
Decreased Photosynthetic Efficiency in Response to Site Translocation and Elevated Temperature Is Mitigated with LPS Exposure in Porites astreoides Symbionts

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reciprocal Transplant and Sample Collection
2.2. Temperature and LPS Experiment with Pulse Amplitude Modulation (PAM) Measurements
2.3. Data Analysis
3. Results
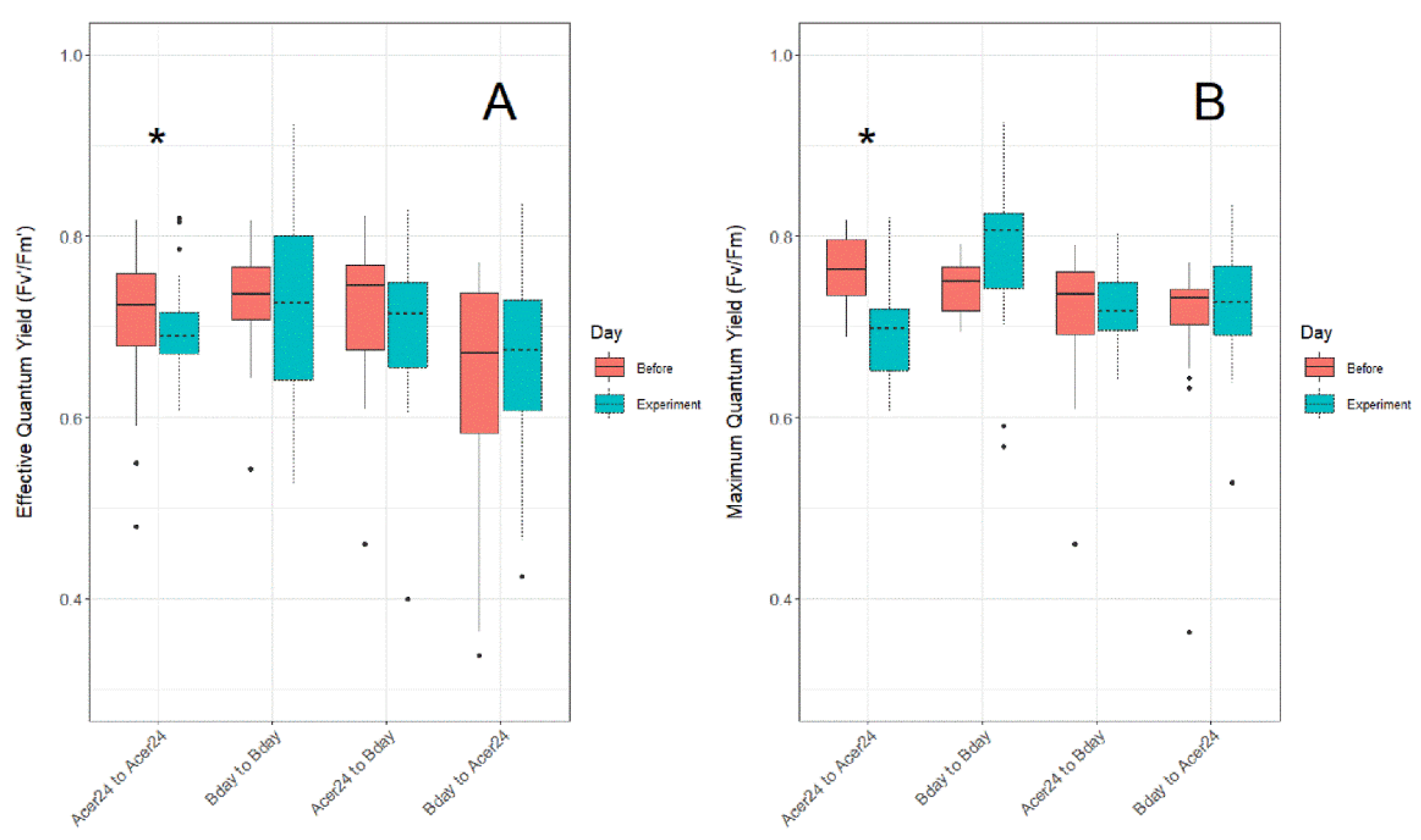
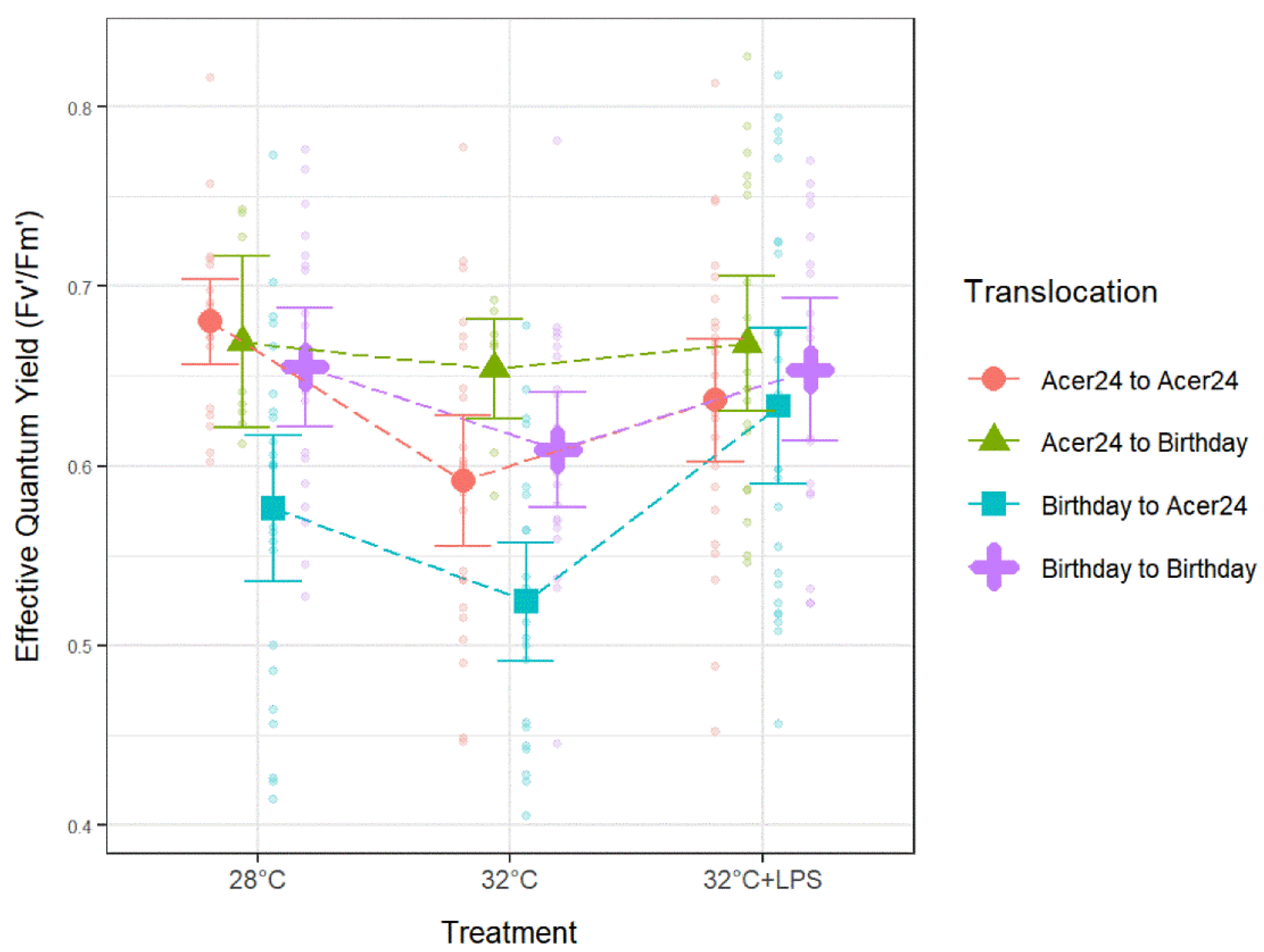
3.1. Quantum Yield (Fv′/Fm′)
3.2. Maximum Quantum Yield (Fv/Fm)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estimate | Std. Error | t Value | Pr (>|t|) | |
|---|---|---|---|---|
| (Intercept) | 2.16136 | 0.13455 | 16.063 | 2.00E-16 |
| Acer24 to Acer24.32 °C | 0.69441 | 0.20253 | 3.429 | 0.0007 |
| Acer24 to Acer24.32 °C + LPS | 0.30682 | 0.18915 | 1.622 | 0.1062 |
| Acer24 to Bday.28 °C | 0.07381 | 0.25633 | 0.288 | 0.7736 |
| Acer24 to Bday.32 °C | 0.17664 | 0.25173 | 0.702 | 0.4836 |
| Acer24 to Bday.32 °C + LPS | 0.07967 | 0.19055 | 0.418 | 0.6763 |
| Bday to Acer24.28 °C | 0.85011 | 0.20716 | 4.104 | 0.0001 |
| Bday to Acer24.32 °C | 1.47639 | 0.22908 | 6.445 | 6.76E-10 |
| Bday to Acer24.32 °C + LPS | 0.33192 | 0.18804 | 1.765 | 0.0789 |
| Bday to Bday.28 °C | 0.16985 | 0.19344 | 0.878 | 0.3808 |
| Bday to Bday.32 °C | 0.53492 | 0.20525 | 2.606 | 0.0098 |
| Bday to Bday.32 °C + LPS | 0.17928 | 0.19895 | 0.901 | 0.3685 |
| 1|FragmentTRUE | NA | NA | NA | NA |
| Estimate | Std. Error | t Value | Pr (>|t|) | |
|---|---|---|---|---|
| (Intercept) | 2.10589 | 0.09575 | 21.993 | 2.00E-16 |
| Acer24 to Acer24.32 °C | 0.13682 | 0.13254 | 1.032 | 0.3030 |
| Acer24 to Acer24.32 °C + LPS | −0.22421 | 0.12499 | −1.794 | 0.0742 |
| Acer24 to Bday.28 °C | −0.44417 | 0.15886 | −2.796 | 0.0056 |
| Acer24 to Bday.32 °C | −0.34692 | 0.15723 | −2.206 | 0.0283 |
| Acer24 to Bday.32 °C + LPS | −0.10313 | 0.13141 | −0.785 | 0.4334 |
| Bday to Acer24.28 °C | −0.14601 | 0.1245 | −1.173 | 0.2421 |
| Bday to Acer24.32 °C | 0.37786 | 0.13922 | 2.714 | 0.0071 |
| Bday to Acer24.32 °C + LPS | −0.02825 | 0.12681 | −0.223 | 0.8239 |
| Bday to Bday.28 °C | −0.47585 | 0.12415 | −3.833 | 0.0002 |
| Bday to Bday.32 °C | −0.14843 | 0.13185 | −1.126 | 0.2614 |
| Bday to Bday.32 °C + LPS | −0.27479 | 0.12747 | −2.156 | 0.0321 |
| 1|FragmentTRUE | NA | NA | NA | NA |
References
- Arto, I.; Dietzenbacher, E. Drivers of the growth in global greenhouse gas emissions. Environ. Sci. Technol. 2014, 48, 5388–5394. [Google Scholar] [CrossRef] [PubMed]
- Althor, G.; Watson, J.E.; Fuller, R.A. Global mismatch between greenhouse gas emissions and the burden of climate change. Sci. Rep. 2016, 6, 20281. [Google Scholar] [CrossRef] [PubMed]
- Wijffels, S.; Roemmich, D.; Monselesan, D.; Church, J.; Gilson, J. Ocean temperatures chronicle the ongoing warming of Earth. Nat. Clim. Chang. 2016, 6, 116–118. [Google Scholar] [CrossRef]
- Richmond, R.H. Coral reefs: Present problems and future concerns resulting from anthropogenic disturbance. Am. Zool. 1993, 33, 524–536. [Google Scholar] [CrossRef] [Green Version]
