
Enterococcus Species and Their Antimicrobial Resistance in an Urban Watershed Affected by Different Anthropogenic Sources


Abstract
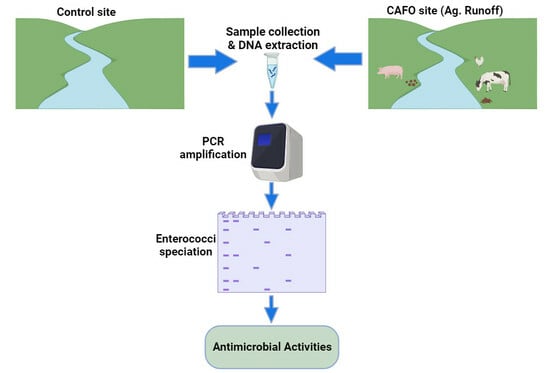
:
1. Introduction
2. Materials and Methods
2.1. Study Area, Sampling Sites, and Sample Collection
2.2. Speciation of Enterococcus Species
2.3. Antimicrobial Resistance Analysis
2.4. PCR Screening of Virulence and Antibiotic Resistance Genes
2.5. Statistical Analysis
3. Results
3.1. Identification of Covariance Structures for Major Physical and Chemical Parameters
3.2. Distribution of Environmental Enterococcus spp. in the Watershed
3.3. Antimicrobial Resistance in Surface Water and Sediment
3.4. Antibiotics Resistance and Virulence Factor Genes in Enterococci Isolated from Sediment and Surface Water Samples
4. Discussion
4.1. Enterococcus Species in the Watershed
4.2. Antimicrobial Resistance in Surface Water and Sediment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zaheer, R.; Cook, S.R.; Barbieri, R.; Goji, N.; Cameron, A.; Petkau, A.; Polo, R.O.; Tymensen, L.; Stamm, C.; Song, J.; et al. Surveillance of Enterococcus spp. reveals distinct species and antimicrobial resistance diversity across a One-Health continuum. Sci. Rep. 2020, 10, 3937. [Google Scholar] [CrossRef] [PubMed]
- Makowska, N.; Bresa, K.; Koczura, R.; Philips, A.; Nowis, K.; Mokracka, J. Urban wastewater as a conduit for pathogenic Gram-positive bacteria and genes encoding resistance to β-lactams and glycopeptides. Sci. Total Environ. 2021, 765, 144176. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, R.E.R.; Micallef, S.A.; Gibbs, S.G.; George, A.; Claye, E.; Sapkota, A.; Joseph, S.W.; Sapkota, A.R. Detection of vancomycin-resistant enterococci (VRE) at four U.S. wastewater treatment plants that provide effluent for reuse. Sci. Total Environ. 2014, 466–467, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, P.; Ziegler, E.; Palmer, K.L. Chlorhexidine induces VanA-type vancomycin resistance genes in enterococci. Antimicrob. Agents Chemother. 2016, 60, 2209–2221. [Google Scholar] [CrossRef] [PubMed]
- Karimi, F.; Samarghandi, M.R.; Shokoohi, R.; Godini, K. Prevalence and removal efficiency of enterococcal species and vancomycin-resistant enterococci of a hospital wastewater treatment plant. Avicenna J. Environ. Health Eng. 2016, 3, e8623. [Google Scholar] [CrossRef]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the Environment. Microbiol. Mol. Biol. Rev. 2012, 76, 685–706. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Hur, H.-G.; Sadowsky, M.; Byappanahalli, M.; Yan, T.; Ishii, S. Environmental Escherichia coli: Ecology and public health implications—A review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Bager, F. DANMAP: Monitoring antimicrobial resistance in Denmark. Int. J. Antimicrob. Agents 2000, 14, 271–274. [Google Scholar] [CrossRef]
