Ion Binding Properties of a Naturally Occurring Metalloantibody


Abstract
:1. Introduction
2. Materials and Methods
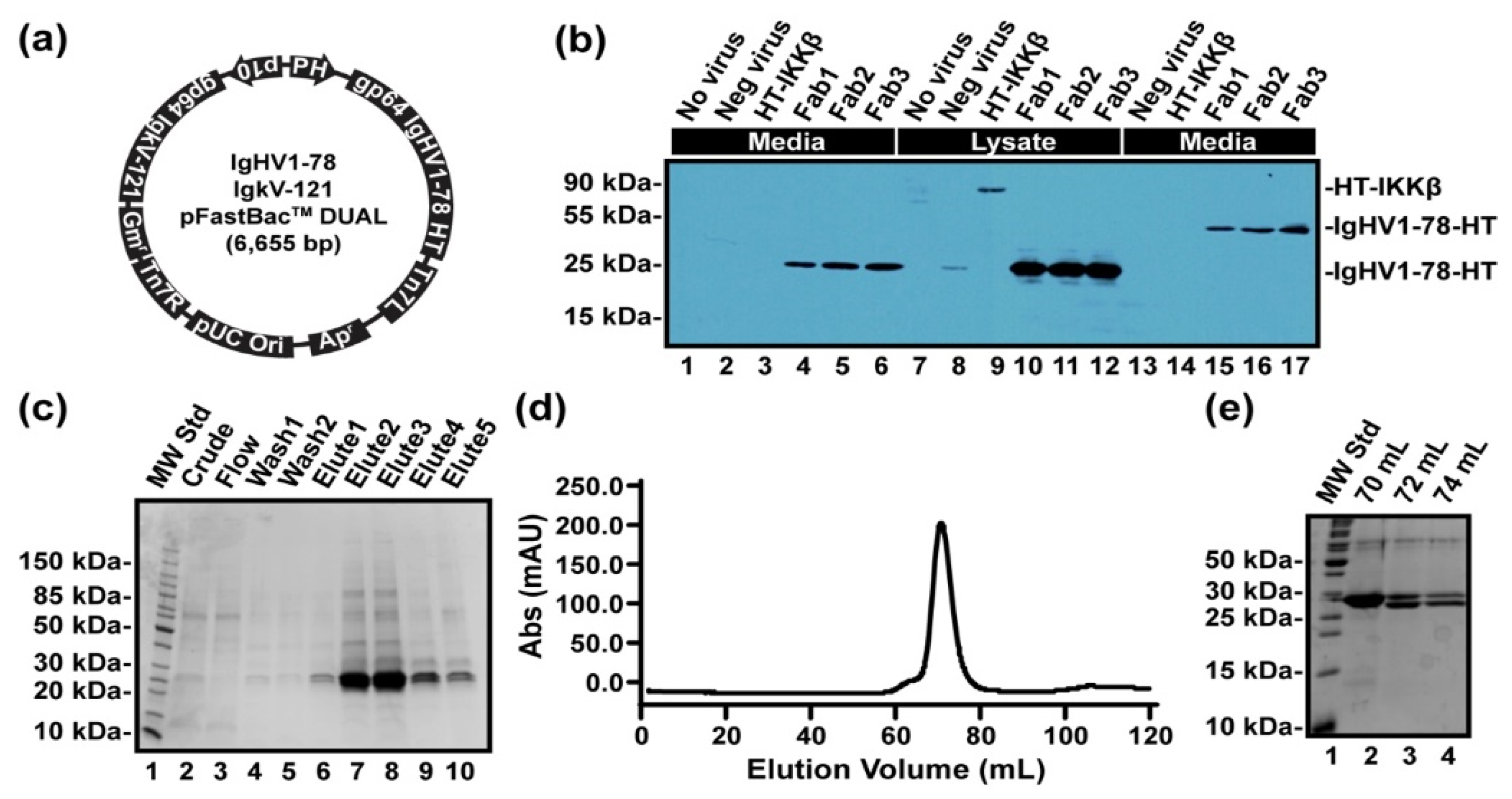
2.1. DNA, Oligonucleotides, and Plasmids
2.2. Antibodies and Fab Fragment Generation
2.3. LT1009 Fab:Ca2+ Complex Formation and Co-Crystallization
2.4. X-ray Crystallography
2.5. Equilibrium Dialysis
2.6. Flame Atomic Absorption Spectroscopy (FAAS)
2.7. Inductively-Coupled Plasma-Mass Spectrometry (ICP-MS)
2.8. Isothermal Titration Calorimetry (ITC)
2.9. Antibody Sequence Analysis
3. Results
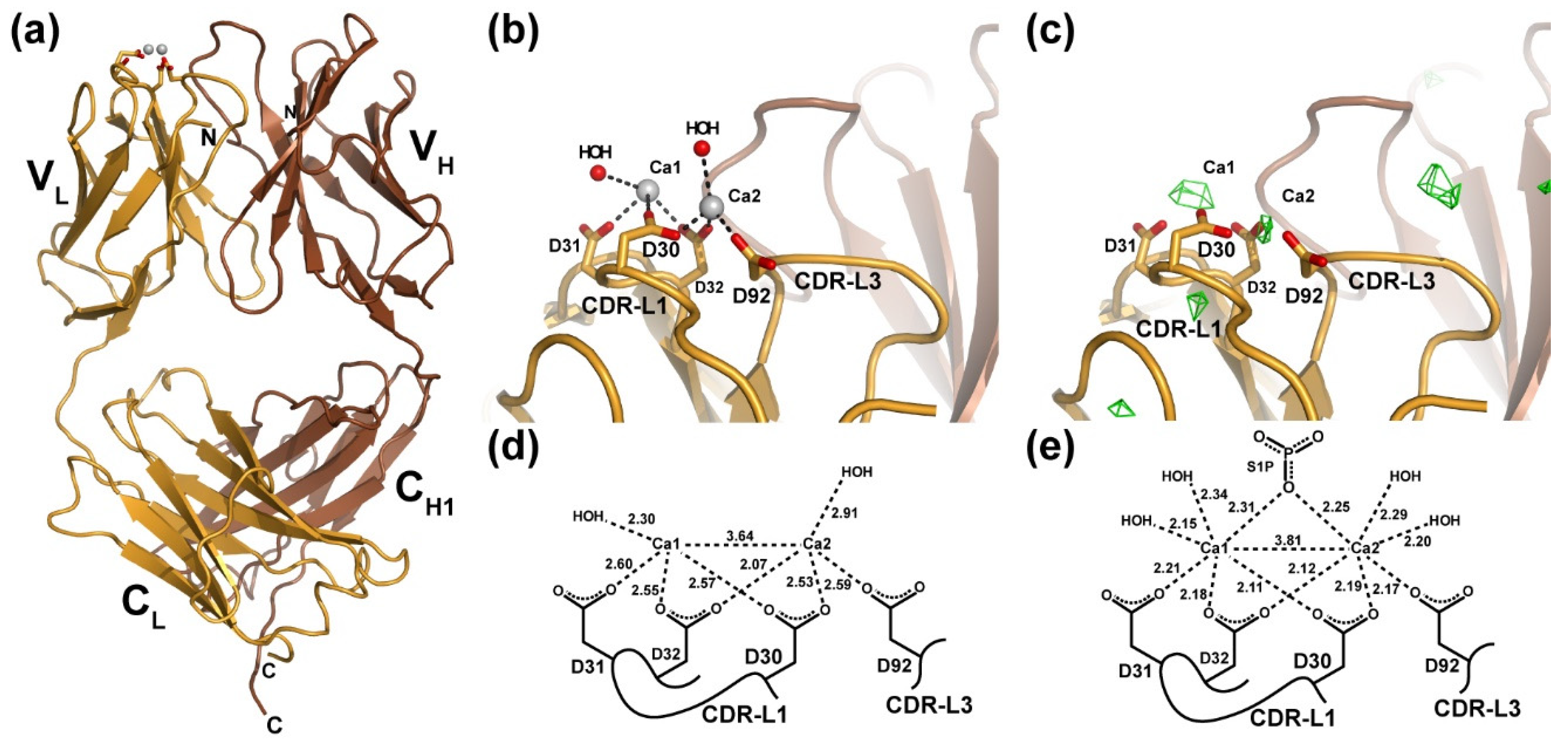
3.1. X-ray Crystal Structure of a Ca2+-Bound Anti-S1P Antibody Fab Fragment in the Absence of Antigen
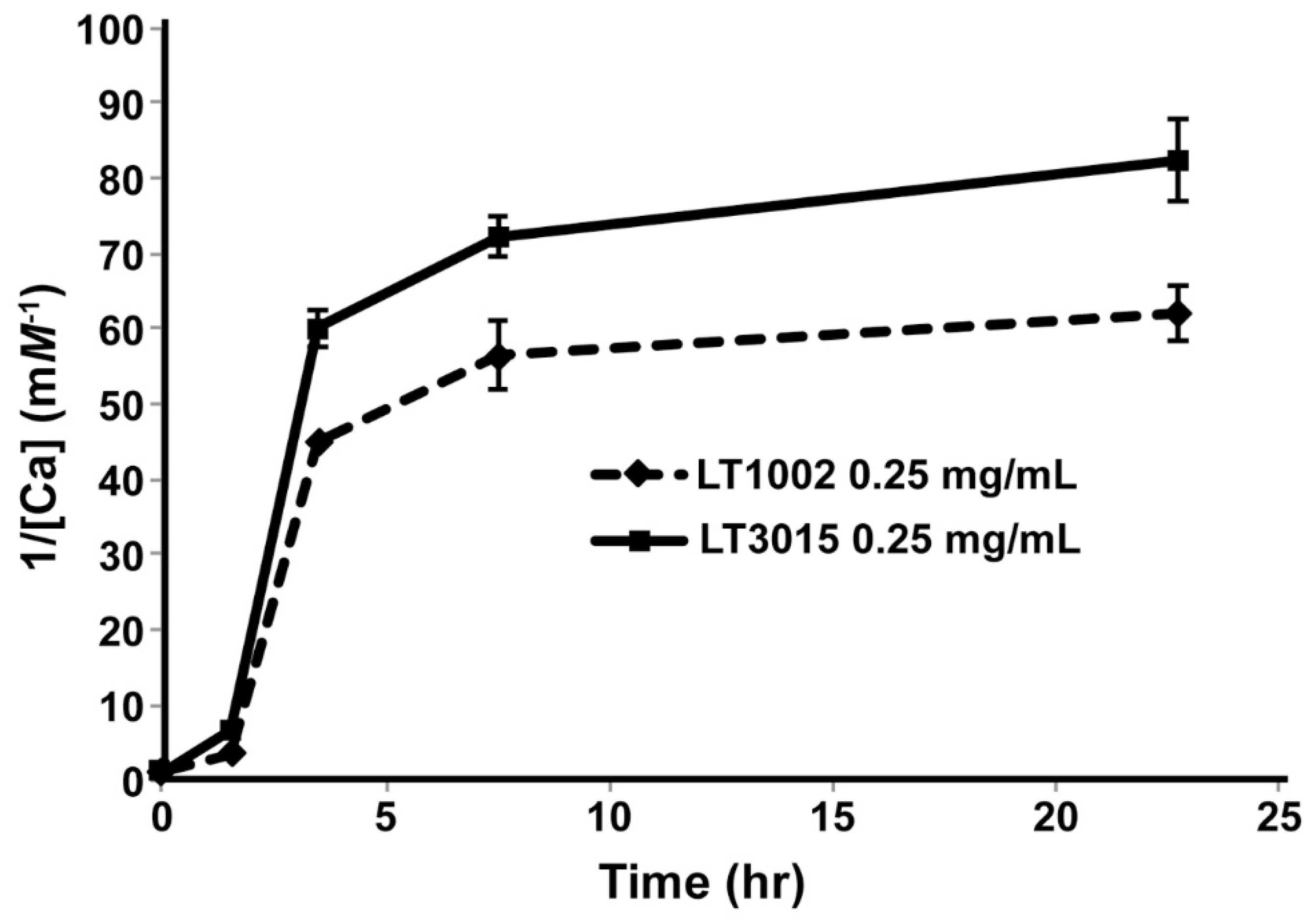
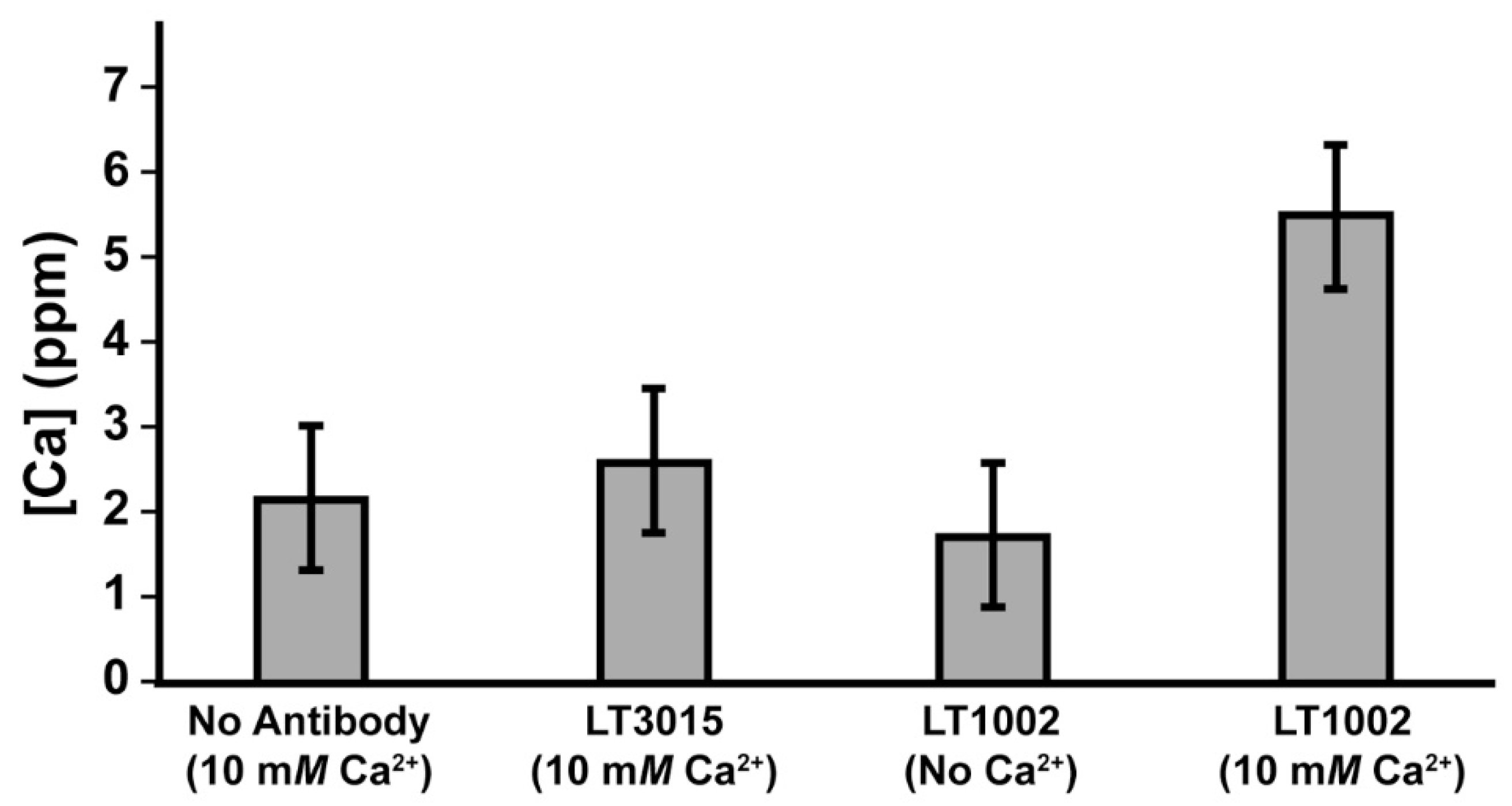
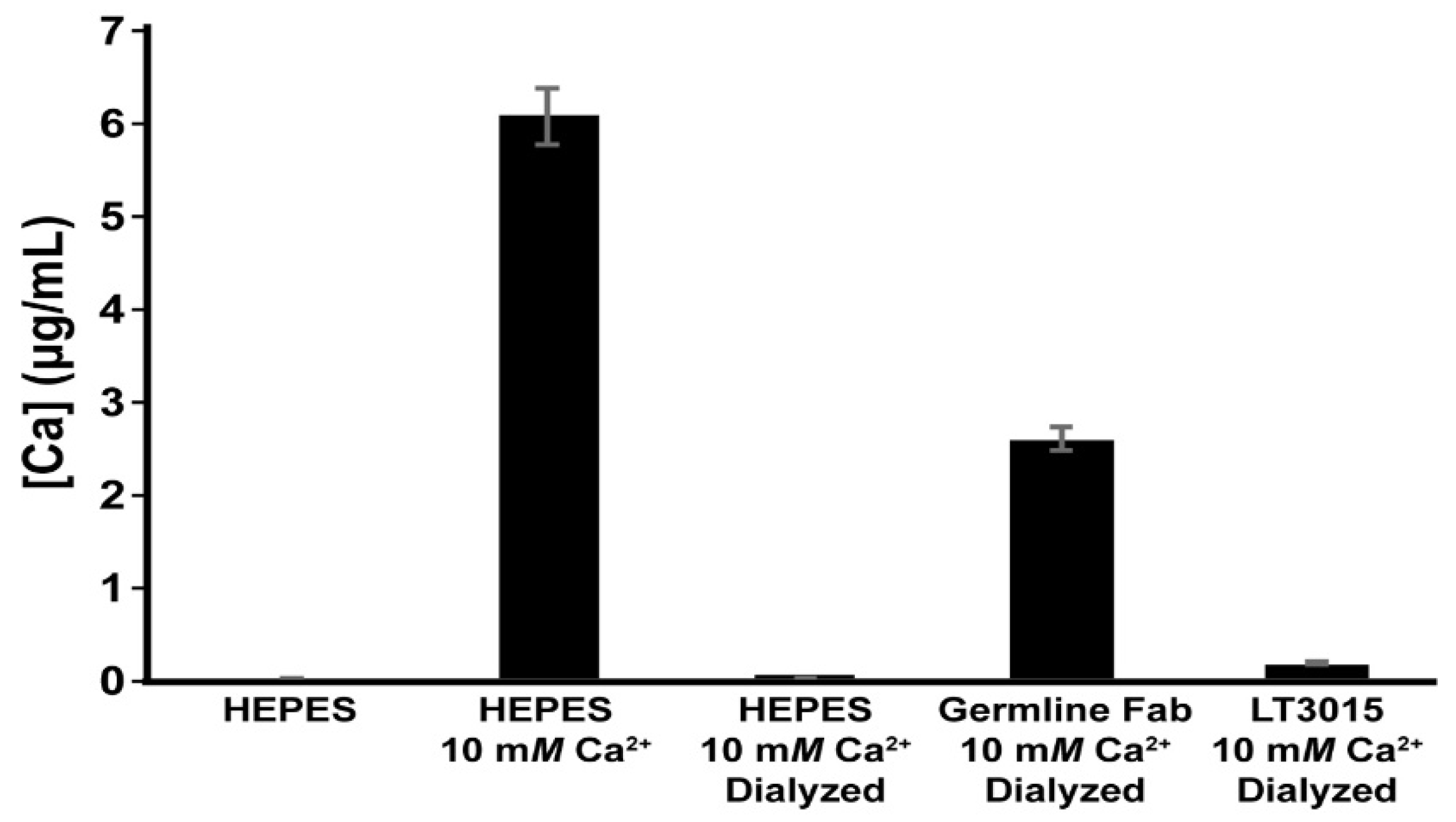
3.2. Detection of Ca2+ Binding by LT1002 in Solution
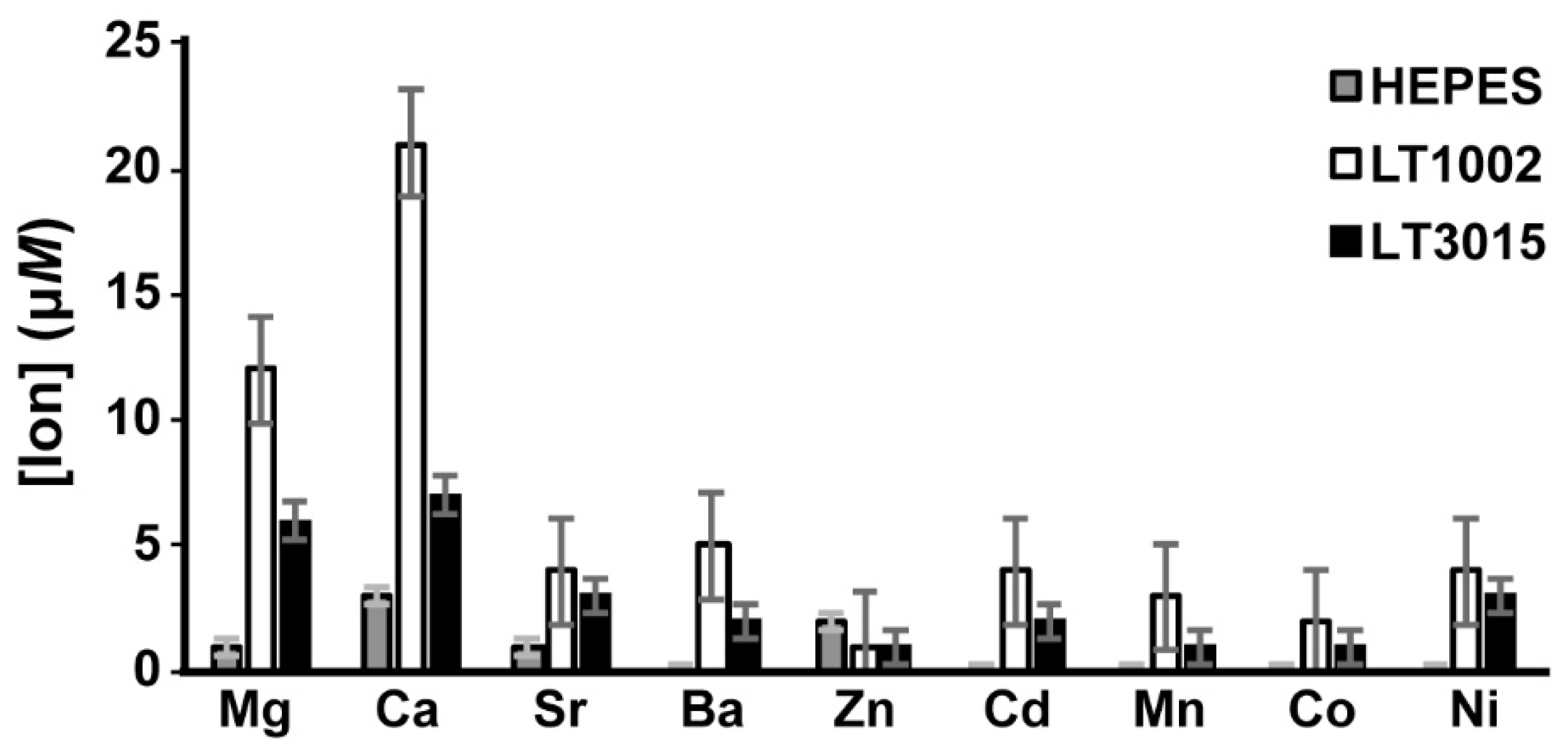
3.3. Metal Ion Binding Specificity of LT1002
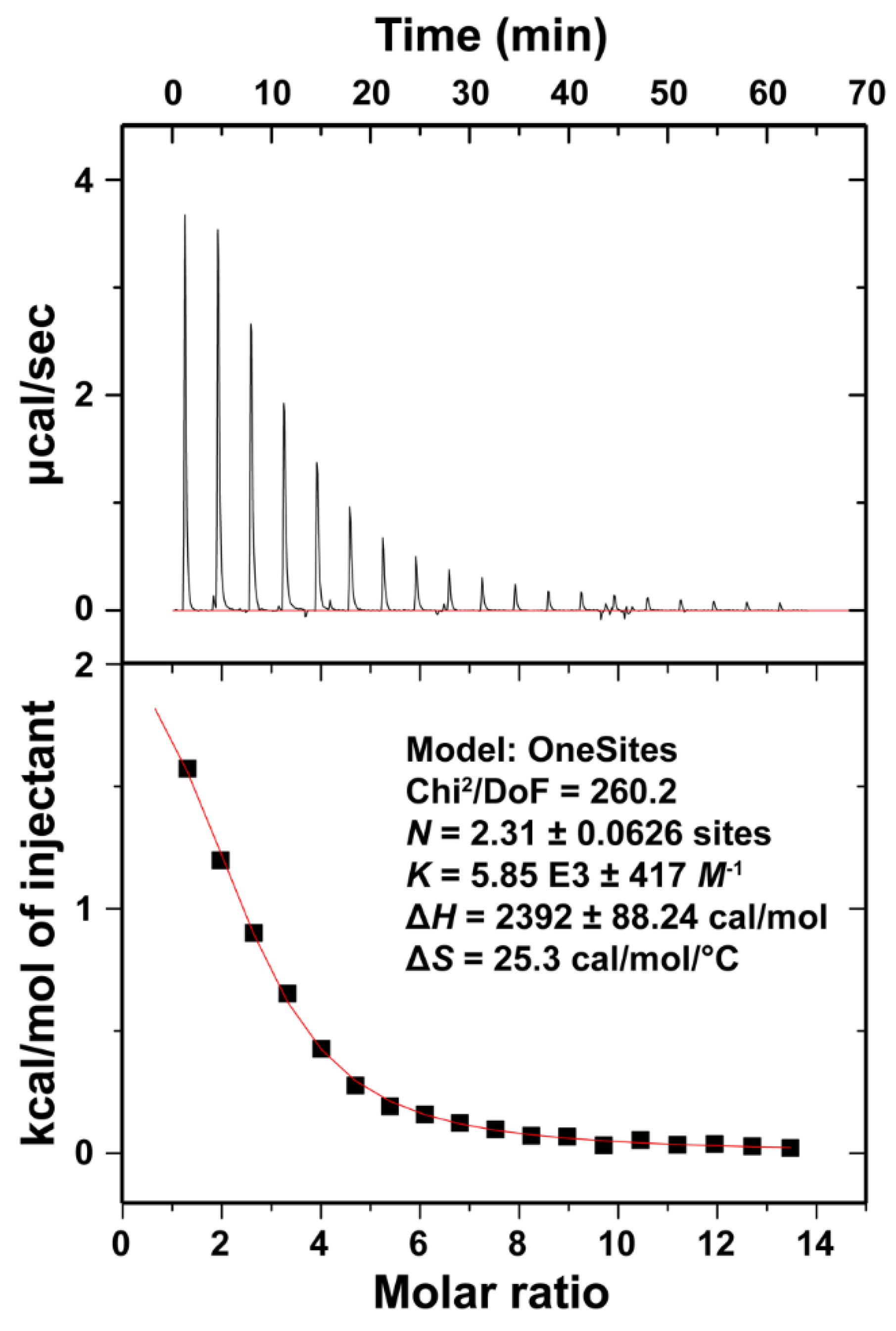
3.4. Isothermal Titration Calorimetry of Ca2+ Binding to LT1002
3.5. Sequence Conservation of LT1002 Ca2+-Coordinating Residues in Other Antibodies
3.6. LT1002 Ca2+-Coordinating Residues Are Encoded in Diverse Light Chain Germline Sequences
3.7. The Germline-Encoded “Precursor” to LT1002 Binds Ca2+
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thomson, A.J.; Gray, H.B. Bio-inorganic chemistry. Curr. Opin. Chem. Biol. 1998, 2, 155–158. [Google Scholar] [CrossRef]
