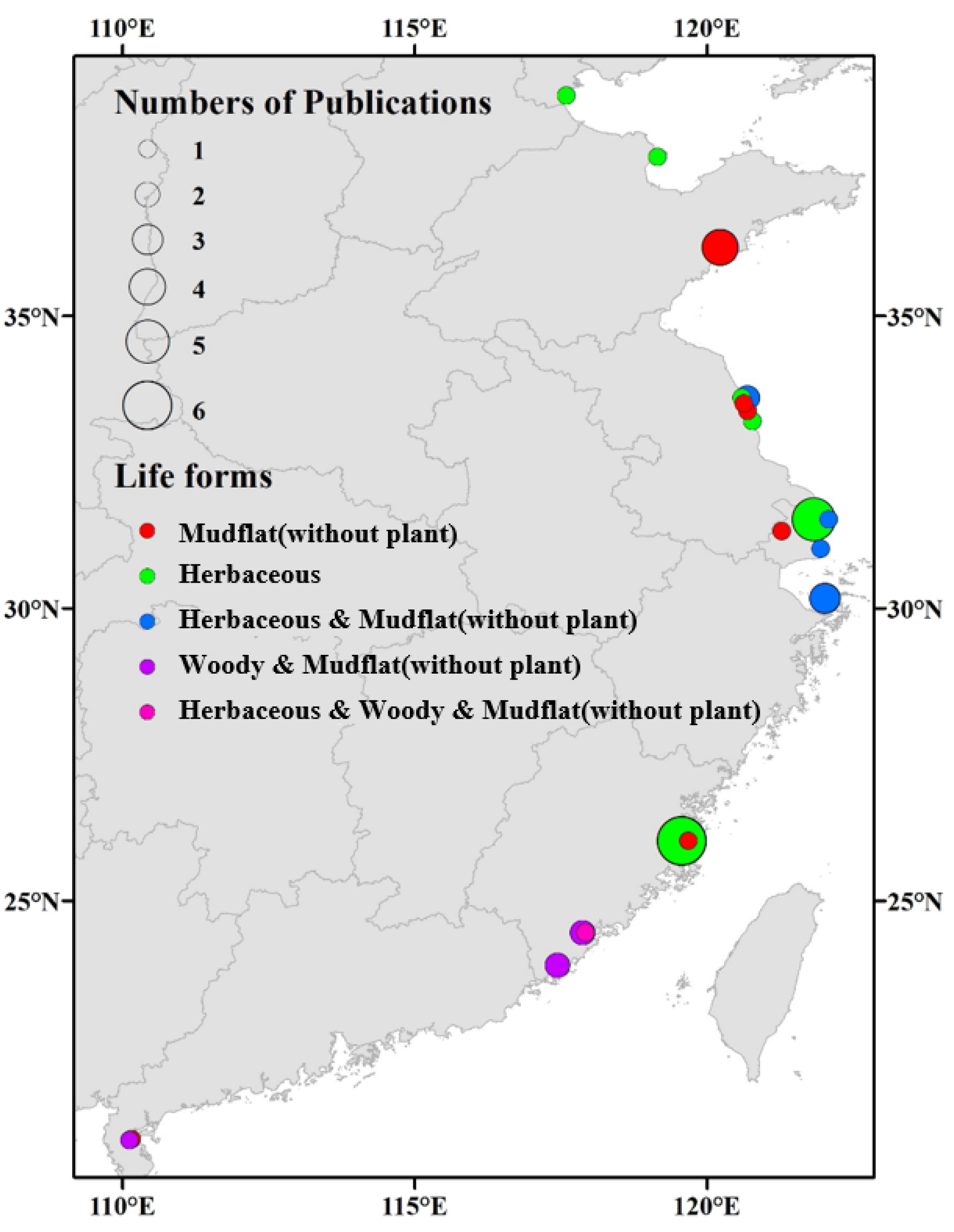
2.1. Literature Research and Data Collection
2.2. Statistical Analysis
3. Results
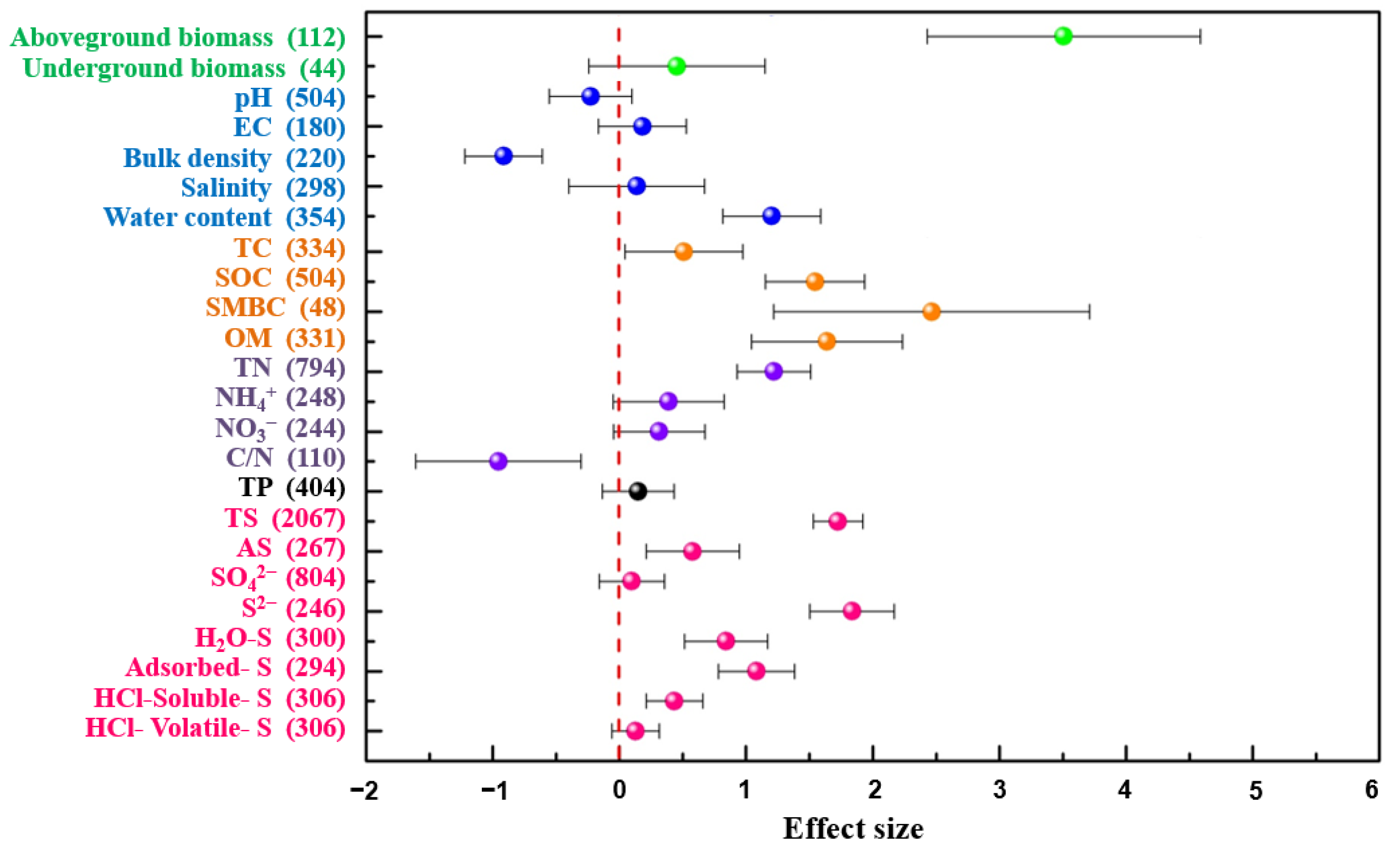
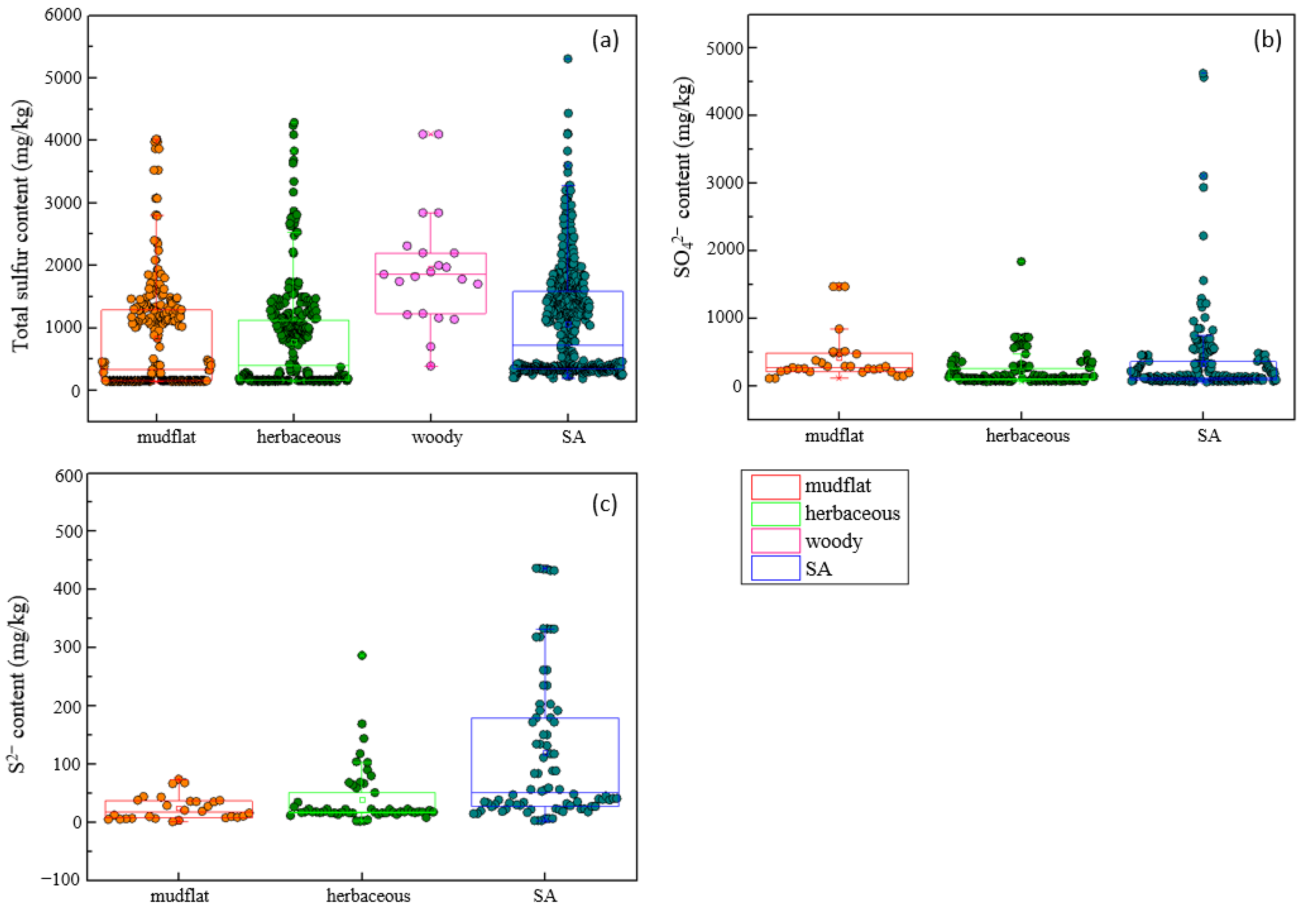
3.1. Effects of S. alterniflora Invasion on Soil Sulfur
3.2. Effects of S. alterniflora Invasion on Soil Physicochemical Properties
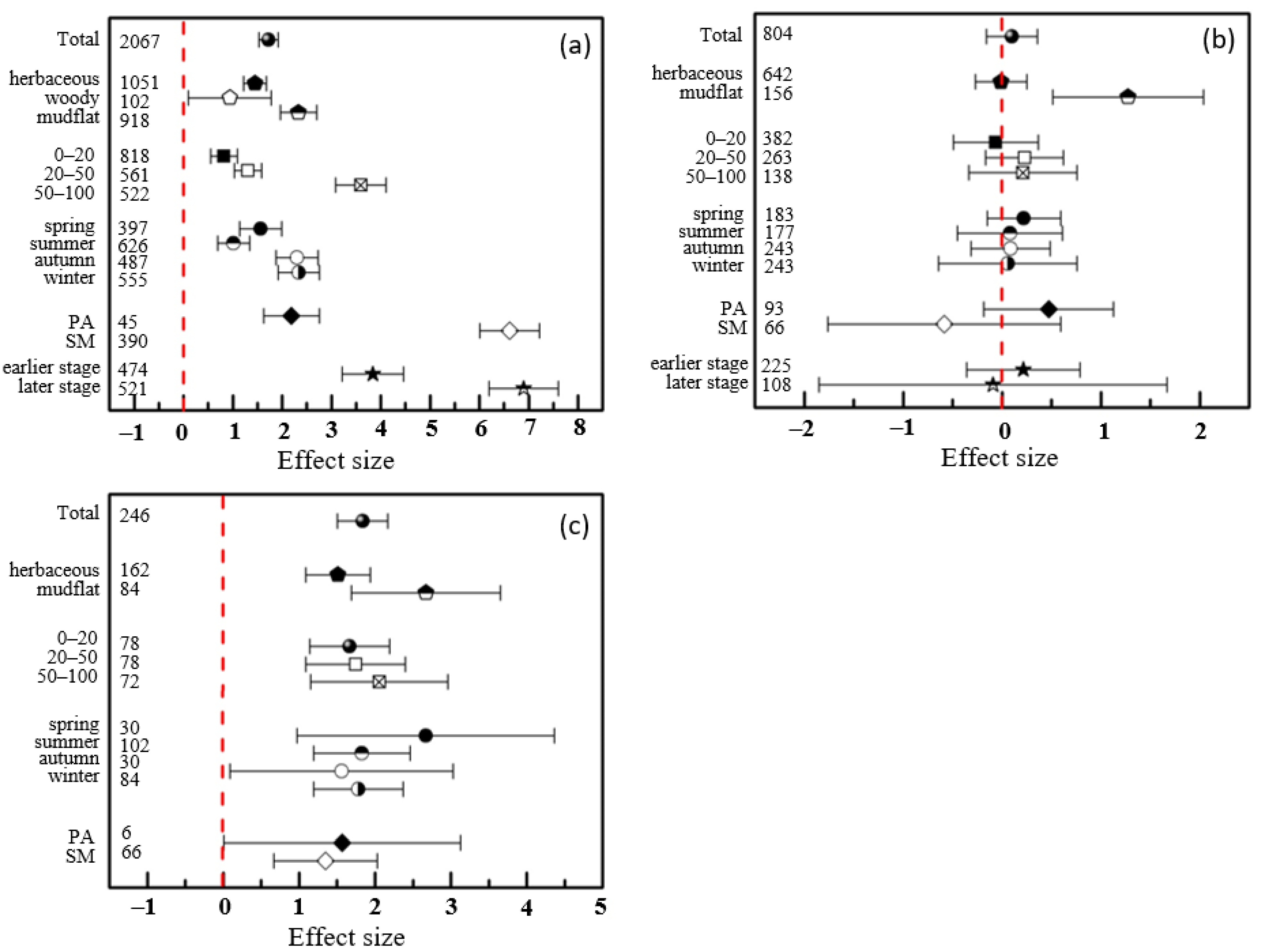
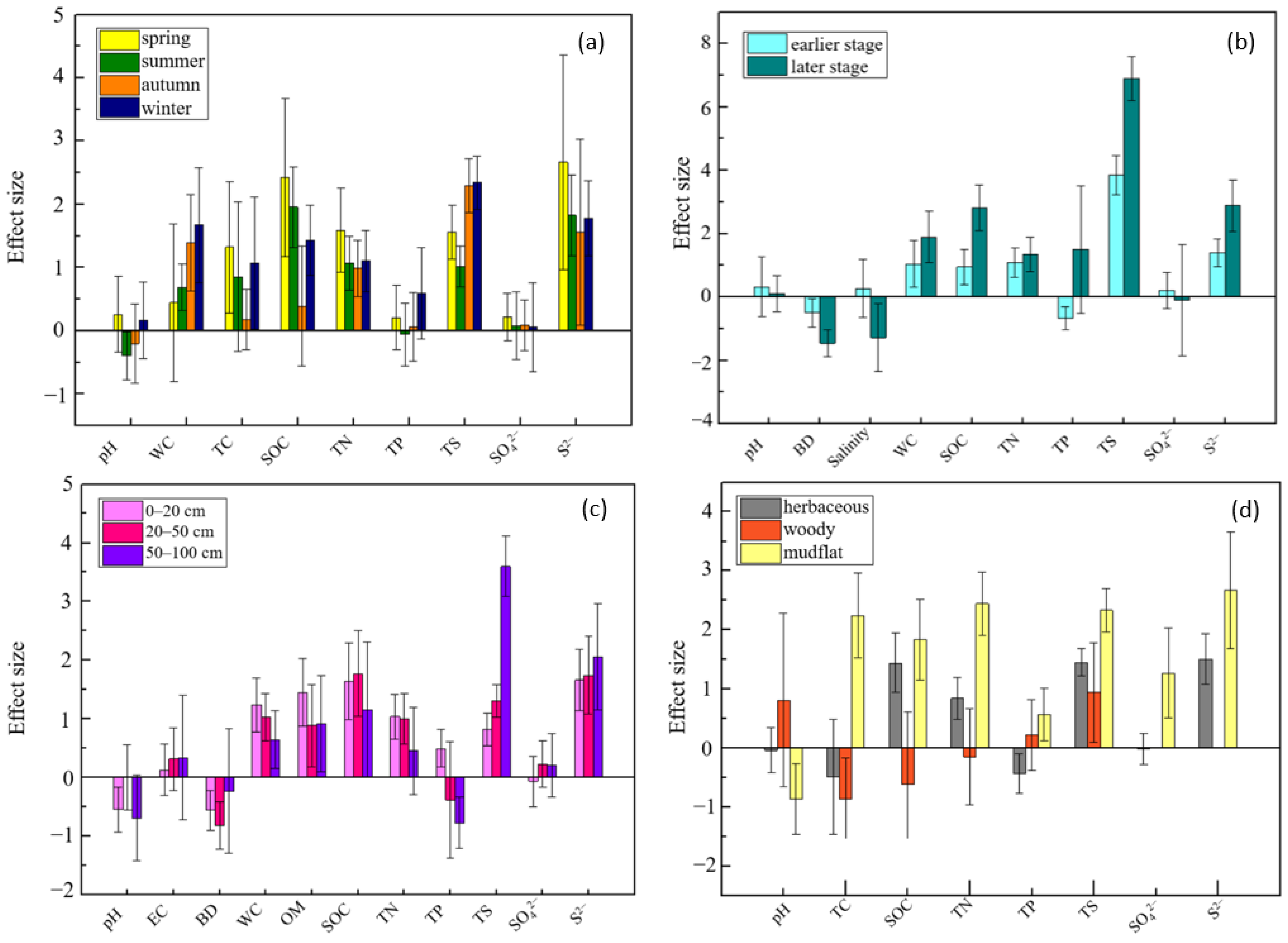
3.3. Effects Differed in Various Spatiotemporal Conditions and Life Forms of Native Species
4. Discussion
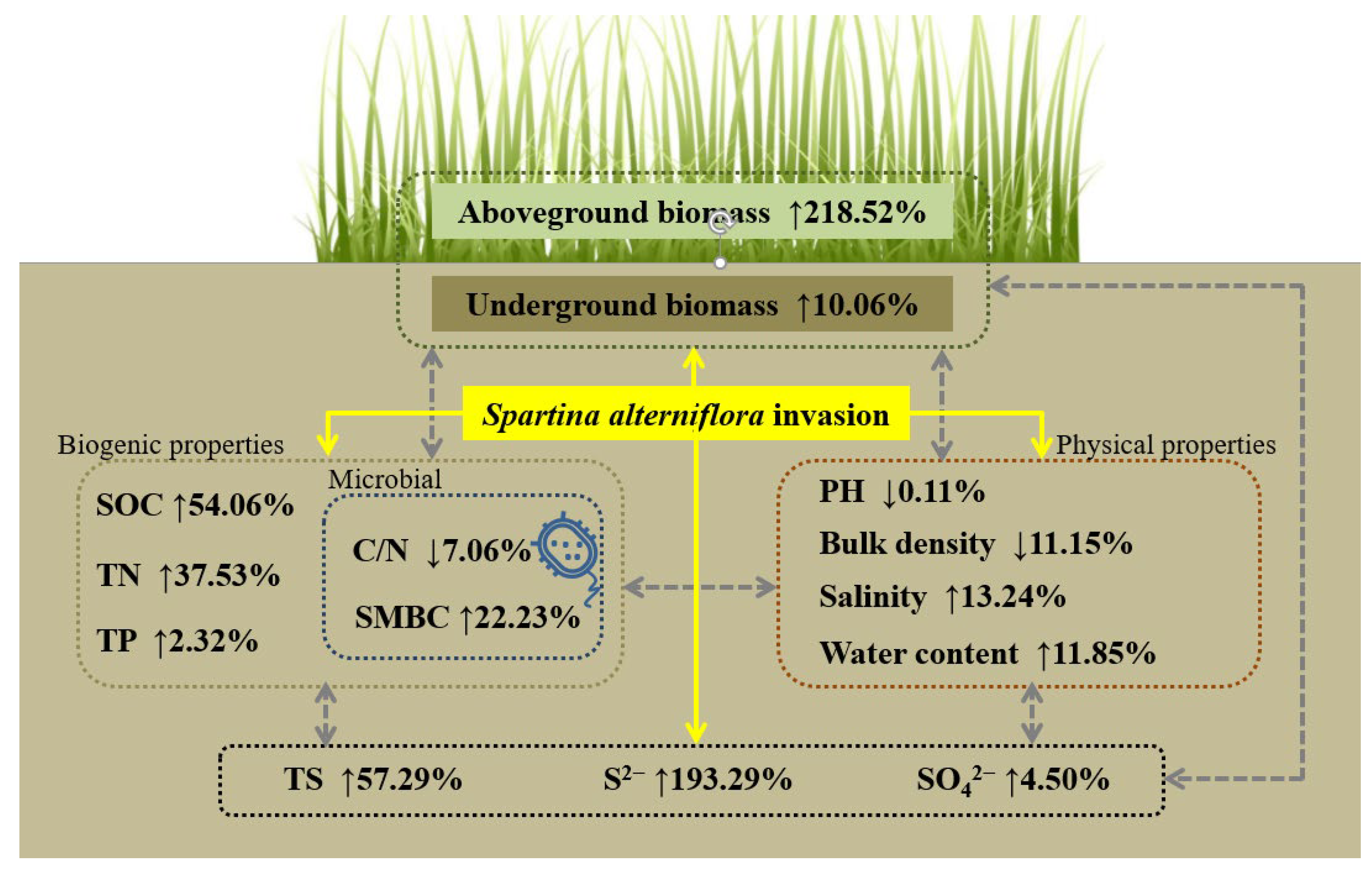
4.1. S. alterniflora Invasion Increased Soil Sulfur Contents
4.2. Invasive Plant Biomass Promotes Soil Nutrient Contents, Changes Physicochemical Properties, and Increases Soil S Contents
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Macreadie, P.I.; Anton, A.; Raven, J.A.; Beaumont, N.; Connolly, R.M.; Friess, D.A.; Kelleway, J.J.; Kennedy, H.; Kuwae, T.; Lavery, P.S.; et al. The future of Blue Carbon science. Nat. Commun. 2019, 10, 3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, J.L.; Mishra, D.R.; Alber, M.; Byrd, K.B. BERM: A Belowground Ecosystem Resiliency Model for estimating Spartina alterniflora belowground biomass. New Phytol. 2021, 232, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Sievers, M.; Brown, C.J.; Tulloch, V.J.D.; Pearson, R.M.; Haig, J.A.; Turschwell, M.P.; Connolly, R.M. The Role of Vegetated Coastal Wetlands for Marine Megafauna Conservation. Trends Ecol. Evol. 2019, 34, 807–817. [Google Scholar] [CrossRef]
- Latombe, G.; Richardson, D.M.; McGeoch, M.A.; Altwegg, R.; Catford, J.A.; Chase, J.M.; Courchamp, F.; Esler, K.J.; Jeschke, J.M.; Landi, P.; et al. Mechanistic reconciliation of community and invasion ecology. Ecosphere 2021, 12, e03359. [Google Scholar] [CrossRef]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.C.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of biodiversity loss on ocean ecosystem services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef] [Green Version]
- Wails, C.N.; Baker, K.; Blackburn, R.; Del Vallé, A.; Heise, J.; Herakovich, H.; Holthuijzen, W.A.; Nissenbaum, M.P.; Rankin, L.; Savage, K.; et al. Assessing changes to ecosystem structure and function following invasion by Spartina alterniflora and Phragmites australis: A meta-analysis. Biol. Invasions 2021, 23, 2695–2709. [Google Scholar] [CrossRef]