- Zainab, S.M.; Junaid, M.; Xu, N.; Malik, R.N. Antibiotics and antibiotic resistant genes (ARGs) in groundwater: A global review on dissemination, sources, interactions, environmental and human health risks. Water Res. 2020, 187, 116455. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Zarzecka, U.; Zakrzewski, A.; Gajewska, J. Enterococci from ready-to-eat food—Horizontal gene transfer of antibiotic resistance genes and genotypic characterization by PCR melting profile. J. Sci. Food Agric. 2019, 99, 1172–1179. [Google Scholar] [CrossRef]
- Choi, J.-M.; Woo, G.-J. Transfer of Tetracycline Resistance Genes with Aggregation Substance in Food-Borne Enterococcus faecalis. Curr. Microbiol. 2015, 70, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Haug, M.C.; Tanner, S.A.; Lacroix, C.; Stevens, M.J.; Meile, L. Monitoring horizontal antibiotic resistance gene transfer in a colonic fermentation model. FEMS Microbiol. Ecol. 2011, 78, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Murinda, S.E.; Graves, A.K. Genetic diversity and antimicrobial resistance of Escherichia coli from human and animal sources uncovers multiple resistances from human sources. PLoS ONE 2011, 6, e20819. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Ma, J.; Murinda, S.E. Bacterial community composition and structure in an Urban River impacted by different pollutant sources. Sci. Total Environ. 2016, 566–567, 1176–1185. [Google Scholar] [CrossRef] [PubMed]
- SAWPA: Santa Ana Watershed Project Authority. Middle Santa Ana River bacterial indicator TMDL 2013 Dry Season Report. Prepared by CDM Smith on behalf of SAWPA and the Middle Santa Ana River Watershed TMDL Task Force. July 2013. Available online: https://sawpa.gov/wp-content/uploads/2018/04/2013-Triennial-Report_Tier-1-Source-Evaluation-Final.pdf (accessed on 1 July 2023).
- RWQCB, Regional Water Quality Control Board. Resolution amending the water quality control plans for the Santa Ana River Basin to incorporate bacterial indicator total maximum daily loads (TMDLs) for middle Santa Ana River Watershed Water bodies. Regional Board Resolution R8-2005-0001. 2005. Available online: https://www.waterboards.ca.gov/board_decisions/adopted_orders/resolutions/2006/rs2006_0030attach.pdf (accessed on 1 July 2023).
- Ibekwe, A.M.; Leddy, M.; Murinda, S.E. Potential human pathogenic bacteria in a mixed urban watershed as revealed by pyrosequencing. PLoS ONE 2013, 8, e79490. [Google Scholar] [CrossRef] [PubMed]
- Ibekwe, A.M.; Leddy, M.B.; Bold, R.M.; Graves, A.K. Bacterial community composition in low-flowing river water with different sources of pollutants. FEMS Microbiol. Ecol. 2012, 79, 155–166. [Google Scholar] [CrossRef]
- American Public Health Association. Standard Methods for the Examination of the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1995. [Google Scholar]
- USEPA. Review of Zoonotic Pathogens in Ambient Waters, 822-R-09-002; USEPA Office of Water, Ed.; Office of Water, U.S. Environmental Protection Agency: Washington, DC, USA, 2009.