- Yonge, C.M.; Nicholls, A.G. Studies on the physiology of corals. IV. The structure, distribution, and physiology of the zooxanthellae. Sci. Rep. Great Barrier Reef Exped. 1928-29 1931, 1, 135–176. [Google Scholar]
- Brodersen, K.E.; Lichtenberg, M.; Ralph, P.J.; Kuhl, M.; Wangpraseurt, D. Radiative energy budget reveals high photosynthetic efficiency in symbiont-bearing corals. J. R. Soc. Interface 2014, 11, 20130997. [Google Scholar] [CrossRef]
- Mucsatine, L.; Porter, J.W. Reef corals: Mutualistic symbioses adapted to nutrient-poor environments. BioScience 1977, 27, 454–460. [Google Scholar]
- Roth, M.S. The engine of the reef: Photobiology of the coral-algal symbiosis. Front. Microbiol. 2014, 5, 422. [Google Scholar] [CrossRef]
- Lesser, M.P. Coral Bleaching: Causes and Mechanisms. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 405–419. [Google Scholar]
- Eakin, C.M.; Morgan, J.A.; Heron, S.F.; Smith, T.B.; Liu, G.; Alvarez-Filip, L.; Baca, B.; Bartels, E.; Bastidas, C.; Bouchon, C.; et al. Caribbean corals in crisis: Record thermal stress, bleaching, and mortality in 2005. PLoS ONE 2010, 5, e13969. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.A.; Bellchambers, L.M.; Depczynski, M.R.; Evans, R.D.; Evans, S.N.; Field, S.N.; Friedman, K.J.; Gilmour, J.P.; Holmes, T.H.; Middlebrook, R.; et al. Unprecedented mass bleaching and loss of coral across 12 degrees of latitude in western Australia in 2010–11. PLoS ONE 2012, 7, e51807. [Google Scholar] [CrossRef] [Green Version]
- Cervino, J.M.; Thompson, F.L.; Gomez-Gil, B.; Lorence, E.A.; Goreau, T.J.; Hayes, R.L.; Winiarski-Cervino, K.B.; Smith, G.W.; Hughen, K.; Bartels, E. The Vibrio core group induces yellow band disease in Caribbean and Indo-Pacific reef-building corals. J. Appl. Microbiol. 2008, 105, 1658–1671. [Google Scholar] [CrossRef] [PubMed]
- Gignoux-Wolfsohn, S.A.; Marks, C.J.; Vollmer, S.V. White band disease transmission in the threatened coral, Acropora cervicornis. Sci. Rep. 2012, 2, 804. [Google Scholar] [CrossRef] [PubMed]
- Aeby, G.S.; Ushijima, B.; Campbell, J.E.; Jones, S.; Williams, G.J.; Meyer, J.L.; Häse, C.; Paul, V.J. Pathogenesis of a tissue loss disease affecting multiple species of corals along the Florida reef tract. Front. Mar. Sci. 2019, 6, 678. [Google Scholar] [CrossRef]
- Cervino, J.M.; Hayes, R.; Goreau, T.J.; Smith, G.W. Zooxanthellae regulation in yellow blotch/band and other coral diseases contrasted with temperature related bleaching: In situ destruction vs. expulsion. Symbiosis 2004, 37, 63–85. [Google Scholar]
- Cervino, J.M.; Hayes, R.L.; Polson, S.W.; Polson, S.C.; Goreau, T.J.; Martinez, R.J.; Smith, G.W. Relationship of Vibrio species infection and elevated temperatures to yellow blotch/band disease in Caribbean corals. Appl. Environ. Microbiol. 2004, 70, 6855–6864. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.V.; Traylor-Knowles, N. Towards an integrated network of coral immune mechanisms. Proc. Biol. Sci. 2012, 279, 4106–4114. [Google Scholar] [CrossRef] [Green Version]
