Variation and Correlations in Departures from Symmetry of Brain Torque, Humeral Morphology and Handedness in an Archaeological Sample of Homo sapiens


Abstract
:1. Introduction
2. Materials
3. Methods
4. Results
5. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trinkaus, E.; Churchill, S.E.; Ruff, C.B. Post-cranial robusticity in Homo II: Humeral bilateral asymmetry and bone plasticity. Am. J. Phys. Anthrop. 1994, 93, 1–34. [Google Scholar] [CrossRef]
- Balzeau, A.; Gilissen, E.; Grimaud-Hervé, D. Shared pattern of quantified endocranial shape asymmetries among anatomically modern humans, great apes and fossil hominins. PLoS ONE 2012, 7, e29581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, C.N.; Stock, J.T. Extreme mobility in the Late Pleistocene? Comparing limb biomechanics among fossil Homo, varsity athletes and Holocene foragers. J. Hum. Evol. 2013, 64, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.G.; Stock, J.T. Human variation in the periosteal geometry of the lower limb: Signatures of behaviour among human Holocene populations. In Reconstructing Mobility: Environmental, Behavioral, and Morphological Determinants; Carlson, K.J., Marchi, D., Eds.; Springer: New York, NY, USA, 2014; pp. 67–90. [Google Scholar]
- Goldberg, E.; Roediger, D.; Kucukboyaci, N.E.; Carlson, C.; Devinsky, O.; Kuzniecky, R.; Cash, S.; Thesen, T. Hemispheric asymmetries of cortical volume in the human brain. Cortex 2013, 49, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Tobias, P.V. The brain of Homo habilis: A new level of organization in cerebral evolution. J. Hum. Evol. 1987, 16, 741–761. [Google Scholar] [CrossRef]
- Grimaud-Hervé, D. L’évolution de l’encéphale chez Homo Erectus et Homo Sapiens: Exemples de l’Asie et de l’Europe; Cahiers de Paléoanthropologie, CNRS: Paris, France, 1997; 406p. [Google Scholar]
- Holloway, R.L.; Broadfield, D.C.; Yuan, M.S. The Human Fossil Record: Brain Endocasts, Paleoneurological Evidence; Wiley-Liss: New York, NY, USA, 2004; 315p. [Google Scholar]
- Balzeau, A.; Grimaud-Hervé, D.; Holloway, R.L.; Détroit, F.; Combès, B.; Prima, S. First description of the Cro-Magnon 1 endocast and study of brain variation an evolution in anatomically modern Homo Sapiens. BMSAP 2013, 25, 1–18. [Google Scholar] [CrossRef]
- Balzeau, A.; Gilissen, E.; Holloway, R.L.; Prima, S.; Grimaud-Hervé, D. Variations in size, shape and asymmetries of the third frontal convolution in hominids: Paleoneurological implications for hominin evolution and the origin of language. J. Hum. Evol. 2014, 76, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Halpern, M.E.; Güntürkün, O.; Hopkins, W.D.; Rogers, L.J. Lateralization of the vertebrate brain: Taking the side of model systems. J. Neurosci. 2005, 25, 10351–10357. [Google Scholar] [CrossRef] [Green Version]
- Hecaen, H.; Albert, M.L. Human Neuropsychology; John Wiley & Sons: New York, NY, USA, 1978; 509p. [Google Scholar]
- Bryden, M.P. Cerebral specialization: Clinical and experimental assessment. In Handbook of Neuropsychology; Boller, F., Grafman, J., Eds.; Elsevier: New York, NY, USA, 1988; Volume 1, pp. 143–159. [Google Scholar]
- Gazzaniga, M.S. On neural circuits and cognition. Neural Comput. 1994, 7, 1–12. [Google Scholar] [CrossRef]
- Gazzaniga, M.S. Principles of human brain organization derived from split-brain studies. Neuron 1995, 14, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Harrington, A. Unfinished business: Models of laterality in the nineteenth century. In Brain Asymmetry; Davidson, R.J., Hugdahl, K., Eds.; The MIT Press: Cambridge, MA, USA, 1995; pp. 3–27. [Google Scholar]