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B.; Ladely, S.R. Effects of tylosin use on erythromycin resistance in enterococci isolated from swine. Appl. Environ. Microbiol. 2004, 70, 4205–4210. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Ibekwe, A.M.; Lesch, S.M.; Bold, R.M.; Leddy, M.B.; Graves, A.K. Variations of indicator bacteria in a large urban watershed. Trans. ASABE 2011, 54, 2227–2236. [Google Scholar] [CrossRef]
- Kenward, M.G.; Roger, J.H. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef]
- Cho, S.; Hiott, L.M.; McDonald, J.M.; Barrett, J.B.; McMillan, E.A.; House, S.L.; Adams, E.S.; Frye, J.G.; Jackson, C.R. Diversity and antimicrobial resistance of Enterococcus from the Upper Oconee Watershed, Georgia. J. Appl. Microbiol. 2019, 128, 1221–1233. [Google Scholar] [CrossRef]
- Cho, S.; Jackson, C.; Frye, J. The prevalence and antimicrobial resistance phenotypes of Salmonella, Escherichia coli and Enterococcus sp. in surface water. Lett. Appl. Microbiol. 2020, 71, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Diarra, M.S.; Rempel, H.; Champagne, J.; Masson, L.; Pritchard, J.; Topp, E. Distribution of antimicrobial resistance and virulence genes in Enterococcus spp. and characterization of isolates from broiler chickens. Appl. Environ. Microbiol. 2010, 76, 8033–8043. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.; Barrett, J.B.; Frye, J.G.; Jackson, C.R. Antimicrobial resistance gene detection and plasmid typing among multidrug resistant enterococci isolated from freshwater environment. Microorganisms 2020, 8, 1338. [Google Scholar] [CrossRef]
- Thumu, S.C.R.; Halami, P.M. Acquired Resistance to Macrolide-Lincosamide-Streptogramin Antibiotics in Lactic Acid Bacteria of Food Origin. Indian J. Microbiol. 2012, 52, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.; Zhanel, G.G.; Sparling, R.; Holley, R.A. Horizontal transfer of antibiotic resistance from Enterococcus faecium of fermented meat origin to clinical isolates of E. faecium and Enterococcus faecalis. Int. J. Food Microbiol. 2015, 199, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.; Mac, K.; Wichmann-Schauer, H.; Klein, G.; Ellerbroek, L. Species distribution and antibiotic resistance patterns of enterococci isolated from food of animal origin in Germany. Int. J. Food Microbiol. 2003, 88, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Del Papa, M.F.; Hancock, L.E.; Thomas, V.C.; Perego, M. Full activation of Enterococcus faecalis gelatinase by a C-terminal proteolytic cleavage. J. Bacteriol. 2007, 189, 8835–8843. [Google Scholar] [CrossRef] [PubMed]
- Thurlow, L.R.; Thomas, V.C.; Narayanan, S.; Olson, S.; Fleming, S.D.; Hancock, L.E. Gelatinase contributes to the pathogenesis of endocarditis caused by Enterococcus faecalis. Infect. Immun. 2010, 78, 4936–4943. [Google Scholar] [CrossRef]
- Ali, L.; Goraya, M.U.; Arafat, Y.; Ajmal, M.; Chen, J.L.; Yu, D. Molecular mechanism of quorum-sensing in Enterococcus faecalis: Its role in virulence and therapeutic approaches. Int. J. Mol. Sci. 2017, 18, 960. [Google Scholar] [CrossRef]
- Zeng, J.; Teng, F.; Murray, B.E. Gelatinase is important for translocation of Enterococcus faecalis across polarized human enterocyte-like T84 cells. Infect. Immun. 2005, 73, 1606–1612. [Google Scholar] [CrossRef]