- Syed, S.; Xu, M.; Wang, Z.; Yu, C.; Lian, B. Invasive Spartina alterniflora in controlled cultivation: Environmental implications of converging future technologies. Ecol. Indic. 2021, 130, 108027. [Google Scholar] [CrossRef]
- Li, B.; Liao, C.H.; Zhang, X.D.; Chen, H.L.; Wang, Q.; Chen, Z.Y.; Gan, X.J.; Wu, J.H.; Zhao, B.; Ma, Z.J.; et al. Spartina alterniflora invasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects. Ecol. Eng. 2009, 35, 511–520. [Google Scholar] [CrossRef]
- Hui, C. Introduced species shape insular mutualistic networks. Proc. Natl. Acad. Sci. USA 2021, 118, e2026396118. [Google Scholar] [CrossRef]
- Mao, D.; Liu, M.; Wang, Z.; Li, L.; Man, W.; Jia, M.; Zhang, Y. Rapid invasion of Spartina alterniflora in the coastal zone of mainland China: Spatiotemporal patterns and human prevention. Sensors 2019, 19, 2308. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xiao, X.; Qiu, S.; Xu, X.; Wang, X.; Chang, Q.; Wu, J.; Li, B. Quantifying latitudinal variation in land surface phenology of Spartina alterniflora saltmarshes across coastal wetlands in China by Landsat 7/8 and Sentinel-2 images. Remote Sens. Environ. 2022, 269, 112810. [Google Scholar] [CrossRef]
- Gigolashvili, T.; Kopriva, S. Transporters in plant sulfur metabolism. Front. Plant Sci. 2014, 5, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, M.A.; Durham, B.P. Sulfur metabolites in the pelagic ocean. Nat. Rev. Microbiol. 2019, 17, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Blum, S.C.; Lehmann, J.; Solomon, D.; Caires, E.F.; Alleoni, L.R.F. Sulfur forms in organic substrates affecting S mineralization in soil. Geoderma 2013, 200, 156–164. [Google Scholar] [CrossRef]
- Jørgensen, B.B.; Findlay, A.J.; Pellerin, A. The biogeochemical sulfur cycle of marine sediments. Front. Microbiol. 2019, 10, 849. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, P.X.; Miao, Z.Q.; Qi, G.F.; Wang, Z.; Yuan, Y.; Ahmad, N.; Cao, M.J.; Hell, R.; Wirtz, M.; et al. SULTR3s function in chloroplast sulfate uptake and affect ABA biosynthesis and the stress response. Plant Physiol. 2019, 180, 593–604. [Google Scholar] [CrossRef]
- Bagarinao, T. Sulfide as an environmental factor and toxicant: Tolerance and adaptations in aquatic organisms. Aquat. Toxicol. 1992, 24, 21–62. [Google Scholar] [CrossRef]
- Raven, J.A.; Scrimgeour, C.M. The influence of anoxia on plants of saline habitats with special reference to the sulphur cycle. Ann. Bot. 1997, 79, 79–86. [Google Scholar] [CrossRef]
