Unified Visual Working Memory without the Anterior Corpus Callosum

 , , , ,
, , , , 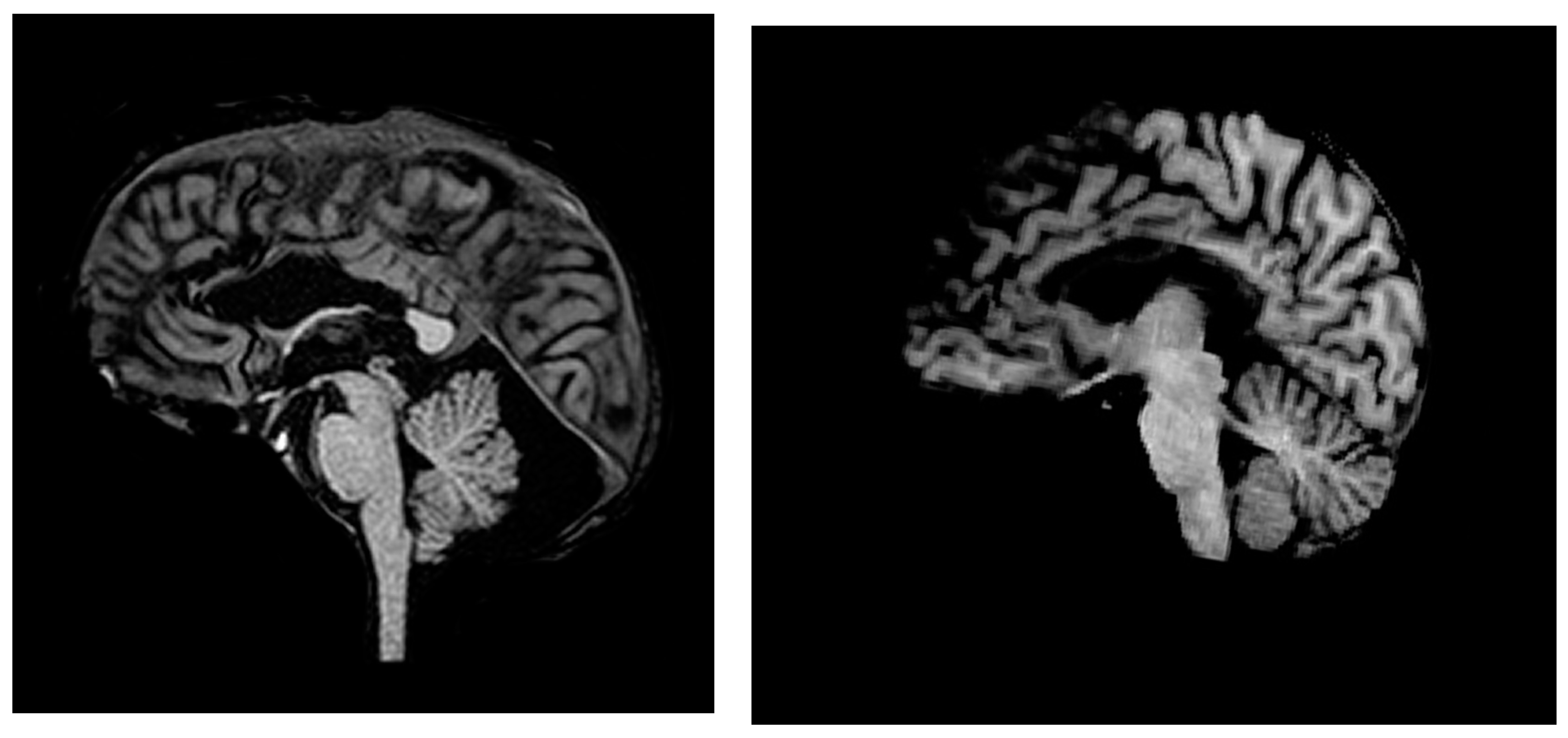{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Method
2.1. Participants
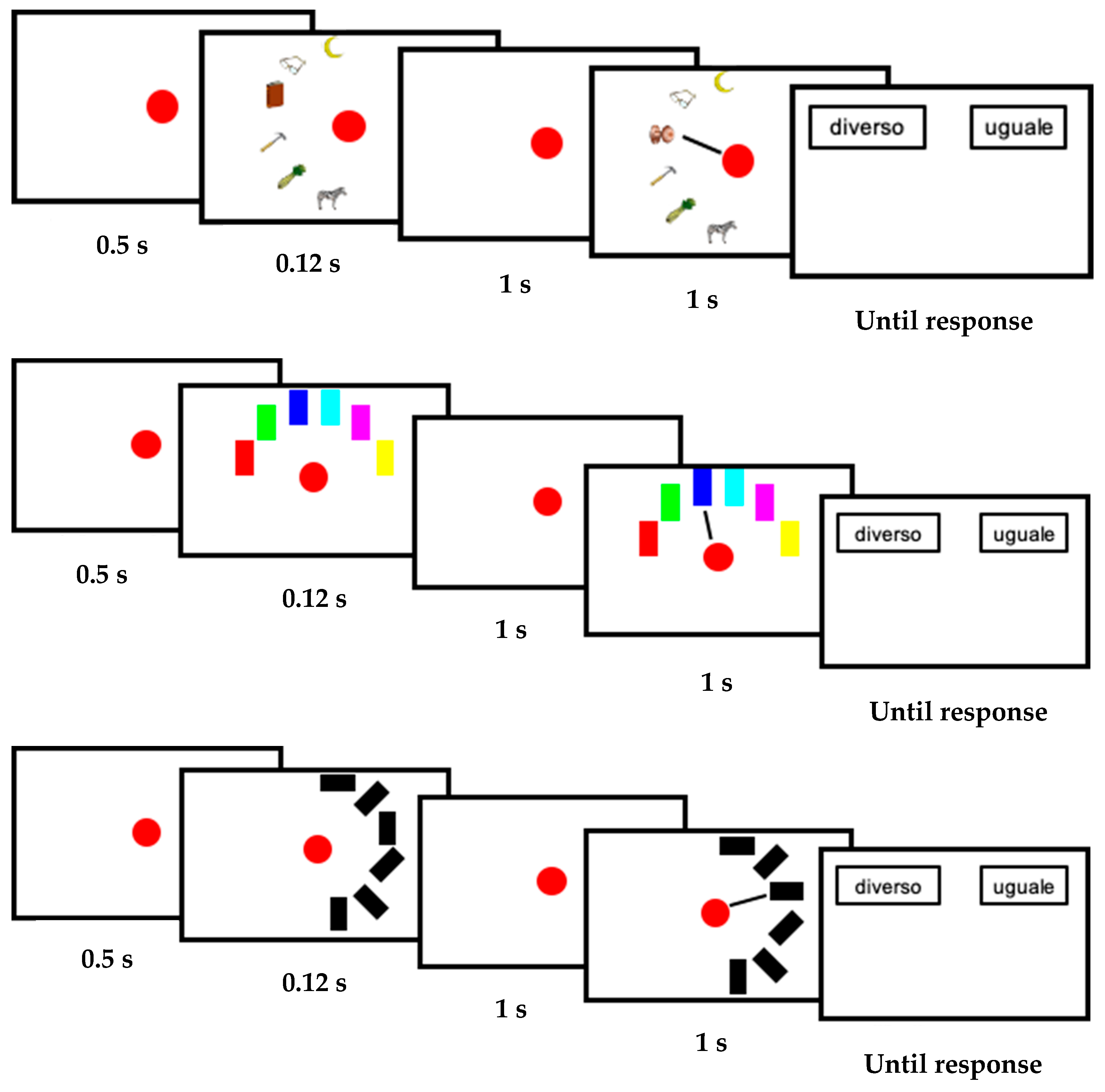
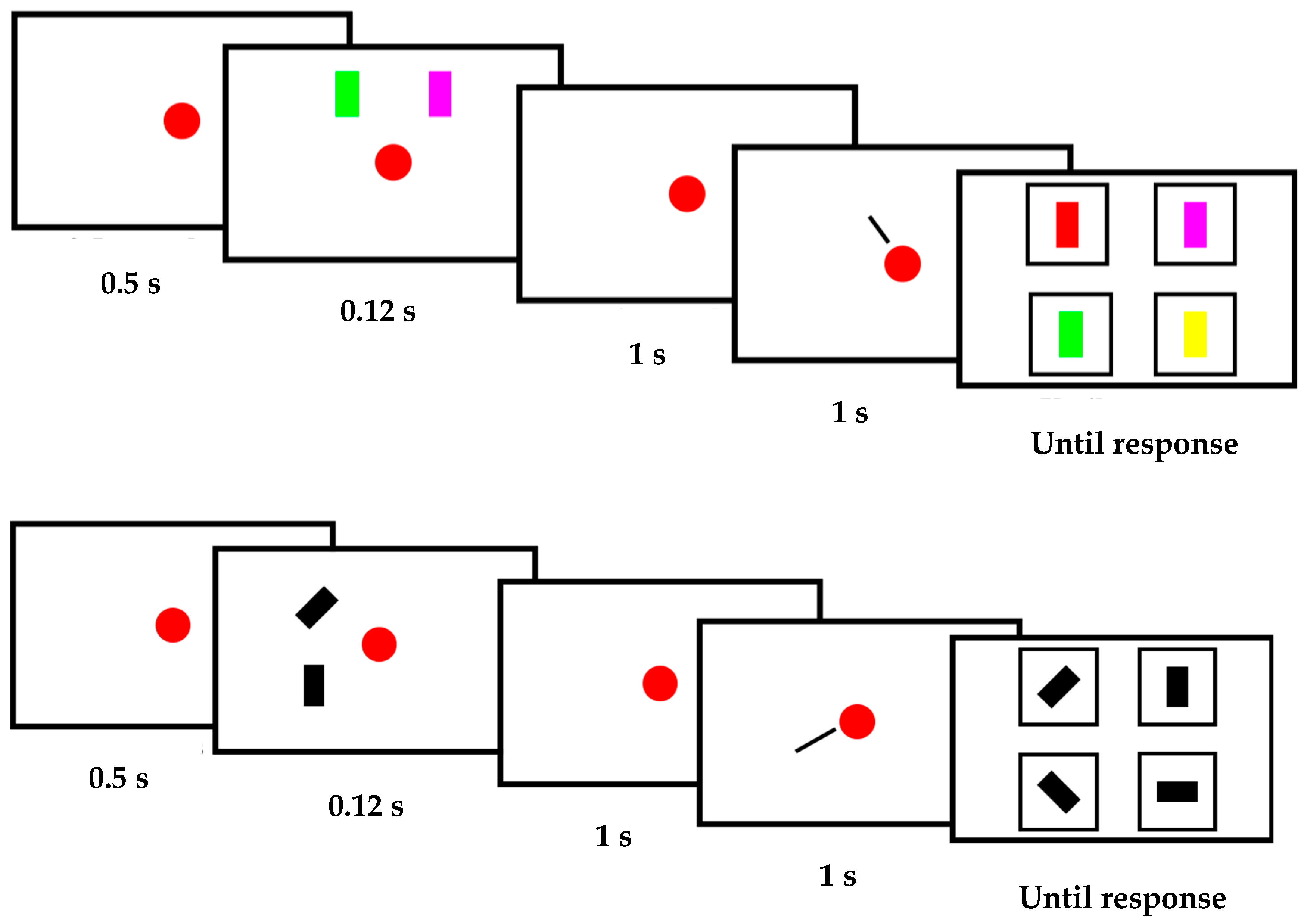
2.2. Stimuli and Procedure
2.3. Analysis
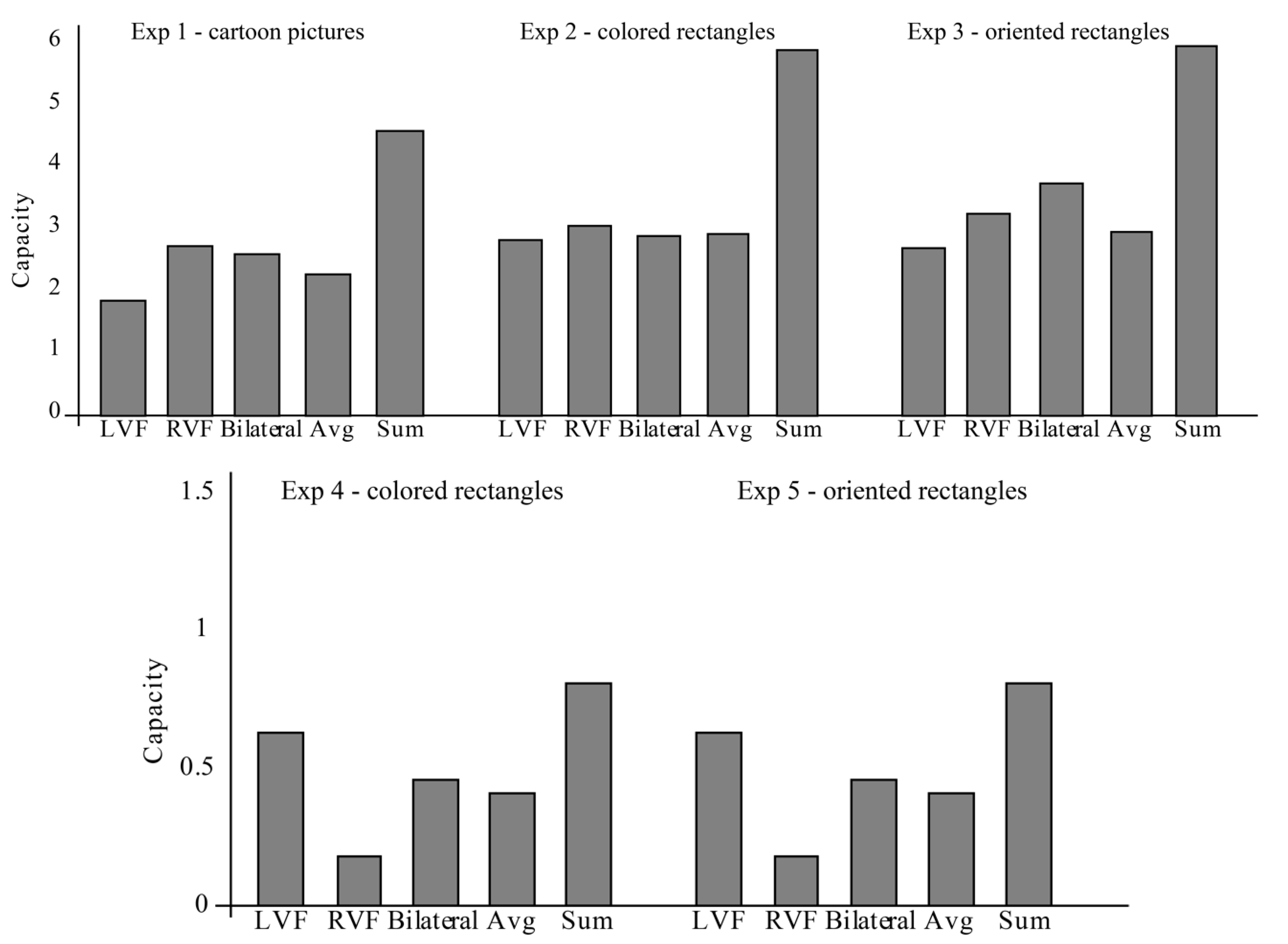
3. Results
4. Discussion
4.1. Visual Working Memory versus General Working Memory
4.2. Asymmetry of Visual Working Memory
4.3. Subcortical Unification
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ester, E.F.; Sprague, T.C.; Serences, J.T. Parietal and frontal cortex encode stimulus-specific mnemonic representations during visual working memory. Neuron 2015, 87, 893–905. [Google Scholar] [CrossRef] [Green Version]
- Miller, E.K.; Erickson, C.A.; Desimone, R. Neural mechanisms of visual working memory in prefrontal cortex of the macaque. J. Neurosci. Off. J. Soc. Neurosci. 1996, 16, 5154–5167. [Google Scholar] [CrossRef]
- Sligte, I.G.; Scholte, H.S.; Lamme, V.A. V4 activity predicts the strength of visual short-term memory representations. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 7432–7438. [Google Scholar] [CrossRef] [Green Version]
- Van Kerkoerle, T.; Self, M.W.; Roelfsema, P.R. Layer-specificity in the effects of attention and working memory on activity in primary visual cortex. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Arcaro, M.J.; McMains, S.A.; Singer, B.D.; Kastner, S. Retinotopic organization of human ventral visual cortex. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 10638–10652. [Google Scholar] [CrossRef] [Green Version]