- Dapkevicius, M.d.L.E.; Sgardioli, B.; Câmara, S.P.A.; Poeta, P.; Malcata, F.X. Current trends of enterococci in dairy products: A comprehensive review of their multiple roles. Foods 2021, 10, 821. [Google Scholar] [CrossRef] [PubMed]
- Spirli, H.; Demirbaş, F.; Dertli, E. Characterization of functional properties of Enterococcus spp. isolated from Turkish white cheese. LWT Food Sci. Technol. 2017, 75, 358–365. [Google Scholar] [CrossRef]
- Zkan, E.R.; Demirci, T.; Akın, N. In vitro assessment of probiotic and virulence potential of Enterococcus faecium strains derived from artisanal goatskin casing Tulum cheeses produced in central Taurus Mountains of Turkey. LWT 2021, 141, 110908. [Google Scholar]
- Iweriebor, B.C.; Obi, L.C.; Okoh, A.I. Virulence and antimicrobial resistance factors of Enterococcus spp. isolated from fecal samples from piggery farms in Eastern Cape, South Africa. BMC Microbiol. 2015, 15, 136. [Google Scholar] [CrossRef] [PubMed]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Łaniewska-Trokenheim, Ł. Virulence factors, antimicrobial resistance and biofilm formation in Enterococcus spp. isolated from retail shrimps. LWT Food Sci. Technol. 2016, 69, 117–122. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, S.; Shang, X.; Wang, X.; Yan, Z.; Li, H.; Li, J. Short communication: Antimicrobial resistance and virulence genes of Enterococcus faecalis isolated from subclinical bovine mastitis cases in China. J. Dairy Sci. 2019, 102, 140–144. [Google Scholar] [CrossRef]
- Cho, S.; Hiott, L.M.; Read, Q.D.; Damashek, J.; Westrich, J.; Edwards, M.; Seim, R.F.; Glinski, D.A.; McDonald, J.M.B.; Ottesen, E.A.; et al. Distribution of antibiotic resistance in a mixed-use watershed and the impact of wastewater treatment plants on antibiotic resistance in surface water. Antibiotics 2023, 12, 1586. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Xu, J.; Xu, Y.; Wang, H.; Guo, C.; Qiu, H.; He, Y.; Zhang, Y.; Li, X.; Meng, W. Occurrence of antibiotics and antibiotic resistance genes in a sewage treatment plant and its effluent-receiving river. Chemosphere 2015, 119, 1379–1385. [Google Scholar] [CrossRef]
- Rodriguez-Mozaz, S.; Chamorro, S.; Marti, E.; Huerta, B.; Gros, M.; Sànchez-Melsió, A.; Borrego, C.M.; Barceló, D.; Balcázar, J.L. Occurrence of antibiotics and antibiotic resistance genes in hospital and urban wastewaters and their impact on the receiving river. Water Res. 2015, 69, 234–242. [Google Scholar] [CrossRef] [PubMed]
- An, X.-L.; Su, J.-Q.; Li, B.; Ouyang, W.-Y.; Zhao, Y.; Chen, Q.-L.; Cui, L.; Chen, H.; Gillings, M.R.; Zhang, T.; et al. Tracking antibiotic resistome during wastewater treatment using high throughput quantitative PCR. Environ. Int. 2018, 117, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Hembach, N.; Schwartz, T. Evaluation of antibiotic resistance dissemination by wastewater treatment plant effluents with different catchment areas in Germany. Sci. Rep. 2020, 10, 8952. [Google Scholar] [CrossRef] [PubMed]
- Ashworth, D.J.; Ibekwe, A.M. System of multi-layered environmental media for the removal of antibiotics from wastewater. J. Environ. Chem. Eng. 2020, 8, 104206. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Species | Amp * | Chl | Gen | Pen | Cip | Ery | Tet | Van |