- Lamers, L.P.M.; Govers, L.L.; Janssen, I.C.J.M.; Geurts, J.J.M.; Van der Welle, M.E.W.; Van Katwijk, M.M.; Van der Heid, T.; Roelofs, J.G.M.; Smolders, A.J.P. Sulfide as a soil phytotoxin—A review. Front. Plant Sci. 2013, 4, 268. [Google Scholar] [CrossRef] [Green Version]
- Bradley, P.M.; Dunn, E.L. Effects of sulfide on the growth of three salt marsh halophytes of the southeastern United States. Am. J. Bot. 1989, 76, 1707–1713. [Google Scholar] [CrossRef]
- Carlson, P.R.; Forrest, J. Uptake of dissolved sulfide by Spartina alterniflora: Evidence from natural sulfur isotope abundance ratios. Science 1982, 216, 633–635. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Sun, Z.; Li, J.; Gao, H.; Wang, H.; Wang, J.; Chen, B. Distributions characteristics and influencing factors of inorganic sulfur forms in soil of Spartina alterniflora marsh and Cyperus malaccensis marsh in the Min River Estuary. Acta Sci. Circumstantiae 2017, 37, 4747–4756. [Google Scholar]
- Li, P.; Xie, W.; Wang, Z.; Yan, Q. Effects of Spartina alterniflora invasion on sulfur content temporal and spatial variation in tidal flat wetland of Jiaozhou Bay. Acta Sci. Circumstantiae 2019, 39, 870–879. [Google Scholar]
- Wang, Z.; Zhang, H.; He, C.; Liu, C.; Liang, X.; Chen, X. Spatiotemporal variability in soil sulfur storage is changed by exotic Spartina alterniflora in the Jiuduansha Wetland, China. Ecol. Eng. 2019, 133, 160–166. [Google Scholar] [CrossRef]
- He, T.; Sun, Z.; Li, J.; Gao, H.; Zhu, H.; Ren, P. Spatial Distribution of total sulfur contents in plant-soil systems of the typical marshes with different flooding regimes in the Min River Estuary. J. Soil Water Conserv. 2016, 30, 246–254. [Google Scholar]
- Xiao, S.; Luo, M.; Liu, Y.; Bai, J.; Yang, Y.; Zhai, Z.; Huang, J. Rhizosphere effect and its associated soil-microbe interactions drive iron fraction dynamics in tidal wetland soils. Sci. Total Environ. 2021, 756, 144056. [Google Scholar] [CrossRef]
- Zheng, Y.; Bu, N.S.; Long, X.E.; Sun, J.; He, C.Q.; Liu, X.Y.; Cui, J.; Liu, D.X.; Chen, X.P. Sulfate reducer and sulfur oxidizer respond differentially to the invasion of Spartina alterniflora in estuarine salt marsh of China. Ecol. Eng. 2017, 99, 182–190. [Google Scholar] [CrossRef]
- Valenzuela, E.I.; Prieto-Davó, A.; López-Lozano, N.E.; Hernández-Eligio, A.; Vega-Alvarado, L.; Juárez, K.; García-González, A.S.; López, M.G.; Cervantes, F.J. Anaerobic methane oxidation driven by microbial reduction of natural organic matter in a tropical wetland. Appl. Environ. Microbiol. 2017, 83, e00645-17. [Google Scholar] [CrossRef] [Green Version]