- Medina, Y.I.; Fernández, A.P.; Morris, D.M.; Canales-Rodríguez, E.J.; Haroon, H.; Garcia-Penton, L.; Augath, M.; García, L.G.; Logothetis, N.; Parker, G.; et al. Brain hemispheric structural efficiency and interconnectivity rightward asymmetry in human and nonhuman primates. Cereb. Cortex 2011, 21, 56–67. [Google Scholar] [CrossRef] [PubMed]
- LeMay, M. Morphological cerebral asymmetries of modern man, fossil man and nonhuman primate. Ann. N. Y. Acad. Sci. 1976, 280, 349–366. [Google Scholar] [CrossRef] [PubMed]
- LeMay, M. Asymmetries of the skull and handedness. J. Neurol. Sci. 1977, 32, 243–253. [Google Scholar] [CrossRef]
- Holloway, R.L.; De La Coste-Lareymondie, M.C. Brain endocast asymmetry in pongids and hominids: Some preliminary findings on the paleontology of cerebral dominance. Am. J. Phys. Anthrop. 1982, 58, 101–110. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Marino, L. Asymmetries in cerebral width in nonhuman primate brains as revealed by magnetic resonance imaging (MRI). Neuropsychologia 2000, 38, 493–499. [Google Scholar] [CrossRef]
- Pilcher, D.L.; Hammock, E.A.D.; Hopkins, W.D. Cerebral volumetric asymmetries in non-human primates: A magnetic resonance imaging study. Laterality 2001, 6, 165–179. [Google Scholar] [CrossRef] [Green Version]
- Good, C.D.; Johnsrude, I.; Ashburner, J.; Henson, R.N.; Friston, K.J.; Frackowiak, R. Cerebral asymmetry and the effects of sex and handedness on brain structure: A voxel-based morphometric analysis of 465 normal adult human brains. Neuroimage 2001, 14, 685–700. [Google Scholar] [CrossRef] [Green Version]
- Watkins, K.E.; Paus, T.; Lerch, J.; Zijdenbos, A.; Collins, D.L.; Neelin, P.; Taylor, J.; Worsley, K.; Evans, A. Structural asymmetries in the human brain: A voxel-based statistical analysis of 142 MRI scans. Cereb. Cortex 2001, 11, 868–877. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Taglialatela, J.P.; Meguerditchian, A.; Nir, T.; Schenker-Ahmed, N.M.; Sherwood, C.C. Gray matter asymmetries in chimpanzees as revealed by voxel-based morphometry. Neuroimage 2008, 42, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Holloway, R.L. Volumetric and asymmetry determinations on recent hominid endocasts: Spy I and II, Djebel Ihroud I, and the Salé Homo erectus specimens. Am. J. Phys. Anthrop. 1981, 5, 385–393. [Google Scholar] [CrossRef]
- Holloway, R.L. The Indonesian Homo erectus brain endocasts revisited. Am. J. Phys. Anthrop. 1981, 55, 503–521. [Google Scholar] [CrossRef]
- LeMay, M.; Kido, D.K. Asymmetries of the cerebral hemispheres on computed tomograms. J. Comput. Assist. Tomogr. 1978, 2, 471–476. [Google Scholar]
- LeMay, M.; Billig, M.S.; Geschwind, N. Asymmetries of the brains and skulls of nonhuman primates. In Primate Brain Evolution, Methods and Concepts; Falk, D., Armstrong, E., Eds.; Plenum Press: New York, NY, USA, 1976; pp. 263–277. [Google Scholar]
- Galaburda, A.M.; LeMay, M.; Kemper, T.L.; Geschwind, N. Right-left asymmetries in the brain. Science 1978, 199, 852–856. [Google Scholar] [CrossRef] [PubMed]
- Kertesz, A.; Black, S.E.; Polk, M.; Howell, J. Cerebral asymmetries on magnetic resonance imaging. Cortex 1986, 22, 117–127. [Google Scholar] [CrossRef]
- Falk, D.; Hildebolt, C.; Cheverud, J.; Vannier, M.W.; Helmkamp, R.C.; Konigsberg, L. Cortical asymmetries in frontal lobes of rhesus monkeys (Macaca mulatta). Brain Res. 1990, 512, 40–45. [Google Scholar] [CrossRef]
- Balzeau, A.; Gilissen, E. Endocranial shape asymmetries in Pan paniscus, Pan troglodytes and Gorilla gorilla assessed via skull based landmark analysis. J. Hum. Evol. 2010, 59, 54–69. [Google Scholar] [CrossRef]
- LeMay, M. Asymmetries of the brains and skulls of nonhuman primates. In Cerebral Lateralization in Nonhuman Species; Glick, S.D., Ed.; Academic Press: New York, NY, USA, 1985; pp. 233–245. [Google Scholar]
- Cain, D.P.; Wada, J.A. An anatomical asymmetry in the baboon brain. Brain Behav. Evol. 1979, 16, 222–226. [Google Scholar] [CrossRef]