|---|---|---|---|---|---|---|---|---|---|
| WWTP | E. faecalis (28) | 0 | 0 | 0 | 0 | 0 | 0 | 1 (3) | 0 |
| E. faecium (24) | 0 | 0 | 0 | 0 | 2 (8) | 21 (87) | 3 (12) | 0 | |
| E. mundtii (58) | 0 | 0 | 0 | 0 | 0 | 48 (83) | 0 | 0 | |
| Urban runoff | E. faecalis (116) | 0 | 1 (1) | 0 | 0 | 10 (8) | 4 (3) | 44 (38) | 0 |
| E. faecium (181) | 0 | 0 | 0 | 3 (2) | 78 (43) | 45 (25) | 109 (60) | 0 | |
| E. mundtii (116) | 0 | 0 | 0 | 0 | 0 | 11 (9) | 3 (2) | 0 | |
| Recreational runoff | E. faecalis (29) | 0 | 1(3) | 0 | 0 | 10 (34) | 0 | 4 (14) | 0 |
| E. faecium (89) | 0 | 0 | 0 | 13 (15) | 42 (47) | 22 (25) | 0 | 0 | |
| E. mundtii (39) | 0 | 0 | 0 | 0 | 0 | 0 | 1 (2) | 0 | |
| Ag. runoff | E. faecalis (48) | 0 | 0 | 0 | 0 | 3 (6) | 3 (6) | 10 (21) | 0 |
| E. faecium (61) | 0 | 0 | 0 | 0 | 36 (59) | 16 (26) | 30 (49) | 0 | |
| E. mundtii (65) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Control | E. faecalis (60) | 0 | 0 | 0 | 8 (13) | 2 (3) | 0 | 0 | 0 |
| E. faecium (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| E. mundtii (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sites | Species | Amp * | Chl | Gen | Pen | Cip | Ery | Tet | Van |
|---|---|---|---|---|---|---|---|---|---|
| WWTP | E. faecalis (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| E. faecium (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| E. mundtii (19) | 0 | 0 | 0 | 0 | 0 | 19 (100) | 0 | 0 | |
| Urban runoff | E. faecalis (113) | 0 | 0 | 0 | 0 | 3 (2) | 8 (7) | 24 (21) | 0 |
| E. faecium (43) | 0 | 0 | 0 | 1 (2) | 33 (77) | 9 (21) | 30 (70) | 0 | |
| E. mundtii (126) | 0 | 0 | 0 | 0 | 0 | 38 (30) | 2 (1) | 0 | |
| Recreational runoff | E. faecalis (0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| E. faecium (61) | 0 | 0 | 0 | 1 (2) | 50 (82) | 33 (54) | 31 (51) | 0 | |
| E. mundtii (80) | 0 | 0 | 0 | 0 | 0 | 36 (45) | 0 | 0 | |
| Ag. runoff | E. faecalis (86) | 0 | 9 (10) | 1 (1) | 0 | 4 (5) | 1 (1) | 18 (21) | 0 |
| E. faecium (11) | 0 | 0 | 0 | 0 | 0 | 0 | 11(100) | 0 | |
| E. mundtii (102) | 0 | 2 (2) | 0 | 0 | 0 | 0 | 5 (5) | 0 | |
| Control | E. faecalis (35) | 0 | 2 (6) | 0 | 0 | 2 (6) | 1 (3) | 19 (54) | 0 |
| E. faecium (22) | 0 | 0 | 0 | 12 (55) | 22 (100) | 0 | 0 | 0 | |
| E. mundtii (20) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sites | Species | tet (A) | tet (C) | tet (Q) | tet (K) | tet (O) | tet (S) | van (A) | van (B) | van (C1) | van (C2) | erm (B) | asa1 | cylA | esp | gelE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WWTP | E. faecalis | 0 | 56 (10) | 0 | 28 (5) | 56 (10) | 83 (15) | 0 | 0 | 0 | 0 | 0 | 78 (14) | 0 | 0 | 78 (14) |
| E. faecium | 0 | 86 (6) | 0 | 14 (1) | 14 (1) | 14 (1) | 0 | 0 | 0 | 0 | 0 | 29 (20) | 0 | 0 | 29 (2) | |
| Urban runoff | E. faecalis | 0 | 79 (48) | 0 | 16 (10) | 57 (35) | 80 (49) | 0 | 0 | 2 (1) | 0 | 8 (5) | 46 (28) | 0 | 0 | 97 (59) |
| E. faecium | 0 | 0 | 0 | 0 | 9 (9) | 3 (3) | 0 | 0 | 5 (5) | |||||||
| Recreational runoff | E. faecalis | 0 | 22 (5) | 0 | 4 (1) | 9 (2) | 87 (20)0 | 0 | 0 | 0 | 13 (3) | 0 | 22 (5) | 4 (1) | 4 (1) | 87 (20) |
| E. faecium | 0 | 98 (55) | 0 | 2 (1) | 20 (11) | 45 (25) | 0 | 0 | 0 | 0 | 5 (3) | 0 | 0 | 0 | 0 | |