- Cowey, A. Cortical maps and visual perception the grindley memorial lecture. Q. J. Exp. Psychol. 1979, 31, 1–17. [Google Scholar] [CrossRef]
- Penfield, W.; Boldrey, E. Somatic motor and sensory representation in the cerebral cortex of man as studied by electrical stimulation. Brain J. Neurol. 1937, 60, 389–443. [Google Scholar] [CrossRef]
- Wandell, B.A.; Brewer, A.A.; Dougherty, R.F. Visual field map clusters in human cortex. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 693–707. [Google Scholar] [CrossRef] [Green Version]
- Gazzaniga, M.S. Cerebral specialization and interhemispheric communication: Does the corpus callosum enable the human condition? Brain 2000, 123, 1293–1326. [Google Scholar] [CrossRef] [Green Version]
- Innocenti, G.M. General organization of callosal connections in the cerebral cortex. In Sensory-Motor Areas and Aspects of Cortical Connectivity; Springer: Boston, MA, USA, 1986; pp. 291–353. [Google Scholar]
- Wahl, M.; Lauterbach-Soon, B.; Hattingen, E.; Jung, P.; Singer, O.; Volz, S.; Klein, J.C.; Steinmetz, H.; Ziemann, U. Human motor corpus callosum: Topography, somatotopy, and link between microstructure and function. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 12132–12138. [Google Scholar] [CrossRef] [Green Version]