- Connelly, M.T.; McRae, C.J.; Liu, P.J.; Traylor-Knowles, N. Lipopolysaccharide treatment stimulates Pocillopora coral genotype-specific immune responses but does not alter coral-associated bacteria communities. Dev. Comp. Immunol. 2020, 109, 103717. [Google Scholar] [CrossRef]
- Mydlarz, L.D.; McGinty, E.S.; Harvell, C.D. What are the physiological and immunological responses of coral to climate warming and disease? J. Exp. Biol. 2010, 213, 934–945. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, K.M.; Gilmore, T.D. Innate immunity and cnidarian-Symbiodiniaceae mutualism. Dev. Comp. Immunol. 2019, 90, 199–209. [Google Scholar] [CrossRef]
- Kvennefors, E.C.; Leggat, W.; Kerr, C.C.; Ainsworth, T.D.; Hoegh-Guldberg, O.; Barnes, A.C. Analysis of evolutionarily conserved innate immune components in coral links immunity and symbiosis. Dev. Comp. Immunol. 2010, 34, 1219–1229. [Google Scholar] [CrossRef]
- Palmer, C.V.; Bythell, J.C.; Willis, B.L. Levels of immunity parameters underpin bleaching and disease susceptibility of reef corals. FASEB J. 2010, 24, 1935–1946. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.C.; Muller, E.M.; van Woesik, R. Coral immunology and resistance to disease. Dis. Aquat. Organ. 2010, 90, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Water, J.; Chaib De Mares, M.; Dixon, G.B.; Raina, J.B.; Willis, B.L.; Bourne, D.G.; van Oppen, M.J.H. Antimicrobial and stress responses to increased temperature and bacterial pathogen challenge in the holobiont of a reef-building coral. Mol. Ecol. 2018, 27, 1065–1080. [Google Scholar] [CrossRef]
- Young, B.D.; Serrano, X.M.; Rosales, S.M.; Miller, M.W.; Williams, D.; Traylor-Knowles, N. Innate immune gene expression in Acropora palmata is consistent despite variance in yearly disease events. PLoS ONE 2020, 15, e0228514. [Google Scholar] [CrossRef]
- Burns, J.H.; Gregg, T.M.; Takabayashi, M. Does coral disease affect symbiodinium? Investigating the impacts of growth anomaly on symbiont photophysiology. PLoS ONE 2013, 8, e72466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, T.; Hawkins, T.D.; Becker, S.; Pontasch, S.; Dove, S.; Hoegh-Guldberg, O.; Leggat, W.; Fisher, P.L.; Davy, S.K. Differential coral bleaching contrasting the activity and response of enzymatic antioxidants in symbiotic partners under thermal stress. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2015, 190, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barshis, D.J.; Ladner, J.T.; Oliver, T.A.; Seneca, F.O.; Traylor-Knowles, N.; Palumbi, S.R. Genomic basis for coral resilience to climate change. Proc. Natl. Acad. Sci. USA 2013, 110, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Palumbi, S.R.; Barshis, D.J.; Traylor-Knowles, N.; Bay, R.A. Mechanisms of reef coral resistance to future climate change. Science 2014, 344, 895–898. [Google Scholar] [CrossRef]
- Barshis, D.J.; Ladner, J.T.; Oliver, T.A.; Palumbi, S.R. Lineage-specific transcriptional profiles of Symbiodinium spp. unaltered by heat stress in a coral host. Mol. Biol. Evol. 2014, 31, 1343–1352. [Google Scholar] [CrossRef] [Green Version]
