
Lateral Bias in Visual Working Memory

1
Department of Neurobiology and Biophysics, Life Science Center, Vilnius University, Sauletekio Ave. 7, LT-10257 Vilnius, Lithuania
2
Department of Psychology, University of Durham, Durham DH1 3LE, UK
*
Authors to whom correspondence should be addressed.
†
These authors have contributed equally to this work.
Symmetry 2022, 14(12), 2509; https://0-doi-org.brum.beds.ac.uk/10.3390/sym14122509
Submission received: 14 October 2022
/
Revised: 9 November 2022
/
Accepted: 22 November 2022
/
Published: 28 November 2022
(This article belongs to the Special Issue Individual Differences in Behavioral and Neural Lateralization)
Abstract
:Highlights
What are the main findings?
- Greater performance in men and women when letters presented in RVF.
- Smaller asymmetry related to better performance for women but not men.
What is the implication of the main finding?
- Lateral bias in visual working memory was independent from lateralization of lexical decisions.
Abstract
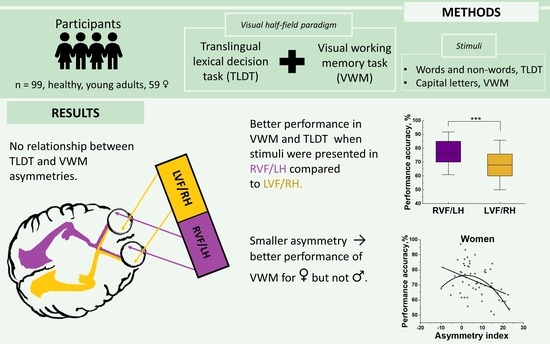
The present study aimed to evaluate functional cerebral asymmetries of visual working memory (VWM) in relation to language lateralization. The bilateral change detection paradigm with capital letters as stimuli and the translingual lexical decision task were used to assess VWM and language asymmetry, respectively, in a sample of 99 younger healthy participants (59 women). Participant attention was cued towards right or left visual half-field. For the VWM task, men and women were more accurate and faster when stimuli were presented in the right visual half-field compared to the left visual half-field. As expected, a significant right visual half-field advantage was demonstrated in the lexical decision task in performance accuracy (but not response time). The results also revealed no relationship between lateralization in VWM and lexical decision. VWM performance accuracy decreased significantly with increasing asymmetry. This relationship was significant for women, but not men. Taken together, the present study demonstrates that the lateral bias in visual working memory is independent from language lateralization, and less lateralized individuals perform better than individuals with larger asymmetries in both visual half-field tasks.
1. Introduction
Working memory (WM) has been defined as the temporary maintenance of information for the performance of a task in the (near) future, including manipulation and updating of the information [1]. WM is an important element of many complex cognitive (language, visuospatial performance, etc.) and emotional processes, is related to intelligence [2,3], and is crucial for adequate functioning in a variety of situations such as learning, reading, and problem solving [1,4].
1.1. Visual Change Detection Task: Classical Version and Bilateral Modification
The visual change-detection task [5,6] is a well-established paradigm for investigating different aspects of visual working memory (WVM) [7,8,9,10]. In the visual change-detection task, participants are briefly (100–500 ms) presented with a memory array of visual objects (e.g., colored bars in various orientations, squares, other shapes), which is followed by a short retention period (900–1000 ms) and a test probe. Participants are instructed to indicate whether the displayed test probe matches the previously viewed memory array or whether the probe has changed. One critical modification is the bilateral (or directional) version of the visual change-detection task [11]. This version of the task relies on the difference between an attended and non-attended side: the memory array is preceded by a centrally displayed arrow cue that points to the side of the screen from which participants encode visual stimuli that participants are asked to memorize, whereas they must ignore stimuli on the contralateral side.