- Waldron, K.J.; Robinson, N.J. How do bacterial cells ensure that metalloproteins get the correct metal? Nat. Rev. Microbiol. 2009, 7, 25–35. [Google Scholar] [CrossRef]
- Hu, C.; Chan, S.I.; Sawyer, E.B.; Yu, Y.; Wang, J. Metalloprotein design using genetic code expansion. Chem. Soc. Rev. 2014, 43, 6498–6510. [Google Scholar] [CrossRef]
- Kretsinger, R.H.; Uversky, V.N.; Permyakov, E.A. Encyclopedia of Metalloproteins; Springer: New York, NY, USA, 2013; p. 2574. [Google Scholar]
- O’Brien, N.; Jones, S.T.; Williams, D.G.; Cunningham, H.B.; Moreno, K.; Visentin, B.; Gentile, A.; Vekich, J.; Shestowsky, W.; Hiraiwa, M.; et al. Production and characterization of monoclonal anti-sphingosine-1-phosphate antibodies. J. Lipid Res. 2009, 50, 2245–2257. [Google Scholar] [CrossRef] [Green Version]
- Wojciak, J.M.; Zhu, N.; Schuerenberg, K.T.; Moreno, K.; Shestowsky, W.S.; Hiraiwa, M.; Sabbadini, R.; Huxford, T. The crystal structure of sphingosine-1-phosphate in complex with a Fab fragment reveals metal bridging of an antibody and its antigen. Proc. Natl. Acad. Sci. USA 2009, 106, 17717–17722. [Google Scholar] [CrossRef] [Green Version]
- Furuta, T.; Ogawa, T.; Katsuda, T.; Fujii, I.; Yamaji, H. Efficient production of an antibody Fab fragment using the baculovirus-insect cell system. J. Biosci. Bioeng. 2010, 110, 577–581. [Google Scholar] [CrossRef]
- Shaul, J.D.; Farina, A.; Huxford, T. The human IKKβ subunit kinase domain displays CK2-like phosphorylation specificity. Biochem. Biophys. Res. Commun. 2008, 374, 592–597. [Google Scholar] [CrossRef]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar]