- Klepac, C.N.; Barshis, D.J. Reduced thermal tolerance of massive coral species in a highly variable environment. Proc. Biol. Sci. 2020, 287, 20201379. [Google Scholar] [CrossRef]
- Salas, B.H.; Haslun, J.A.; Strychar, K.B.; Ostrom, P.H.; Cervino, J.M. Site-specific variation in gene expression from Symbiodinium spp. associated with offshore and inshore Porites astreoides in the lower Florida Keys is lost with bleaching and disease stress. PLoS ONE 2017, 12, e0173350. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.K.; Lundgren, P.; Lutz, A.; Raina, J.B.; Howells, E.J.; Paley, S.A.; Willis, B.L.; van Oppen, M.J.H. Genetic markers for antioxidant capacity in a reef-building coral. Sci. Adv. 2016, 2, e1500842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drury, C.; Lirman, D. Genotype by environment interactions in coral bleaching. Proc. Biol. Sci. 2021, 288, 20210177. [Google Scholar] [CrossRef] [PubMed]
- Barott, K.L.; Huffmyer, A.S.; Davidson, J.M.; Lenz, E.A.; Matsuda, S.B.; Hancock, J.R.; Innis, T.; Drury, C.; Putnam, H.M.; Gates, R.D. Coral bleaching response is unaltered following acclimatization to reefs with distinct environmental conditions. Proc. Natl. Acad. Sci. USA 2021, 118, e2025435118. [Google Scholar] [CrossRef]
- Hauff, B.; Haslun, J.A.; Strychar, K.B.; Ostrom, P.H.; Cervino, J.M. Symbiont diversity of zooxanthellae (Symbiodinium spp.) in Porites astreoides and Montastraea cavernosa from a reciprocal transplant in the lower Florida Keys. Int. J. Biol. 2016, 8, 9–22. [Google Scholar] [CrossRef]
- Haslun, J.; Hauff, B.; Strychar, K.; Cervino, J. Decoupled seasonal stress as an indication of chronic stress in Montastraea cavernosa and Porites astreoides inhabiting the Florida reef tract. Int. J. Mar. Sci. 2016, 6, 1–20. [Google Scholar] [CrossRef]
- Bhagooli, R.; Hidaka, M. Photoinhibition, bleaching susceptibility and mortality in two scleractinian corals, Platygyra ryukyuensis and Stylophora pistillata, in response to thermal and light stresses. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2003, 137, 547–555. [Google Scholar] [CrossRef]
- Silverstein, R.N.; Cunning, R.; Baker, A.C. Tenacious D: Symbiodinium in clade D remain in reef corals at both high and low temperature extremes despite impairment. J. Exp. Biol. 2017, 220, 1192–1196. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.V.; McGinty, E.S.; Cummings, D.J.; Smith, S.M.; Bartels, E.; Mydlarz, L.D. Patterns of coral ecological immunology: Variation in the responses of Caribbean corals to elevated temperature and a pathogen elicitor. J. Exp. Biol. 2011, 214, 4240–4249. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio: Boston, MA, USA, 2021. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Ware, M.; Garfield, E.N.; Nedimyer, K.; Levy, J.; Kaufman, L.; Precht, W.; Winters, R.S.; Miller, S.L. Survivorship and growth in staghorn coral (Acropora cervicornis) outplanting projects in the Florida Keys National Marine Sanctuary. PLoS ONE 2020, 15, e0231817. [Google Scholar] [CrossRef] [PubMed]