- Pallud, C.; Van Cappellen, P. Kinetics of microbial sulfate reduction in estuarine sediments. Geochim. Cosmochim. Acta 2006, 70, 1148–1162. [Google Scholar] [CrossRef]
- Pester, M.; Knorr, K.H.; Friedrich, M.W.; Wagner, M.; Loy, A. Sulfate-reducing microorganisms in wetlands—fameless actors in carbon cycling and climate change. Front. Microbiol. 2012, 3, 72. [Google Scholar] [CrossRef] [Green Version]
- Maricle, B.R.; Lee, R.W. Aerenchyma development and oxygen transport in the estuarine cordgrasses Spartina alterniflora and S. anglica. Aquat. Bot. 2002, 74, 109–120. [Google Scholar] [CrossRef]
- Wan, S.; Liu, X.; Mou, X.; Zhao, Y. Comparison of Carbon, Nitrogen, and Sulfur in Coastal Wetlands Dominated by Native and Invasive Plants in the Yancheng National Nature Reserve, China. Chin. Geogr. Sci. 2020, 30, 202–216. [Google Scholar] [CrossRef]
- Tong, X.; Sun, Z.; Zeng, A.; Chen, B.; Wang, H. Effects of Spartina alterniflora invasion in a seaward direction on variations of inorganic sulfur forms in marsh soils of the Minjiang River estuary, China. J. Appl. Ecol. 2019, 30, 3518–3526. [Google Scholar]
- Bai, S.; Li, J.; He, Z.; Van Nostrand, J.D.; Tian, Y.; Lin, G.; Zhou, J.; Zheng, T. GeoChip-based analysis of the functional gene diversity and metabolic potential of soil microbial communities of mangroves. Appl. Microbiol. Biotechnol. 2013, 97, 7035–7048. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.P.; Sun, J.; Wang, Y.; Zhang, H.Y.; He, C.Q.; Liu, X.Y.; Bu, N.S.; Long, X.E. Temporal and spatial impact of Spartina alterniflora invasion on methanogens community in Chongming Island, China. J. Microbiol. 2018, 56, 507–515. [Google Scholar] [CrossRef]
- Gao, F.L.; Wang, Y.H.; Whitt, A.A.; Wang, H.D.; Ma, C.C.; Guo, H.Y. Belowground responses of Phragmites australis and Suaeda salsa to salinity and water depth changes. Pakistan J. Bot. 2018, 50, 853–861. [Google Scholar]
- Wang, H.; Liao, G.; D’Souza, M.; Yu, X.; Yang, J.; Yang, X.; Zheng, T. Temporal and spatial variations of greenhouse gas fluxes from a tidal mangrove wetland in Southeast China. Environ. Sci. Pollut. Res. 2016, 23, 1873–1885. [Google Scholar] [CrossRef]
- He, C.; Cheng, L.; Wang, D.; Zhao, Z.; Wang, Z.; Wang, F.; Wang, X.; Zhang, P.; Chen, X.; Liu, X. Spartina alterniflora raised soil sulfide content by regulating sulfur cycle-associated bacteria in the Jiuduansha Wetland of China. Plant Soil 2021, 469, 107–121. [Google Scholar] [CrossRef]