- King, A.N.; Fleming, J.K.; Knapik, S.S.; Visentin, B.; Wojciak, J.M.; Huxford, T. High-affinity pan-specific monoclonal antibodies that target cysteinyl leukotrienes and show efficacy in an acute model of colitis. J. Lipid Res. 2017, 58, 1386–1398. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Vagin, A.A.; Dodson, E.J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D Biol. Crystallogr. 1997, 53, 240–255. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B., 3rd; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [Green Version]
- DeLano, W.L. The PyMOL Molecular Graphics System; DeLano Scientific: Palo Alto, CA, USA, 2002. [Google Scholar]
- Ye, J.; Ma, N.; Madden, T.L.; Ostell, J.M. IgBLAST: An immunoglobulin variable domain sequence analysis tool. Nucleic Acids Res. 2013, 41, W34–W40. [Google Scholar] [CrossRef] [Green Version]
- Swindells, M.B.; Porter, C.T.; Couch, M.; Hurst, J.; Abhinandan, K.R.; Nielsen, J.H.; Macindoe, G.; Hetherington, J.; Martin, A.C. abYsis: Integrated antibody sequence and structure-management, analysis, and prediction. J. Mol. Biol. 2017, 429, 356–364. [Google Scholar] [CrossRef] [Green Version]
- Madden, T.L.; Tatusov, R.L.; Zhang, J. Applications of network BLAST server. Methods Enzymol. 1996, 266, 131–141. [Google Scholar]