- Muller, E.M.; Bartels, E.; Baums, I.B. Bleaching causes loss of disease resistance within the threatened coral species Acropora cervicornis. eLife 2018, 7, e35066. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.A.; McDougall, K.E.; Woodley, C.M.; Fauth, J.E.; Richmond, R.H.; Kushmaro, A.; Gibb, S.W.; Loya, Y.; Ostrander, G.K.; Kramarsky-Winter, E. Heat-stress and light-stress induce different cellular pathologies in the symbiotic dinoflagellate during coral bleaching. PLoS ONE 2013, 8, e77173. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, I.M.; Kuhl, M.; Larkum, A.W.D.; Ralph, P.J. Heat budget and thermal microenvironment of shallow-water corals: Do massive corals get warmer than branching corals? Limnol. Oceanogr. 2008, 53, 1548–1561. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, I.M.; Kuhl, M.; Larkum, A.W.; Ralph, P.J. Effects of flow and colony morphology on the thermal boundary layer of corals. J. R. Soc. Interface 2011, 8, 1785–1795. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Dubinsky, Z. Light-shade adaptation of Stylophora pistillata, a hermatypic coral from the Gulf of Eilat. Nature 1981, 289, 172–174. [Google Scholar] [CrossRef]
- Dove, S.G.; Lovell, C.; Fine, M.; Deckenback, J.; Hoegh-Guldberg, O.; Iglesias-Prieto, R.; Anthony, K.R. Host pigments: Potential facilitators of photosynthesis in coral symbioses. Plant Cell Environ. 2008, 31, 1523–1533. [Google Scholar] [CrossRef]
- Smith, E.G.; D’Angelo, C.; Salih, A.; Wiedenmann, J. Screening by coral green fluorescent protein (GFP)-like chromoproteins supports a role in photoprotection of zooxanthellae. Coral Reefs 2013, 32, 463–474. [Google Scholar] [CrossRef] [Green Version]
- Ralph, P.J.; Larkum, A.W.D.; Kühl, M. Temporal patterns in effective quantum yield of individual zooxanthellae expelled during bleaching. J. Exp. Mar. Biol. Ecol. 2005, 316, 17–28. [Google Scholar] [CrossRef]
- Downs, C.A.; Fauth, J.E.; Halas, J.C.; Dustan, P.; Bemiss, J.; Woodley, C.M. Oxidative stress and seasonal coral bleaching. Free Radic. Biol. Med. 2002, 33, 533–543. [Google Scholar] [CrossRef]
- McGinty, E.S.; Pieczonka, J.; Mydlarz, L.D. Variations in reactive oxygen release and antioxidant activity in multiple Symbiodinium types in response to elevated temperature. Microb. Ecol. 2012, 64, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Roberty, S.; Furla, P.; Plumier, J.C. Differential antioxidant response between two Symbiodinium species from contrasting environments. Plant Cell Environ. 2016, 39, 2713–2724. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.A.; Petrou, K.; Gates, R.D. Coral bleaching from a single cell perspective. ISME J. 2018, 12, 1558–1567. [Google Scholar] [CrossRef] [PubMed]
- Richier, S.; Furla, P.; Plantivaux, A.; Merle, P.L.; Allemand, D. Symbiosis-induced adaptation to oxidative stress. J. Exp. Biol. 2004, 208 Pt 2, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Gardner, S.G.; Raina, J.B.; Nitschke, M.R.; Nielsen, D.A.; Stat, M.; Motti, C.A.; Ralph, P.J.; Petrou, K. A multi-trait systems approach reveals a response cascade to bleaching in corals. BMC Biol. 2017, 15, 117. [Google Scholar] [CrossRef]