- Holt, J.L.; Delvenne, J. A bilateral advantage in controlling access to visual short-term memory. Exp. Psychol. 2014, 61, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umemoto, A.; Drew, T.; Ester, E.F.; Awh, E. A bilateral advantage for storage in visual working memory. Cognition 2010, 117, 69–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, M.M. Visual working memory as visual attention sustained internally over time. Neuropsychologia 2011, 49, 1407–1409. [Google Scholar] [CrossRef] [PubMed]
- Olivers, C.N.; Meijer, F.; Theeuwes, J. Feature-based memory-driven attentional capture: Visual working memory content affects visual attention. J. Exp. Psychol. Hum. Percept. Perform. 2006, 32, 1243. [Google Scholar] [CrossRef]
- Howe, P.D.; Cohen, M.A.; Pinto, Y.; Horowitz, T.S. Distinguishing between parallel and serial accounts of multiple object tracking. J. Vis. 2010, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Pinto, Y.; Howe, P.D.; Cohen, M.A.; Horowitz, T.S. The more often you see an object, the easier it becomes to track it. J. Vis. 2010, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Pinto, Y.; Scholte, H.S.; Lamme, V.A.F. Tracking moving identities: After attending the right location, the identity does not come for free. PLoS ONE 2012, 7, e42929. [Google Scholar] [CrossRef]
- Pylyshyn, Z.W.; Storm, R.W. Tracking multiple independent targets: Evidence for a parallel tracking mechanism. Spat. Vis. 1988, 3, 179–197. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, G.A.; Cavanagh, P. Independent resources for attentional tracking in the left and right visual hemifields. Psychol. Sci. 2005, 16, 637–643. [Google Scholar] [CrossRef]