- Healey, D.; Dianda, L.; Moore, J.P.; McDougal, J.S.; Moore, M.J.; Estess, P.; Buck, D.; Kwong, P.D.; Beverley, P.C.; Sattentau, Q.J. Novel anti-CD4 monoclonal antibodies separate human immunodeficiency virus infection and fusion of CD4+ cells from virus binding. J. Exp. Med. 1990, 172, 1233–1242. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Hamer, D.H.; Hendrickson, W.A.; Sattentau, Q.J.; Kwong, P.D. Interfacial metal and antibody recognition. Proc. Natl. Acad. Sci. USA 2005, 102, 14575–14580. [Google Scholar] [CrossRef] [Green Version]
- Stanfield, R.L.; Zemla, A.; Wilson, I.A.; Rupp, B. Antibody elbow angles are influenced by their light chain class. J. Mol. Biol. 2006, 357, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Gagné, O.C.; Hawthorne, F.C. Bond-length distributions for ions bonded to oxygen: Alkali and alkaline-earth metals. Acta Crystallogr. B Struct. Sci. Cryst. Eng. Mater. 2016, 72, 602–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, J.K.; Wojciak, J.M.; Campbell, M.A.; Huxford, T. Biochemical and structural characterization of lysophosphatidic acid binding by a humanized monoclonal antibody. J. Mol. Biol. 2011, 408, 462–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman Katz, A.; Glusker, J.P.; Beebe, S.A.; Bock, C.W. Calcium ion coordination: A comparison with that of beryllium, magnesium, and zinc. J. Am. Chem. Soc. 1996, 118, 5752–5763. [Google Scholar] [CrossRef]
- Grossoehme, N.E.; Spuches, A.M.; Wilcox, D.E. Application of isothermal titration calorimetry in bioinorganic chemistry. J. Biol. Inorg. Chem. 2010, 15, 1183–1191. [Google Scholar] [CrossRef]