- Edreva, A. Generation and scavenging of reactive oxygen species in chloroplasts: A submolecular approach. Agric. Ecosyst. Environ. 2005, 106, 119–133. [Google Scholar] [CrossRef]
- Kristiansen, K.A.; Jensen, P.E.; Moller, I.M.; Schulz, A. Monitoring reactive oxygen species formation and localization in living cells by use of the fluorescent probe CM-H2DCFDA and confocal laser microscopy. Physiol. Plant 2009, 136, 369–383. [Google Scholar] [CrossRef]
- Lesser, M.P. Phylogenetic signature of light and thermal stress for the endosymbiotic dinoflagellates of corals (Family Symbiodiniaceae). Limnol. Oceanogr. 2019, 64, 1852–1863. [Google Scholar] [CrossRef]
- Tchernov, D.; Gorbunov, M.Y.; de Vargas, C.; Yadav, S.N.; Milligan, A.J.; Haggblom, M.; Falkowski, P.G. Membrane lipids of symbiotic algae are diagnostic of sensitivity to thermal bleaching in corals. Proc. Natl. Acad. Sci. USA 2004, 101, 13531–13535. [Google Scholar] [CrossRef] [Green Version]
- Wietherger, A.; Starzak, D.E.; Gould, K.S.; Davy, S.K. Differential ROS generation in response to stress in Symbiodinium spp. Biol. Bull. 2018, 234, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ayalon, I.; de Barros Marangoni, L.F.; Benichou, J.I.C.; Avisar, D.; Levy, O. Red Sea corals under artificial light pollution at night (ALAN) undergo oxidative stress and photosynthetic impairment. Glob. Chang. Biol. 2019, 25, 4194–4207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, A.H.; Bhagooli, R.; Ralph, P.J.; Takahashi, S. Coral bleaching: The role of the host. Trends Ecol. Evol. 2009, 24, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Haslun, J.A.; Salas, B.H.; Strychar, K.B.; Ostrom, N.E.; Cervino, J.M. Biotic stress contributes to seawater temperature induced stress in a site-specific manner for Porites astreoides. Mar. Biol. 2018, 165, 160. [Google Scholar] [CrossRef]
- Haslun, J.A.; Salas, B.H.; Strychar, K.B.; Cervino, J.M.; Ostrom, N.E. Variation in immune-related gene expression provides evidence of local adaptation in Porites astreoides (Lamarck, 1816) between inshore and offshore meta-populations inhabiting the lower Florida reef tract, USA. Water 2021, 13, 2107. [Google Scholar] [CrossRef]





| Contrast | Estimate | SE | df | z Ratio | p Value |
|---|---|---|---|---|---|
| Acer to Acer.28 °C–Acer to Acer.32 °C | −0.69441 | 0.203 | Inf | −3.429 | 0.0006 |
| Acer to Acer.28 °C–(Acer to Acer.32 °C + LPS) | −0.30682 | 0.189 | Inf | −1.622 | 0.1048 |
| Acer to Acer.28 °C–Acer to Birthday.28 °C | −0.07381 | 0.256 | Inf | −0.288 | 0.7734 |
| Acer to Acer.28 °C–Birthday to Acer.28 °C | −0.85011 | 0.207 | Inf | −4.104 | 0.0001 |
| Acer to Acer.28 °C–Birthday to Birthday.28 °C | −0.16985 | 0.193 | Inf | −0.878 | 0.3799 |
| Acer to Acer.32 °C–(Acer to Acer.32 °C + LPS) | 0.3876 | 0.201 | Inf | 1.924 | 0.0544 |
| Acer to Acer.32 °C–Acer to Birthday.32 °C | 0.51777 | 0.261 | Inf | 1.983 | 0.0474 |
| Acer to Acer.32 °C–Birthday to Acer.32 °C | −0.78198 | 0.239 | Inf | −3.267 | 0.0011 |
| Acer to Acer.32 °C–Birthday to Birthday.32 °C | 0.15949 | 0.217 | Inf | 0.736 | 0.4616 |