- Xia, L.; Yang, W.; Zhao, H.; Xiao, Y.; Qing, H.; Zhou, C.; An, S. High Soil Sulfur Promotes Invasion of Exotic Spartina alterniflora into Native Phragmites australis Marsh. Clean Soil Air Water 2015, 43, 1666–1671. [Google Scholar] [CrossRef]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.T.; Rothstein, H.R. A basic introduction to fixed-effect and random-effects models for meta-analysis. Res. Synth. Methods 2010, 1, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Duval, S.; Tweedie, R. A Nonparametric “Trim and Fill” Method of Accounting for Publication Bias in Meta-Analysis. J. Am. Stat. Assoc. 2000, 95, 89–98. [Google Scholar]
- He, Q.; Bertness, M.D.; Altieri, A.H. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 2013, 16, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Maggi, E.; Benedetti-Cecchi, L.; Castelli, A.; Chatzinikolaou, E.; Crowe, T.P.; Ghedini, G.; Kotta, J.; Lyons, D.A.; Ravaglioli, C.; Rilov, G.; et al. Ecological impacts of invading seaweeds: A meta-analysis of their effects at different trophic levels. Divers. Distrib. 2015, 21, 1–12. [Google Scholar] [CrossRef]
- Davidson, I.C.; Cott, G.M.; Devaney, J.L.; Simkanin, C. Differential effects of biological invasions on coastal blue carbon: A global review and meta-analysis. Glob. Chang. Biol. 2018, 24, 5218–5230. [Google Scholar] [CrossRef]
- Koch, M.S.; Mendelssohn, I.A. Sulphide as a Soil Phytotoxin: Differential Responses in Two Marsh Species. J. Ecol. 1989, 77, 565–578. [Google Scholar] [CrossRef]
- Zeleke, J.; Sheng, Q.; Wang, J.G.; Huang, M.Y.; Xia, F.; Wu, J.H.; Quan, Z.X. Effects of Spartina alterniflora invasion on the communities of methanogens and sulfate-reducing bacteria in estuarine marsh sediments. Front. Microbiol. 2013, 4, 243. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wu, Y.; Kang, Q.; Zhang, J. Spatial variations of carbon, nitrogen, phosphorous and sulfur in the salt marsh sediments of the Yangtze Estuary in China. Estuar. Coast. Shelf Sci. 2007, 71, 47–59. [Google Scholar] [CrossRef]
- Xiao, D.; Zhang, L.; Zhu, Z. The range expansion patterns of Spartina alterniflora on salt marshes in the Yangtze Estuary, China. Estuar. Coast. Shelf Sci. 2010, 88, 99–104. [Google Scholar] [CrossRef]