- Cavanagh, P.; Alvarez, G.A. Tracking multiple targets with multifocal attention. Trends Cogn. Sci. 2005, 9, 349–354. [Google Scholar] [CrossRef]
- Drew, T.; Mance, I.; Horowitz, T.S.; Wolfe, J.M.; Vogel, E.K. A soft handoff of attention between cerebral hemispheres. Curr. Biol. 2014, 24, 1133–1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabri, M.; Polonara, G.; Mascioli, G.; Paggi, A.; Salvolini, U.; Manzoni, T. Contribution of the corpus callosum to bilateral representation of the trunk midline in the human brain: An fMRI study of callosotomized patients. Eur. J. Neurosci. 2006, 23, 3139–3148. [Google Scholar] [CrossRef] [PubMed]
- Pizzini, F.B.; Polonara, G.; Mascioli, G.; Beltramello, A.; Foroni, R.; Paggi, A.; Salvolini, U.; Tassinari, G.; Fabri, M. Diffusion tensor tracking of callosal fibers several years after callosotomy. Brain Res. 2010, 1312, 10–17. [Google Scholar] [CrossRef]
- Corballis, M.C.; Birse, K.; Paggi, A.; Manzoni, T.; Pierpaoli, C.; Fabri, M. Mirror-image discrimination and reversal in the disconnected hemispheres. Neuropsychologia 2010, 48, 1664–1669. [Google Scholar] [CrossRef] [PubMed]
- Sligte, I.G.; Scholte, H.S.; Lamme, V.A.F. Are there multiple visual short-term memory stores? PLoS ONE 2008, 3, e1699. [Google Scholar] [CrossRef] [PubMed]
- Sperling, G. The information available in brief visual presentations. Psychol. Monogr. Gen. Appl. 1960, 74, 1–29. [Google Scholar] [CrossRef]
- Thomas, L.E.; Irwin, D.E. Voluntary eyeblinks disrupt iconic memory. Percept. Psychophys. 2006, 68, 475–488. [Google Scholar] [CrossRef] [Green Version]