| (Acer to Acer.32 °C + LPS)–(Acer to Birthday.32 °C + LPS) | 0.22715 | 0.189 | Inf | 1.199 | 0.2304 |
| (Acer to Acer.32 °C + LPS)–(Birthday to Acer.32 °C + LPS) | −0.0251 | 0.187 | Inf | −0.134 | 0.8932 |
| (Acer to Acer.32 °C + LPS)–(Birthday to Birthday.32 °C + LPS) | 0.12754 | 0.198 | Inf | 0.645 | 0.5192 |
| Acer to Birthday.28 °C–Acer to Birthday.32 °C | −0.10283 | 0.305 | Inf | −0.337 | 0.7358 |
| Acer to Birthday.28 °C–(Acer to Birthday.32 °C + LPS) | −0.00586 | 0.257 | Inf | −0.023 | 0.9818 |
| Acer to Birthday.28 °C–Birthday to Acer.28 °C | −0.7763 | 0.269 | Inf | −2.885 | 0.0039 |
| Acer to Birthday.28 °C–Birthday to Birthday.28 °C | −0.09604 | 0.259 | Inf | −0.371 | 0.7104 |
| Acer to Birthday.32 °C–(Acer to Birthday.32 °C + LPS) | 0.09697 | 0.252 | Inf | 0.385 | 0.7003 |
| Acer to Birthday.32 °C–Birthday to Acer.32 °C | −1.29975 | 0.282 | Inf | −4.606 | 0.0001 |
| Acer to Birthday.32 °C–Birthday to Birthday.32 °C | −0.35828 | 0.263 | Inf | −1.361 | 0.1735 |
| (Acer to Birthday.32 °C + LPS)–(Birthday to Acer.32 °C + LPS) | −0.25225 | 0.188 | Inf | −1.34 | 0.1804 |
| (Acer to Birthday.32 °C + LPS)–(Birthday to Birthday.32 °C + LPS) | −0.09961 | 0.199 | Inf | −0.5 | 0.617 |
| Birthday to Acer.28 °C–Birthday to Acer.32 °C | −0.62628 | 0.243 | Inf | −2.574 | 0.01 |
| Birthday to Acer.28 °C–(Birthday to Acer.32 °C + LPS) | 0.51819 | 0.205 | Inf | 2.527 | 0.0115 |
| Birthday to Acer.28 °C–Birthday to Birthday.28 °C | 0.68026 | 0.21 | Inf | 3.238 | 0.0012 |
| Birthday to Acer.32 °C–(Birthday to Acer.32 °C + LPS) | 1.14448 | 0.227 | Inf | 5.037 | 0.0001 |
| Birthday to Acer.32 °C–Birthday to Birthday.32 °C | 0.94147 | 0.242 | Inf | 3.896 | 0.0001 |
| (Birthday to Acer.32 °C + LPS)–(Birthday to Birthday.32 °C + LPS) | 0.15264 | 0.197 | Inf | 0.776 | 0.438 |
| Birthday to Birthday.28 °C–Birthday to Birthday.32 °C | −0.36508 | 0.208 | Inf | −1.754 | 0.0795 |
| Birthday to Birthday.28 °C–(Birthday to Birthday.32 °C + LPS) | −0.00943 | 0.202 | Inf | −0.047 | 0.9628 |
| Birthday to Birthday.32 °C–(Birthday to Birthday.32 °C + LPS) | 0.35565 | 0.213 | Inf | 1.667 | 0.0955 |
| Contrast | Estimate | SE | df | z Ratio | p Value |
|---|---|---|---|---|---|
| Acer to Acer.28 °C–Acer to Acer.32 °C | −0.13682 | 0.133 | Inf | −1.032 | 0.3019 |
| Acer to Acer.28 °C–(Acer to Acer.32 °C + LPS) | 0.22421 | 0.125 | Inf | 1.794 | 0.0728 |
| Acer to Acer.28 °C–Acer to Birthday.28 °C | 0.44417 | 0.159 | Inf | 2.796 | 0.0052 |
| Acer to Acer.28 °C–Birthday to Acer.28 °C | 0.14601 | 0.125 | Inf | 1.173 | 0.2409 |
| Acer to Acer.28 °C–Birthday to Birthday.28 °C | 0.47585 | 0.124 | Inf | 3.833 | 0.0001 |
| Acer to Acer.32 °C–(Acer to Acer.32 °C + LPS) | 0.36103 | 0.122 | Inf | 2.963 | 0.0031 |
| Acer to Acer.32 °C–Acer to Birthday.32 °C | 0.48374 | 0.155 | Inf | 3.126 | 0.0018 |
| Acer to Acer.32 °C–Birthday to Acer.32 °C | −0.24104 | 0.136 | Inf | −1.767 | 0.0772 |
| Acer to Acer.32 °C–Birthday to Birthday.32 °C | 0.28525 | 0.129 | Inf | 2.213 | 0.0269 |