- Zhang, P.; Li, B.; Wu, J.; Hu, S. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Chen, X.; Nie, M.; Fang, S.; Tang, B.; Quan, Z.; Li, B.; Fang, C. Effects of Spartina alterniflora Invasion on the Abundance, Diversity, and Community Structure of Sulfate Reducing Bacteria along a Successional Gradient of Coastal Salt Marshes in China. Wetlands 2017, 37, 221–232. [Google Scholar] [CrossRef]
- Bu, N.; Hu, Y.; Yang, X.; Zhang, X.; Wang, J.; Li, B.; Fang, C.; Song, Y. Effects of Spartina alterniflora invasion on soil physical and chemical properties in wetlands of the Yangtze River. Resour. Environ. Yangtze Basin 2017, 26, 100–109. [Google Scholar]
- Kraus, D.W.; Doeller, J.E. Oxidation of sulfide by Spartina alterniflora roots. Limnol. Oceanogr. 1999, 44, 1155–1159. [Google Scholar]
- Ferreira, T.O.; Otero, X.L.; Vidal-Torrado, P.; Macías, F. Effects of bioturbation by root and crab activity on iron and sulfur biogeochemistry in mangrove substrate. Geoderma 2007, 142, 36–46. [Google Scholar] [CrossRef]
- Cheng, X.; Luo, Y.; Chen, J.; Lin, G.; Chen, J.; Li, B. Short-term C4 plant Spartina alterniflora invasions change the soil carbon in C3 plant-dominated tidal wetlands on a growing estuarine Island. Soil Biol. Biochem. 2006, 38, 3380–3386. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, L.; Liu, N. C, N, and P Concentrations and Their Stoichiometry of Leaves and Roots with Different Life Forms in Hainan Province. J. Trop. Subtrop. Bot. 2020, 28, 131–135. [Google Scholar]
- Chen, Q.; Jian, S.; Ma, K.; Chen, P. Differences in macrobenthic faunal communities in mangrove wetland habitats (Zhanjiang, China) invaded and non-invaded by exotic cordgrass Spartina alterniflora. Ecol. Res. 2018, 33, 1113–1123. [Google Scholar] [CrossRef]
- Yu, X.; Yang, J.; Liu, L.; Tian, Y.; Yu, Z. Effects of Spartina alterniflora invasion on biogenic elements in a subtropical coastal mangrove wetland. Environ. Sci. Pollut. Res. 2015, 22, 3107–3115. [Google Scholar] [CrossRef]
- Hou, D.; He, D.; Hong, W.; You, W.; Wu, L.; Ji, Z.; Xiao, S. Influence of Invasive Spartina alterniflora on Soil Ecosystem in Coastal Wetland. Wetl. Sci Manag. 2015, 11, 67–72. [Google Scholar]
- Zhang, L.; Guo, Z.; Li, Z. Carbon storage and carbon sink of mangrove wetland: Research progress. J. Appl. Ecol. 2013, 24, 1153–1159. [Google Scholar]