- Cheverud, J.M.; Falk, D.; Hildebolt, C.; Moore, A.J.; Helmkamp, R.C.; Vannier, M.W. Heritability and association of cortical petalias in rhesus macaques (Macaca mulatta). Brain Behav. Evol. 1990, 35, 368–372. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Phillips, K.; Bania, A.; Calcutt, S.E.; Gardner, M.; Russell, J.; Schaeffer, J.; Lonsdorf, E.V.; Ross, S.R.; Schapiro, S.J. Hand preferences for coordinated bimanual actions in 777 great apes: Implications for the evolution of handedness in hominins. J. Hum. Evol. 2011, 60, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Bogart, S.L.; Mangin, J.-F.; Schapiro, S.J.; Reamer, L.; Bennett, A.J.; Pierre, P.J.; Hopkins, W.D. Cortical sulci asymmetries in chimpanzees and macaques: A new look at an old idea. Neuroimage 2012, 61, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Corballis, M.C.; Badzakova-Trajkov, G.; Häberling, I.S. Right hand, left brain: Genetic and evolutionary bases of cerebral asymmetries for language and manual action. WIREs Cogn. Sci. 2012, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ubelaker, D.H.; Zarenko, K.M. Can Handedness be Determined from Skeletal Remains? A Chronological Review of the Literature. J. Forensic Sci. 2012, 57, 1421–1426. [Google Scholar] [PubMed]
- Shaw, C.N. Is ‘hand preference’ coded in the hominin skeleton? An in-vivo study of bilateral morphological variation. J. Hum. Evol. 2011, 61, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Kubicka, A.M.; Nowaczewska, W.; Balzeau, A.; Piontek, J. Bilateral asymmetry of the humerus in Neandertals, Australian aborigines and medieval humans. Am. J. Phys. Anthrop. 2018, 167, 46–60. [Google Scholar] [CrossRef]
- Shaw, C.N.; Ryan, T.M. Does skeletal anatomy reflect adaptation to locomotor Patterns? Cortical and trabecular architecture in human and nonhuman Anthropoids. Am. J. Phys. Anthrop. 2012, 147, 187–200. [Google Scholar] [CrossRef]
- Ćwirko-Godycki, M. Early Medieval Burial Ground on the Lednicki Promontory. Materiały i Prace Antropologiczne 1956, 11, 1–56. [Google Scholar]
- Buikstra, J.E.; Ubelaker, D.H. Standards for Data Collection from Human Skeletal Remains; Arkansas Archeological Survey Research: Fayetteville, NC, USA, 1994; 44, 272pp. [Google Scholar]
- White, T.D.; Folkens, P.A. The Human Bone Manual; Elsevier: Oxford, UK, 2005. [Google Scholar]
- Wrzesińska, A.; Wrzesiński, J. Wczesnośredniowieczne cmentarzysko w Dziekanowicach, stanowisko 22. Studia Lednickie 1998, 5, 13–27. [Google Scholar]
- Makohonienko, M. Analiza palinologiczna zawartości ziemi z misy brązowej z wczesnośredniowiecznego cmentarzyska w Dziekanowicach, stanowisko 22. Studia Lednickie 2000, 6, 207–212. [Google Scholar]
- Makowiecki, D.; Tomek, T.; Bochenski, Z.M. Birds in Early Medieval Greater Poland: Consumption and Hawking. Int. J. Osteoarchaeol. 2014, 24, 358–364. [Google Scholar] [CrossRef]
- Profico, A.; Schlager, S.; Valoriani, V.; Buzi, C.; Melchionna, M.; Veneziano, A.; Raia, P.; Moggi-Cecchi, J.; Manzi, G. Reproducing the internal and external anatomy of fossil bones: Two new automatic digital tools. Am. J. Phys. Anthrop. 2018, 166, 979–986. [Google Scholar] [CrossRef]
- Profico, A.; Buzi, C.; Melchionna, M.; Veneziano, A.; Raia, P. Technical note: Endomaker, a new algorithm for fully automatic extraction of cranial endocasts and the calculation of their volumes. Am. J. Phys. Anthrop. 2020, accepted, in press. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.B. Body size, body shape, and long bone strength in modern humans. J. Hum. Evol. 2000, 38, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.B. Moment Macro for NIH Image and Image J. Available online: https://www.hopkinsmedicine.org/fae/mmacro.html (accessed on 6 February 2020).