| (Acer to Acer.32 °C + LPS)–(Acer to Birthday.32 °C + LPS) | −0.12109 | 0.121 | Inf | −1.004 | 0.3155 |
| (Acer to Acer.32 °C + LPS)–(Birthday to Acer.32 °C + LPS) | −0.19596 | 0.116 | Inf | −1.695 | 0.0901 |
| (Acer to Acer.32 °C + LPS)–(Birthday to Birthday.32 °C + LPS) | 0.05058 | 0.116 | Inf | 0.435 | 0.6637 |
| Acer to Birthday.28 °C–Acer to Birthday.32 °C | −0.09725 | 0.178 | Inf | −0.547 | 0.5844 |
| Acer to Birthday.28 °C–(Acer to Birthday.32 °C + LPS) | −0.34105 | 0.155 | Inf | −2.194 | 0.0282 |
| Acer to Birthday.28 °C–Birthday to Acer.28 °C | −0.29816 | 0.15 | Inf | −1.992 | 0.0463 |
| Acer to Birthday.28 °C–Birthday to Birthday.28 °C | 0.03167 | 0.149 | Inf | 0.212 | 0.8321 |
| Acer to Birthday.32 °C–(Acer to Birthday.32 °C + LPS) | −0.2438 | 0.154 | Inf | −1.585 | 0.1129 |
| Acer to Birthday.32 °C–Birthday to Acer.32 °C | −0.72478 | 0.161 | Inf | −4.515 | 0.0001 |
| Acer to Birthday.32 °C–Birthday to Birthday.32 °C | −0.19849 | 0.154 | Inf | −1.287 | 0.198 |
| (Acer to Birthday.32 °C + LPS)–(Birthday to Acer.32 °C + LPS) | −0.07488 | 0.123 | Inf | −0.611 | 0.5411 |
| (Acer to Birthday.32 °C + LPS)–(Birthday to Birthday.32 °C + LPS) | 0.17167 | 0.123 | Inf | 1.393 | 0.1635 |
| Birthday to Acer.28 °C–Birthday to Acer.32 °C | −0.52387 | 0.129 | Inf | −4.073 | 0.0001 |
| Birthday to Acer.28 °C–(Birthday to Acer.32 °C + LPS) | −0.11776 | 0.115 | Inf | −1.023 | 0.3062 |
| Birthday to Acer.28 °C–Birthday to Birthday.28 °C | 0.32984 | 0.112 | Inf | 2.941 | 0.0033 |
| Birthday to Acer.32 °C–(Birthday to Acer.32 °C + LPS) | 0.40611 | 0.131 | Inf | 3.103 | 0.0019 |
| Birthday to Acer.32 °C–Birthday to Birthday.32 °C | 0.52629 | 0.136 | Inf | 3.877 | 0.0001 |
| (Birthday to Acer.32 °C + LPS)–(Birthday to Birthday.32 °C + LPS) | 0.24654 | 0.118 | Inf | 2.084 | 0.0371 |
| Birthday to Birthday.28 °C–Birthday to Birthday.32 °C | −0.32741 | 0.12 | Inf | −2.723 | 0.0065 |
| Birthday to Birthday.28 °C–(Birthday to Birthday.32 °C + LPS) | −0.20105 | 0.115 | Inf | −1.742 | 0.0815 |
| Birthday to Birthday.32 °C–(Birthday to Birthday.32 °C + LPS) | 0.12636 | 0.124 | Inf | 1.022 | 0.3069 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harman, T.E.; Hauff-Salas, B.; Haslun, J.A.; Cervino, J.M.; Strychar, K.B. Decreased Photosynthetic Efficiency in Response to Site Translocation and Elevated Temperature Is Mitigated with LPS Exposure in Porites astreoides Symbionts. Water 2022, 14, 366. https://0-doi-org.brum.beds.ac.uk/10.3390/w14030366
Harman TE, Hauff-Salas B, Haslun JA, Cervino JM, Strychar KB. Decreased Photosynthetic Efficiency in Response to Site Translocation and Elevated Temperature Is Mitigated with LPS Exposure in Porites astreoides Symbionts. Water. 2022; 14(3):366. https://0-doi-org.brum.beds.ac.uk/10.3390/w14030366
Chicago/Turabian StyleHarman, Tyler E., Briana Hauff-Salas, Joshua A. Haslun, James M. Cervino, and Kevin B. Strychar. 2022. "Decreased Photosynthetic Efficiency in Response to Site Translocation and Elevated Temperature Is Mitigated with LPS Exposure in Porites astreoides Symbionts" Water 14, no. 3: 366. https://0-doi-org.brum.beds.ac.uk/10.3390/w14030366