| Ag. runoff | E. faecalis | 0 | 78 (28) | 0 | 0 | 17 (6) | 83 (30) | 0 | 0 | 0 | 3 (1) | 3 (1) | 17 (6) | 0 | 19 (7) | 94 (34) |
| E. faecium | 0 | 95 (40) | 0 | 5 (2) | 19 (8) | 45 (19) | 0 | 0 | 0 | 0 | 5 (2) | 2 (1) | 0 | 0 | 2 (1) | |
| Control | E. faecalis | 0 | 55 (11) | 0 | 0 | 0 | 95 (19) | 0 | 0 | 0 | 0 | 5 (1) | 70 (14) | 0 | 0 | 100 (20) |
| E. faecium |
| Sites | Species | tet (A) | tet (C) | tet (Q) | tet (K) | tet (O) | tet (S) | van (A) | van (B) | van (C1) | van (C2) | erm (B) | asa1 | cylA | esp | gelE |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WWTP | E. faecalis | |||||||||||||||
| E. faecium | ||||||||||||||||
| Urban runoff | E. faecalis | 0 | 90 (45) | 0 | 6 (3) | 40 (20) | 88 (44) | 0 | 0 | 0 | 0 | 4 (2) | 38 (19) | 0 | 10 (5) | 100 (50) |
| E. faecium | 0 | 75 (15) | 0 | 25 (5) | 0 | 50 (10) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Recreational runoff | E. faecalis | |||||||||||||||
| E. faecium | 0 | 97 (28) | 0 | 0 | 3 (1) | 62 (18) | 0 | 0 | 0 | 0 | 0 | 7 (2) | 0 | 0 | 3 (1) | |
| Ag. runoff | E. faecalis | 0 | 83 (33) | 0 | 25 (10) | 50 (20) | 65 (26) | 0 | 0 | 0 | 0 | 0 | 65 (26) | 33 (13) | 48 (19) | 78 (31) |
| E. faecium | 0 | 100 (8) | 0 | 63 (50) | 25 (2) | 38 (3) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| E. mundtii | ||||||||||||||||
| Control | E. faecalis | 0 | 57 (8) | 0 | 21 (3) | 7 (1) | 50 (7) | 0 | 0 | 0 | 0 | 0 | 86 (12) | 0 | 43 (6) | 100 (14) |
| E. faecium | 0 | 100 (6) | 0 | 0 | 0 | 100 (6) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Sample Name | Species | No. Isolates | Amp | Gen | Cip | Ery | Chl | Pen | Tet | Van |
|---|---|---|---|---|---|---|---|---|---|---|
| Recreation sediment | E. faecium | 26 | S | S | R | R | S | S | R | S |
| Urban runoff sediment | E. faecium | 9 | S | S | R | R | S | S | R | S |
| Agricultural water | E. faecium | 2 | S | S | R | R | S | S | R | S |
| Recreation water | E. faecium | 1 | S | S | R | S | S | R | R | S |
| Recreation water | E. faecium | 8 | S | S | R | R | S | S | R | S |
| WWTP | E. faecium | 1 | S | S | R | R | S | S | R | S |
| Urban Runoff water | E. faecium | 1 | S | S | R | I | S | R | R | S |
| Urban Runoff water | E. faecium | 15 | S | S | R | R | S | S | R | S |
| E. faecalis | ||||||||||
| Agricultural sediment | E. faecalis | 2 | S | S | R | I | R | S | R | I |
| Control | E. faecalis | 1 | S | S | R | I | R | S | R | I |
| Urban Runoff sediment | E. faecalis | 15 | S | S | R | R | S | S | R | S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibekwe, A.M.; Obayiuwana, A.C.; Murinda, S.E. Enterococcus Species and Their Antimicrobial Resistance in an Urban Watershed Affected by Different Anthropogenic Sources. Water 2024, 16, 116. https://0-doi-org.brum.beds.ac.uk/10.3390/w16010116
Ibekwe AM, Obayiuwana AC, Murinda SE. Enterococcus Species and Their Antimicrobial Resistance in an Urban Watershed Affected by Different Anthropogenic Sources. Water. 2024; 16(1):116. https://0-doi-org.brum.beds.ac.uk/10.3390/w16010116
Chicago/Turabian StyleIbekwe, Abasiofiok M., Amarachukwu C. Obayiuwana, and Shelton E. Murinda. 2024. "Enterococcus Species and Their Antimicrobial Resistance in an Urban Watershed Affected by Different Anthropogenic Sources" Water 16, no. 1: 116. https://0-doi-org.brum.beds.ac.uk/10.3390/w16010116