- Lieberman, D.E.; Polk, J.D.; Demes, B. Predicting long bone loading from cross-sectional geometry. Am. J. Phys. Anthr. 2004, 123, 156–171. [Google Scholar] [CrossRef] [PubMed]
- Stock, J.T.; Shaw, C.N. Which measures of diaphyseal robusticity are robust? A comparison of external methods of quantifying the strength of long bone diaphyses to cross-sectional geometric properties. Am. J. Phys. Anthr. 2007, 134, 412–423. [Google Scholar] [CrossRef]
- Carlson, K.J.; Grine, F.E.; Pearson, O.M. Robusticity and Sexual Dimorphism in the Postcranium of Modern Hunter-Gatherers from Australia. Am. J. Phys. Anthr. 2007, 134, 9–23. [Google Scholar] [CrossRef]
- Davies, T.G.; Stock, J.T. The influence of relative body breadth on the diaphyseal morphology of the human lower limb. Am. J. Hum. Biol. 2014, 26, 822–835. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry analyses revisited. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 279–319. [Google Scholar]
- Palmer, A.R. From symmetry to asymmetry: Phylogenetic patterns of asymmetry variation in animals and their evolutionary significance. Proc. Natl. Acad. Sci. USA 1996, 93, 14279–14286. [Google Scholar] [CrossRef] [Green Version]
- Grubbs, F.E. procedures for detecting outlying observations in samples. Technometrics 1969, 11, 1–21. [Google Scholar] [CrossRef]
- Palmer, A.R. Fluctuating asymmetry analyses: A primer. In Developmental Instability: Its Origins and Evolutionary Implications; Markow, T.A., Ed.; Kluwer: Dordrecht, The Netherlands, 1994; pp. 335–364. [Google Scholar]
- Graham, J.H.; Emlen, J.M.; Freeman, D.C.; Leamy, L.J.; Kieser, J.A. Directional asymmetry and the measurement of developmental instability. Biol. J. Linn. Soc. 1998, 64, 1–16. [Google Scholar] [CrossRef]
- Van Valen, L. A study of fluctuating asymmetry. Evolution 1962, 16, 125–142. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry; Freeman: New York, NY, USA, 1995. [Google Scholar]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 223–225. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Palaeontological Statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Amrhein, V.; Greenland, S.; McShane, B. Scientists rise up against statistical significance. Nature 2019, 567, 305–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontulainen, S.; Sievänen, H.; Kannus, P.; Pasanen, M.; Vuori, I. Effect of long-term impact-loading on mass, size, and estimated strength of humerus and radius of female racquet-sports players: A peripheral quantitative computed tomography study between young and old starters and controls. J. Bone Miner. Res. 2003, 18, 352–359. [Google Scholar] [CrossRef]