- Pinto, Y.; Sligte, I.G.; Shapiro, K.L.; Lamme, V.A.F. Fragile visual short-term memory is an object-based and location-specific store. Psychon. Bull. Rev. 2013, 20, 732–739. [Google Scholar] [CrossRef] [Green Version]
- Sligte, I.G.; Vandenbroucke, A.R.; Scholte, H.S.; Lamme, V. Detailed sensory memory, sloppy working memory. Front. Psychol. 2010, 1, 175. [Google Scholar] [CrossRef] [Green Version]
- Sligte, I.G.; Wokke, M.E.; Tesselaar, J.P.; Scholte, H.S.; Lamme, V.A. Magnetic stimulation of the dorsolateral prefrontal cortex dissociates fragile visual short-term memory from visual working memory. Neuropsychologia 2011, 49, 1578–1588. [Google Scholar] [CrossRef]
- Makovski, T. Are multiple visual short-term memory storages necessary to explain the retro-cue effect? Psychon. Bull. Rev. 2012, 19, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberauer, K.; Farrell, S.; Jarrold, C.; Lewandowsky, S. What limits working memory capacity? Psychol. Bull. 2016, 142, 758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, Y.; Vandenbroucke, A.R.; Otten, M.; Sligte, I.G.; Seth, A.K.; Lamme, V.A.F. Conscious visual memory with minimal attention. J. Exp. Psychol. Gen. 2017, 146, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Vandenbroucke, A.R.; Sligte, I.G.; Lamme, V.A. Manipulations of attention dissociate fragile visual short-term memory from visual working memory. Neuropsychologia 2011, 49, 1559–1568. [Google Scholar] [CrossRef]