- Wilcox, D.E. Isothermal titration calorimetry of metal ions binding to proteins: An overview of recent studies. Inorg. Chim. Acta 2008, 361, 857–867. [Google Scholar] [CrossRef]
- Ràfols, C.; Bosch, E.; Barbas, R.; Prohens, P. The Ca2+-EDTA chelation as standard reaction to validate isothermal titration calorimeter measurements (ITC). Talanta 2016, 154, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Kubota, T.; Akatsuka, T.; Kanai, Y. A monoclonal anti-double stranded DNA antibody from an autoimmune MRL/Mp-lpr/lpr mouse: Specificity and idiotype in serum immunoglobulins. Immunol. Lett. 1986, 14, 53–58. [Google Scholar] [CrossRef]
- Kubota, T.; Watanabe, N.; Kanai, Y.; Stollar, B.D. Enhancement of oxidative cleavage of DNA by the binding sites of two anti-double-stranded DNA antibodies. J. Biol. Chem. 1996, 271, 6555–6561. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Ishizawa, M.; Kubota, T. Monoclonal anti-dsDNA antibody 2C10 escorts DNA to intracellular DNA sensors in normal mononuclear cells and stimulates secretion of multiple cytokines implicated in lupus pathogenesis. Clin. Exp. Immunol. 2020, 199, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Landry, R.C.; Klimowicz, A.C.; Lavictoire, S.J.; Borisova, S.; Kottachchi, D.T.; Lorimer, I.A.; Evans, S.V. Antibody recognition of a conformational epitope in a peptide antigen: Fv-peptide complex of an antibody fragment specific for the mutant EGF receptor, EGFRvIII. J. Mol. Biol. 2001, 308, 883–893. [Google Scholar] [CrossRef]