- Galaburda, A.M. Anatomic basis of cerebral dominance. In Brain Asymmetry; Davidson, R.J., Hugdahl, K., Eds.; The MIT Press: Cambridge, MA, USA, 1995; pp. 51–73. [Google Scholar]
- Sládek, V.; Ruff, C.B.; Berner, M.; Holt, B.; Niskanen, M.; Schuplerová, E.; Hora, M. The impact of subsistence changes on humeral bilateral asymmetry in Terminal Pleistocene and Holocene Europe. J. Hum. Evol. 2016, 92, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Trinkaus, E.; Ruff, C.B. Diaphyseal Cross-sectional Geometry of Near Eastern Middle Palaeolithic Humans: The Humerus. J. Archaeol. Sci. 1999, 26, 173–184. [Google Scholar] [CrossRef]
- Pearson, O. Activity, climate, and postcranial robusticity. Curr. Anthropol. 2000, 41, 569–607. [Google Scholar] [CrossRef]
- Kubicka, A.M.; Lubiatowski, P.; Dlugosz, J.D.; Romanowski, L.; Piontek, J. Directional Asymmetry of Upper Limbs in a Medieval Population from Poland: A Combination of Linear and Geometric Morphometrics. Am. J. Hum. Biol. 2016, 28, 817–824. [Google Scholar] [CrossRef]
- Pearson, O.M. Postcranial remains and the origin of modern humans. Evol. Anthropol. 2000, 9, 229–247. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Parameter | Description |
|---|---|---|
| HBIOL | the biomechanical length of the humerus | measured from the most proximal point on the head to the most distal point on the lateral lip of the trochlea |
| J | the polar moment of area | indicates resistance to bending and torsional rigidity |
| %CA | percentage of cortical area | calculated as %CA = (CA/TA) × 100, it shows the distribution of cortical area versus total subperiosteal area |
| Imax/Imin | ratio of maximum to minimum second moments of area | calculated as Imax/Imin, ratio close to 1 indicates circular shape of the cross-section |
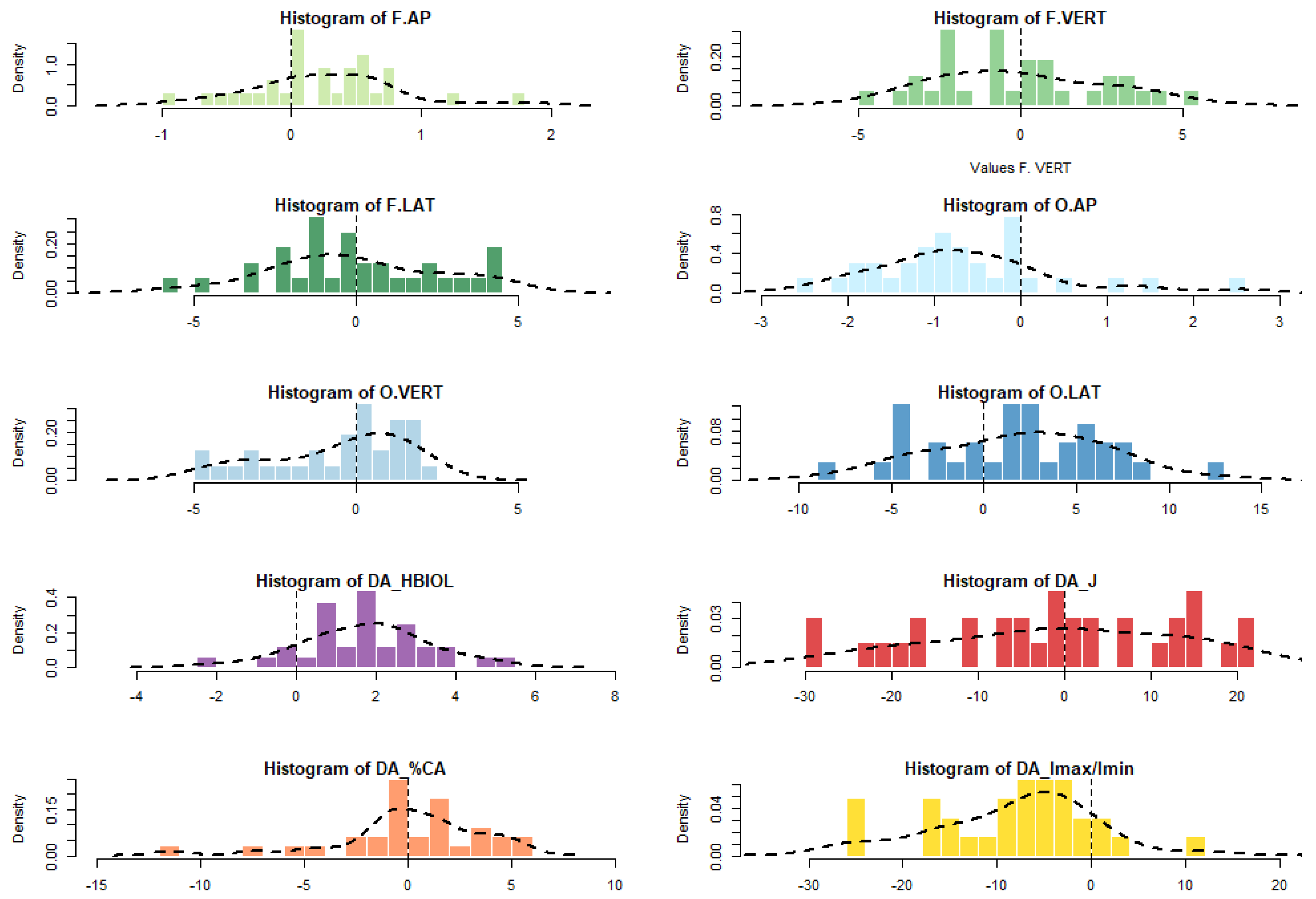
| F.AP | F.VERT | F.LAT | O.AP | O.VERT | O.LAT | HBIOL | J | %CA | Imax/Imin | |
|---|---|---|---|---|---|---|---|---|---|---|
| N | 32 | 32 | 32 | 32 | 31 | 32 | 32 | 32 | 31 | 32 |