- Pinto, Y.; Neville, D.A.; Otten, M.; Corballis, P.M.; Lamme, V.A.F.; de Haan, E.H.F.; Foschi, N.; Fabri, M. Split brain: Divided perception but undivided consciousness. Brain 2017, 140, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Pinto, Y.; de Haan, E.H.F.; Lamme, V.A.F. The split-brain phenomenon revisited: A single conscious agent with split perception. Trends Cogn. Sci. 2017, 21, 835–851. [Google Scholar] [CrossRef]
- Dundas, E.M.; Plaut, D.C.; Behrmann, M. The joint development of hemispheric lateralization for words and faces. J. Exp. Psychol. Gen. 2013, 142, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Hong, K.; Lee, S.K.; Kim, J.; Kim, K.; Nam, H. Visual working memory revealed by repetitive transcranial magnetic stimulation. J. Neurol. Sci. 2000, 181, 50–55. [Google Scholar] [CrossRef]
- Raymond, J.E.; Shapiro, K.L.; Arnell, K.M. Temporary suppression of visual processing in an RSVP task: An attentional blink? J. Exp. Psychol. Hum. Percept. Perform. 1992, 18, 849–860. [Google Scholar] [CrossRef]
- Shapiro, K.L.; Raymond, J.; Arnell, K. The attentional blink. Trends Cogn. Sci. 1997, 1, 291–296. [Google Scholar] [CrossRef]
- Giesbrecht, B.; Kingstone, A. Right hemisphere involvement in the attentional blink: Evidence from a split-brain patient. Brain Cogn. 2004, 55, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Ptito, A.; Brisson, B.; Dell’Acqua, R.; Lassonde, M.; Jolicœur, P. The attentional blink within and across the hemispheres: Evidence from a patient with a complete section of the corpus callosum. Biol. Psychol. 2009, 82, 64–69. [Google Scholar] [CrossRef]