- Li, Z.; Woo, C.J.; Iglesias-Ussel, M.D.; Ronai, D.; Scharff, M.D. The generation of antibody diversity through somatic hypermutation and class switch recombination. Genes Dev. 2004, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Jean, C.; Folch, G.; Lefranc, M.P. Nomenclature and overview of the mouse (Mus musculus and Mus sp.) immunoglobulin kappa (IGK) genes. Exp. Clin. Immunogenet. 2001, 18, 255–279. [Google Scholar] [CrossRef]
- Brekke, K.M.; Garrard, W.T. Assembly and analysis of the mouse immunoglobulin kappa gene sequence. Immunogenetics 2004, 56, 490–505. [Google Scholar] [CrossRef]
- Vollmer, J.; Fritz, M.; Dormoy, A.; Weltzien, H.U.; Moulon, C. Dominance of the BV17 element in nickel-specific human T cell receptors relates to severity of contact sensitivity. Eur. J. Immunol. 1997, 27, 1865–1874. [Google Scholar] [CrossRef]
- Lu, L.; Vollmer, J.; Moulon, C.; Weltzien, H.U.; Marrack, P.; Kappler, J. Components of the ligand for a Ni++ reactive human T cell clone. J. Exp. Med. 2003, 197, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Crawford, F.; Marrack, P.; Kappler, J.W.; Dai, S. T-cell receptor (TCR) interaction with peptides that mimic nickel offers insight into nickel contact allergy. Proc. Natl. Acad. Sci. USA 2012, 109, 18517–18522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mordasini, T.; Curioni, A.; Andreoni, W. Why do divalent metal ions either promote