The bilateral version of the change-detection VWM task allows to differentiate between ipsi- and contralateral visual pathways as stimuli presented to the right and left visual half-fields are dominantly processed by the left and right hemispheres, respectively. However, in the majority of studies using the bilateral change-detection task, contra- and ipsilateral brain responses and performance have been investigated without taking into account the visual half-field to which attention has been directed (e.g., [2,11,12,13,14]), and the studies that included visual half-field in the statistical design reported contradictory results [15,16,17,18,19,20,21,22,23,24,25]. Some of these studies reported faster and more accurate responses towards serially presented words in the RVF as compared to LVF, suggesting a RVF/LH advantage following lateralized encoding [18] and retrieval [19]. The lateralized color WM study [15] revealed LVH/RH advantage in accuracy for both encoding and retrieval. This advantage was most pronounced in a high compared to low memory load condition and was interpreted by the authors as RH superiority in encoding and retrieval of color stimuli when memory load is high, and a stronger LH involvement (i.e., reduced asymmetry) when memory load is low, probably due to a LH verbalization strategy when memorizing color stimuli. Notably, in this study, the LVF/RH advantage was found in 90% of men but only in 60% of women [15]. In an early psychophysical study [26], participants were asked to memorize either digits (verbal memory task) or digit positions (spatial memory task) presented for 80 ms in the right or left visual half-field. Similar to Clapp et al. [15], the results revealed visual half-field and sex/gender-specific findings: men recalled more digits (verbal memory) from the RVF than LVF, whereas women did not show any performance differences between visual half-fields. In addition, the recall of digit positions (i.e., spatial memory) did not differ as a function of the visual hemi-field despite the classical of a RH specialization for the spatial component of WM (e.g., [27,28]). More recent studies that administered the change detection task [16,17,21,22,23,25] did not find visual half-field differences in memory capacity, accuracy, or response time, but this might be partly due to the small sample sizes (n ≤ 16), which caused small statistical power in these studies.
1.2. Stimuli in Change Detection VWM Studies
All change detection VWM studies mentioned above used shapes as stimuli. In fact, we found only one change detection VWM study [29] that used spatially organized letters to assess WM performance in parallel to ERPs recording. However, alphanumerical stimuli (letters and numbers) are frequently used in other WM paradigms such as the N-back task [30], serial order task [31], Sternberg task [32], and in the studies investigating the phonological loop (storage system in WM) [33]. However, letters are problematic as they can be memorized verbally or as simple visual stimuli (similar to shapes) [29]. If letters are stored in WM as verbal information, the RVF/LH advantage might simply indicate the well-known RVF/LH advantage for language processing [34]. For example, in an fMRI WM study, Axmacher et al. [35] demonstrated that verbal (but not spatial) WM processing mainly depends on language dominance, i.e., the regions that were engaged in language functions were also more involved in verbal WM processing. In other words, the LVF/RVF advantage in WM processing might be confounded by verbal/non-verbal (spatial) stimulus type and the hemispheric language specialization of participants.
To take stimulus type as potential confounder into account when investigating VWM, the present study therefore administered both (i) a VWM task with lateralized presentation of capital letters, and (ii) the translingual lexical decision task (TLDT, [36,37]). The TLDT is an established visual half-field task for investigating the RVF/LH advantage in language lateralization [36,37]. The visual half-field technique is a simple, cost effective and well-established paradigm for investigating functional cerebral asymmetries in the visual domain (e.g., [38] for overview, but see also [34,39] for critical opinion). A large positive correlation of RVF/LH advantages in TLDT and VWM tasks would suggest a common left-lateralized processing mechanism for lexical decisions and letter storage in WM, and would supplement the idea that the laterality bias in letters processing in VWM depends on the language dominant hemisphere [35]. If, on the other hand, VHF advantages in both tasks were found to be unrelated, this would suggest that letters are stored and encoded as visual, not verbal stimuli. The results would also supplement previous findings of an ERP study, which found that the CDA (i.e., ERP component considered to be a load-dependent marker of VWM encoding and storage, [11]) was elicited by colored squares and letters, and its amplitude was load-dependent for both types of stimuli, suggesting usage of same neural mechanisms; that is, VWM in both tasks (instead of verbal WM for letters) [29].