- Luck, S.J.; Hillyard, S.A.; Mangun, G.R.; Gazzaniga, M.S. Independent attentional scanning in the separated hemispheres of split-brain patients. J. Cogn. Neurosci. 1994, 6, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J.; Hillyard, S.A.; Mangun, G.R.; Gazzaniga, M.S. Independent hemispheric attentional systems mediate visual-search in split-brain patients. Nature 1989, 342, 543–545. [Google Scholar] [CrossRef] [PubMed]
- de Haan, E.H.F.; Corballis, P.M.; Hillyard, S.A.; Marzi, C.A.; Seth, A.; Lamme, V.A.F.; Volz, L.; Fabri, M.; Schechter, E.; Bayne, T.; et al. Split-brain: What we know now and why this is important for understanding consciousness. Neuropsychol. Rev. 2020, 30, 224–233. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto, Y.; de Haan, E.H.F.; Villa, M.-C.; Siliquini, S.; Polonara, G.; Passamonti, C.; Lattanzi, S.; Foschi, N.; Fabri, M. Unified Visual Working Memory without the Anterior Corpus Callosum. Symmetry 2020, 12, 2106. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12122106
Pinto Y, de Haan EHF, Villa M-C, Siliquini S, Polonara G, Passamonti C, Lattanzi S, Foschi N, Fabri M. Unified Visual Working Memory without the Anterior Corpus Callosum. Symmetry. 2020; 12(12):2106. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12122106
Chicago/Turabian StylePinto, Yair, Edward H.F. de Haan, Maria-Chiara Villa, Sabrina Siliquini, Gabriele Polonara, Claudia Passamonti, Simona Lattanzi, Nicoletta Foschi, and Mara Fabri. 2020. "Unified Visual Working Memory without the Anterior Corpus Callosum" Symmetry 12, no. 12: 2106. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12122106