or inhibit enzymatic reactions? The case of BamHI restriction endonuclease from combined quantum-classical simulations. J. Biol. Chem. 2003, 278, 4381–4384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeraer, Y.; Rabijns, A.; Collet, J.-F.; van Schaftingen, E.; de Ranter, C. How calcium inhibits the magnesium-dependent enzyme human phosphoserine phosphatase. Eur. J. Biochem. 2004, 271, 3421–3427. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LT1009 Fab:Ca2+ Complex | |
|---|---|
| Data collection | |
| X-ray source | ALS 4.2.2 |
| Wavelength (Å) | 1.0000 |
| Space Group | I222 |
| Unit cell (Å) | |
| a | 87.80 |
| b | 114.41 |
| c | 133.70 |
| Molecules/asymm. unit | 1 |
| Resolution Range (Å) 1 | 50.0–2.55 (2.59–2.55) |
| Rsym (%) | 8.7 (77.0) |
| Observations | 161,860 |
| Unique reflections | 22,135 |
| Completeness (%) | 100.0 (100.0) |
| Redundancy | 7.3 (5.8) |
| <I/σ> | 23.6 (2.3) |
| Refinement | |
| Number of reflections | 22,124 |
| Rwork (%) | 20.9 (35.3) |
| Rfree2 | 26.0 (46.4) |
| Protein atoms | 3365 |
| Ca2+/H2O/SO42− atoms | 42 |
| Geometry (R.m.s.d.) | |
| Bond lengths (Å) | 0.012 |
| Bond angles (°) | 1.142 |
| Mean B (Å 2) | |
| Protein | 60.5 |
| Solvent | 51.5 |
| Calcium ions | 93.6 |
| Ramachandran plot 3 | |
| Favored | 93.6 |
| Allowed | 6.2 |
| Disallowed | 0.2 |
| MolProbity score 4 | 1.81 |
| PDB accession code | 6VRT |
| Antibody | CDR-L1 | CDR-L2 | CDR-L3 | Organism | Antigen | Metal |
| LT1002 | ITTTDIDDDMN | EGNILRP | LQSDNLPFT | Mouse | S1P | Ca2+ |