1.3. Asymmetry–Performance Relationships
Brain lateralization has been found in human and non-human species, which leads many authors to conclude that it must be evolutionarily adaptive as it allows, for example, to perform multiple tasks simultaneously, avoids unnecessary duplications of functions, and increases processing speed due to reduction in interhemispheric communication (for reviews see [40,41,42]). In terms of cognitive performance, one can assume that larger functional cerebral asymmetries indicate more effective processing, and consequently better performance. However, results of studies investigating asymmetry–performance relationships in humans are inconsistent. Some studies revealed better performance in more lateralized participants [43,44,45,46,47,48], whereas others revealed the opposite [36,49,50,51,52]. Reasons why these inconsistencies occur are not fully understood. Factors contributing to these inconsistencies include variation between mental processes [53] and sensory modalities (e.g., [46,51]). For example, Boles et al. [53], aiming to evaluate asymmetry–performance relationships for different cognitive functions, assembled data from several studies that implemented visual half-field and dichotic listening tasks. The results demonstrated positive as well as negative asymmetry–performance relationships, which, according to the authors, depend on the age at which cerebral lateralization for different processes develop. The authors suggested that asymmetry–performance relationships are positive in early (auditory linguistic processing) and late (spatial positional processing) lateralizing processes, but negative for processes that reveal a laterality bias in the age between 5 and 11 years (e.g., spatial quantitative, visual lexical processing). In two independent studies, Hirnstein et al. [46,51] revealed U-shaped asymmetry–performance relationships that were negative for visually presented stimuli (word-matching and face-decision tasks) [51], but positive for dichotic listening [46]. In addition, individual differences, such as participants’ age [44,46,48,54,55] and sex/gender [46,48,50,51,56] have been shown to affect brain lateralization, and therefore could be relevant in the context of the asymmetry–performance relationship. For example, Hirnstein et al. [51] evaluated asymmetry–performance relationships in word-matching and face-decision tasks and found that pronounced lateralization caused less accurate and slower performance in word matching task for both men and women. However, in a face-decision task, women with strong left-hemispheric biases performed slower but more accurately, contrary to men who demonstrated an inverted U-shaped asymmetry–performance relationship. Plessen et al. [48] demonstrated in a large (n = 215) MRI study that sex significantly moderated not just asymmetries in cortical thickness but also the correlations of cortical thickness asymmetry with vocabulary scores and verbal working memory. Specifically, they found that a positive relationship between cortical thickness (especially along the right mesial surface of the brain) and performance was stronger in men than women. Nevertheless, other studies failed to find substantial differences between men and women in asymmetry–performance relationships [36,46,50,56,57]. To the best of our knowledge, there are no studies evaluating asymmetry–performance relationship for VWM.
1.4. Aims and Hypotheses of the Present Study
Taken together, there is conflicting evidence that does not allow to convincingly conclude that visual half-field differences in VWM exist. The assumption of an RVF advantage is almost entirely based on indirect evidence from the literature. It also remains unclear how potential laterality effects in VWM are related to performance and to what extent previous findings are confounded by interindividual differences such as participants’ sex/gender. In addition, it is unclear whether letters stored in WM use verbal or visual WM resources. Therefore, the aims of the current study were (i) to evaluate behavioral right vs. left visual half-field asymmetry using a bilateral visual change-detection VWM task, (ii) to assess the relationship between asymmetries for lexical decision and VWM, (iii) to investigate relationship between VWM asymmetry and performance, and finally (iv) to determine to what extent sex/gender influences VWM lateralization in VWM per se and the lateralization–performance relationships.
It was hypothesized that an RVF/LH advantage would emerge for both the VWM and TLDT task, and that language asymmetry would correlate positively with VWM asymmetry if the latter is processed in verbal memory. On the other hand, if letters are stored as visual, not linguistic stimuli, then VWM and TLDT asymmetry indices will not correlate. We expected that assessment of the VWM performance in parallel to the evaluation of the language lateralization in a relatively large sample, involving both males and females, would reveal the potential effect of language lateralization on letters storage in WM.
2. Methods
2.1. Participants
We recruited 112 participants (21.79 ± 2.45 years, 68 women, 44 men) via personal contact and class advertisement in Vilnius University Life Sciences Center (see sub-section Demographic questionnaire for more details). Aiming to avoid possible effect of age- [13], medication-, and neural system-related disorders, general inclusion criteria were younger adult age (18–35 years), no history of head trauma or neuropsychiatric disorders, and not taking any medication affecting the CNS. Due to neuropsychiatric disorders (depression, panic disorder, epilepsy, head trauma) data of 8 participants were excluded; consequently, data of 61 women and 43 men were used for further analyses. All participants had normal or corrected to normal vision. All participants were White, as other ethnicities are still relatively rare in Lithuania.
The sample size was estimated using G*Power [58]. The a priori power analysis for ANCOVA test recommended a sample size of 96 participants to achieve a large effect size (f = 0.4; ƞp2 = 0.14) with an α value of 0.05, and power (1 − β error probability) of 0.85.
Before the beginning of the experiment, participants signed the informed consent. The study was conducted in accordance with the guidelines of the declaration of Helsinki [59].