- Varon-Lopez, M.; Dias, A.C.F.; Fasanella, C.C.; Durrer, A.; Melo, I.S.; Kuramae, E.E.; Andreote, F.D. Sulphur-oxidizing and sulphate-reducing communities in Brazilian mangrove sediments. Environ. Microbiol. 2014, 16, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, L.; Shi, C.; Lu, Y. Evaluating the relationship between the photochemical reflectance index and light use efficiency in a mangrove forest with Spartina alterniflora invasion. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 778–785. [Google Scholar] [CrossRef]
- Taketani, R.G.; Moitinho, M.A.; Mauchline, T.H.; Melo, I.S. Co-occurrence patterns of litter decomposing communities in mangroves indicate a robust community resistant to disturbances. PeerJ 2018, 6, e5710. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Luo, Y.; Jiang, L.; Zhou, X.; Wu, X.; Fang, C.; Chen, J.; Li, B. Invasion of Spartina alterniflora enhanced ecosystem carbon and nitrogen stocks in the Yangtze Estuary, China. Ecosystems 2007, 10, 1351–1361. [Google Scholar] [CrossRef]
- Zhang, Y.; Hua, Z.S.; Lu, H.; Oehmen, A.; Guo, J. Elucidating functional microorganisms and metabolic mechanisms in a novel engineered ecosystem integrating C, N, P and S biotransformation by metagenomics. Water Res. 2019, 148, 219–230. [Google Scholar] [CrossRef]
- Hu, N.; Li, H.; Tang, Z.; Li, Z.; Li, G.; Jiang, Y.; Hu, X.; Lou, Y. Community size, activity and C: N stoichiometry of soil microorganisms following reforestation in a Karst region. Eur. J. Soil Biol. 2016, 73, 77–83. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Luo, Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 3072. [Google Scholar] [CrossRef]
- Guimarães, R.M.L.; Ball, B.C.; Tormena, C.A.; Giarola, N.F.B.; da Silva, Á.P. Relating visual evaluation of soil structure to other physical properties in soils of contrasting texture and management. Soil Tillage Res. 2013, 127, 92–99. [Google Scholar] [CrossRef]
- Xu, S.; Lin, C.; Qiu, P.; Song, Y.; Yang, W.; Xu, G.; Feng, X.; Yang, Q.; Yang, X.; Niu, A. Tungsten- and cobalt-dominated heavy metal contamination of mangrove sediments in Shenzhen, China. Mar. Pollut. Bull. 2015, 100, 562–566. [Google Scholar] [CrossRef]
- Weaver, A.R.; Kissel, D.E.; Chen, F.; West, L.T.; Adkins, W.; Rickman, D.; Luvall, J.C. Mapping Soil pH Buffering Capacity of Selected Fields in the Coastal Plain. Soil Sci. Soc. Am. J. 2004, 68, 662–668. [Google Scholar] [CrossRef]
- Lu, Y.; Si, B.; Li, H.; Biswas, A. Elucidating controls of the variability of deep soil bulk density. Geoderma 2019, 348, 146–157. [Google Scholar] [CrossRef]