| Min | −0.9 | −4.9 | −5.6 | −2.6 | −4.8 | −8.6 | −2.1 | −29.3 | −7.4 | −25.7 |
| 25 prcntil | −0.1 | −2.4 | −1.8 | −1.3 | −2.4 | −2.0 | 0.6 | −11.6 | −0.9 | −14.7 |
| Mean (R-L) | 0.2 | −0.1 | −0.1 | −0.7 | −0.6 | 1.9 | 1.7 | −1.3 | 0.4 | −7.6 |
| Median | 0.3 | −0.5 | −0.3 | −0.8 | 0.3 | 2.1 | 1.9 | −0.6 | 0.3 | −6.2 |
| 75 prcntil | 0.6 | 2.2 | 2.0 | −0.1 | 1.2 | 5.3 | 2.8 | 12.8 | 2.2 | −2.6 |
| Max | 1.8 | 5.4 | 4.5 | 2.5 | 2.5 | 12.7 | 5.2 | 21.6 | 5.3 | 12.3 |
| Sum | 7.9 | −2.9 | −1.6 | −21.1 | −17.3 | 59.9 | 55.3 | −41.7 | 13.2 | −241.8 |
| FA4a | 0.4 | 2.1 | 2.1 | 0.8 | 1.7 | 3.7 | 1.3 | 11.2 | 2.9 | 6.3 |
| Std. error | 0.1 | 0.5 | 0.5 | 0.2 | 0.4 | 0.8 | 0.3 | 2.6 | 0.5 | 1.6 |
| Variance | 0.3 | 6.7 | 6.6 | 1.1 | 4.5 | 22.6 | 2.5 | 208.6 | 8.5 | 82.0 |
| Stand. dev | 0.6 | 2.6 | 2.6 | 1.1 | 2.1 | 4.8 | 1.6 | 14.4 | 2.9 | 9.1 |
| ((R-L)/0) t | 2.5 | −0.2 | −0.1 | −3.5 | −1.4 | 2.2 | 6.2 | −0.5 | 0.1 | −5.4 |
| p | 3.4 × 10−2 | 0.84 | 0.91 | 4 × 10−3 | 0.15 | 3.3 × 10−2 | 3.9 × 10−6 | 0.61 | 0.93 | 2.9 × 10−5 |
| Skewness | 0.4 | 0.3 | 0.1 | 1.0 | −0.6 | −0.1 | −0.1 | −0.2 | −0.5 | −0.1 |
| Ts | 0.9 | 0.7 | 0.2 | 2.3 | −1.5 | −0.2 | −0.3 | −0.6 | −1.2 | −0.2 |
| P | > 0.05 | > 0.05 | > 0.05 | < 0.05 | > 0.05 | > 0.05 | > 0.05 | > 0.05 | > 0.05 | > 0.05 |
| Kurtosis | 1.2 | −0.7 | −0.4 | 1.7 | −0.8 | −0.2 | 0.3 | −0.8 | 0.7 | 0.4 |
| P | > 0.05 | > 0.05 | > 0.05 | < 0.05 | > 0.05 | > 0.05 | > 0.05 | > 0.05 | > 0.05 | > 0.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balzeau, A.; Ball-Albessard, L.; Kubicka, A.M. Variation and Correlations in Departures from Symmetry of Brain Torque, Humeral Morphology and Handedness in an Archaeological Sample of Homo sapiens. Symmetry 2020, 12, 432. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12030432
Balzeau A, Ball-Albessard L, Kubicka AM. Variation and Correlations in Departures from Symmetry of Brain Torque, Humeral Morphology and Handedness in an Archaeological Sample of Homo sapiens. Symmetry. 2020; 12(3):432. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12030432
Chicago/Turabian StyleBalzeau, Antoine, Lou Ball-Albessard, and Anna Maria Kubicka. 2020. "Variation and Correlations in Departures from Symmetry of Brain Torque, Humeral Morphology and Handedness in an Archaeological Sample of Homo sapiens" Symmetry 12, no. 3: 432. https://0-doi-org.brum.beds.ac.uk/10.3390/sym12030432