| Q425 | ITSTDIDDDMN | EGNTLRP | LQSDTLPLT | Mouse | CD4 | Ca2+/Ba2+ |
| 2C10 | ITNTDIDDDMN | EGNTLRP | LQSDNMPLT | Mouse | dsDNA | |
| EGFRvIII | ITSTDIDDDMN | EGNTLRP | LQSDNLPLF | Mouse | EGFR | |
| Gene | CDR-L1 | CDR-L2 | CDR-L3 | Organism | ||
| Igkv17-121 | ITSTDIDDDMN | EGNTLRP | LQSDNLP.. | Mouse | ||
| Igkv17-127 | ITSTDIDDDMN | EGNTLRP | LQSDNMP.. | Mouse | ||
| Igkv17-134 1 | THNTDIDDEMH | EGNTLHP | LQSGNMP.. | Mouse | ||
| Igkv5-2 | KASQDIDDDMN | EATTLVP | LQHDNFP.. | Human | ||
| Igkv17S1 | KTSTDIDDDMN | EGNTLRP | QQSDNVP.. | Rat | ||
| Vk5.4 | RAGQDIDDDMN | DATTLVS | LQHDNFP.. | Macaque |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farokhi, E.; Fleming, J.K.; Erasmus, M.F.; Ward, A.D.; Wu, Y.; Gutierrez, M.G.; Wojciak, J.M.; Huxford, T. Ion Binding Properties of a Naturally Occurring Metalloantibody. Antibodies 2020, 9, 10. https://0-doi-org.brum.beds.ac.uk/10.3390/antib9020010
Farokhi E, Fleming JK, Erasmus MF, Ward AD, Wu Y, Gutierrez MG, Wojciak JM, Huxford T. Ion Binding Properties of a Naturally Occurring Metalloantibody. Antibodies. 2020; 9(2):10. https://0-doi-org.brum.beds.ac.uk/10.3390/antib9020010
Chicago/Turabian StyleFarokhi, Elinaz, Jonathan K. Fleming, M. Frank Erasmus, Aaron D. Ward, Yunjin Wu, Maria G. Gutierrez, Jonathan M. Wojciak, and Tom Huxford. 2020. "Ion Binding Properties of a Naturally Occurring Metalloantibody" Antibodies 9, no. 2: 10. https://0-doi-org.brum.beds.ac.uk/10.3390/antib9020010