- Chi, Z.; Zhu, Y.; Li, H.; Wu, H.; Yan, B. Unraveling bacterial community structure and function and their links with natural salinity gradient in the Yellow River Delta. Sci. Total Environ. 2021, 773, 145673. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Bai, J.; Gao, Z.; Wang, J.; Zhao, Q. Effects of Water Level and Salinity on Total Sulfur Contents in Salt Marsh Soils of the Yellow River Delta, China. Wetlands 2016, 36, 137–143. [Google Scholar] [CrossRef]
- Zhao, K.; Song, J.; Feng, G.; Zhao, M.; Liu, J. Species, types, distribution, and economic potential of halophytes in China. Plant Soil 2011, 342, 495–509. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, Q.; Li, Y.; Li, X.; Wang, X. Inner-annual variation of soil water content and groundwater level in a typical islet wetland of Lake Poyang. J. Lake Sci. 2014, 26, 260–268. [Google Scholar]
- Jiang, L.F.; Luo, Y.Q.; Chen, J.K.; Li, B. Ecophysiological characteristics of invasive Spartina alterniflora and native species in salt marshes of Yangtze River estuary, China. Estuar. Coast. Shelf Sci. 2009, 81, 74–82. [Google Scholar] [CrossRef]
- Liao, C.Z.; Luo, Y.Q.; Fang, C.M.; Chen, J.K.; Li, B. Litter pool sizes, decomposition, and nitrogen dynamics in Spartina alterniflora-invaded and native coastal marshlands of the Yangtze Estuary. Oecologia 2008, 156, 589–600. [Google Scholar] [CrossRef]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zheng, H.; He, C.Q.; Xu, Q.Y.; Zhan, Y.W.; Lei, Y.R.; Du, W.; Yang, J.N. Allelopathic effects of invasive Spartina alterniflora root exudates in soil on the offspring (seeds) of Scirpus mariqueter. Allelopath. J. 2012, 29, 251–262. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Cheng, L.; He, C.; Wang, F.; Liu, J.; Li, Y.; Chen, X.; Liu, X.; Lv, G.; Wang, D. Spartina alterniflora Invaded Coastal Wetlands by Raising Soil Sulfur Contents: A Meta-Analysis. Water 2022, 14, 1633. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101633
Zhao Z, Cheng L, He C, Wang F, Liu J, Li Y, Chen X, Liu X, Lv G, Wang D. Spartina alterniflora Invaded Coastal Wetlands by Raising Soil Sulfur Contents: A Meta-Analysis. Water. 2022; 14(10):1633. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101633
Chicago/Turabian StyleZhao, Zhenzhen, Liyu Cheng, Chiquan He, Feifei Wang, Jialin Liu, Yuanhang Li, Xueping Chen, Xiaoyan Liu, Gaoming Lv, and Daoyuan Wang. 2022. "Spartina alterniflora Invaded Coastal Wetlands by Raising Soil Sulfur Contents: A Meta-Analysis" Water 14, no. 10: 1633. https://0-doi-org.brum.beds.ac.uk/10.3390/w14101633