2.2. Demographic Questionnaire
Participants completed a self-reported questionnaire that assessed demographic information (age, sex, current health, and languages spoken). Although men (23.02 ± 2.48 years) were on average older than women (20.90 ± 2.04 years, t(102) = 4.77, p < 0.0001, Cohen’s d = 0.93), this difference was numerically small (i.e., 2.12 years of age) and unlikely to be relevant for the present study. Moreover, age did not correlate significantly with any sex/gender-related parameters (i.e., memory capacity and asymmetry indices of performance, all r ≤ 0.11, p ≥ 0.23). Consequently, participants’ ages were not included in further analyses. Ninety-nine participants were university students or had completed university with a BSc/MSc degree, 5 participants (2 women, 3 men) indicated having secondary education.
Although the impact of sex steroids was not the key focus of the present study, previous research indicated that sex hormones can affect functional brain asymmetries [30,60]. To rule out that the study sample was biased with respect to hormonal status of women, we added a self-reported questionnaire including questions about women’s menstrual cycle and hormonal contraceptive use. According to the questionnaire, 63 women were naturally cycling and three were hormonal contraceptive users. Among naturally cycling women, 41 reported that their cycles were regular (mean duration 24–35 days) and provided information about their cycle day during the experiment. Based on the self-report, 15 were in the early follicular, 15 women were in the late follicular, 6 women in the mid-luteal and 5 women in the late luteal phase of their menstrual cycle. It should be noted that questionnaire data can only provide a rough estimate of women’s cycle phase but suggests that female participants of the present study were heterogenous in their hormonal status, and therefore it is unlikely that their behavioral data were hormonally biased.
2.3. Handedness
Participants’ hand preference was estimated with the Lithuanian version of the Edinburgh handedness inventory [61]. The questionnaire consisted of 10 original activities suggested by Oldfield and the same response format as Oldfield’s [61] original measure was used. Participants had to indicate the hand they prefer to use for each activity by putting a single cross in the left or right column. If the preference to use a particular hand was so strong that the participant would never use the other hand unless it is necessary, two crosses were to be put in the preferred column. The laterality index (LI) was calculated using the following formula: LI = ((sum of right-hand crosses − sum of left-hand crosses)/(sum of all crosses)) × 100. The scores of the LI ranged from −100 to 100 (mean 54.37 ± SD 58.15). Negative values indicated left-handed (n = 13) and positive values indicated right-handed preference (n = 85), one person scored zero, indicating no hand preference. The mean hand preference for men was (44.16 ± 65.13), and for women was (63.47 ± 50.39, t(102) = 1.70, p = 0.092). Due to the numerical sex/gender differences in handedness, and similar to Hausmann et al. [36], handedness was used as covariate in the statistical design.
2.4. Language Lateralization
The current study adopted the well-established visual half-field task (i.e., translingual lexical decision task, TLDT, Figure 1A) and experimental design from two previous studies [36,37]. The TLDT was programmed in DMDX [62]. In each trial, two stimuli, either word(s) and/or non-word(s), were presented simultaneously to the right-visual field (RVF) and left-visual field (LVF). Sixteen lowercased words (agenda, alibi, aura, casino, film, gala, garage, jazz, jury, menu, radio, piano, snob, studio, taxi, virus) and 48 non-words were created using the pseudoword creator.
“Wuggy” [63] were used (see [36,37] for more details about word selection and [36] for the list of the word and non-word combinations. All stimuli were displayed in black Courier New font, lower case, size 12 on a computer screen with a white background. The stimulus eccentricity was between 2° and 5° of visual angle horizontally and 0.5° of visual angle vertically. Each trial started with a fixation cross presented for 1000 ms, followed by two word/non-word stimuli presented to the RVF and LVF for 100 ms. Participants had 2000 ms to decide and indicate by pressing a button on a keyboard, whether a meaningful word was presented to the LVF (left-sided button press with left index finger, “F” button), RVF (right-sided button press with right index finger, “J” button) or if there was no word on either side (by pressing the “space” button with the thumbs of both hands). If no answer was given, the next trial started.
In total, there were 256 trials; 64 trials when a meaningful word appeared in the LVF, 64 trials when a meaningful word appeared in the RVF, and 128 trials when no meaningful word appeared. After 128 trials participants took a self-paced break. Prior to the main experiment, participants performed 10 practice trials. The TLDT took approximately 20–30 min to complete.
2.5. VWM Task
The present study adopted the change detection paradigm [5,29] to measure visual working memory parameters (Figure 1B). All stimuli were presented in black Courier New font, in capital letters, on a gray (rgb 192; 192; 192) background. Participants were seated at an approximately 57 cm eye distance from the display, so that 1 cm corresponded to a 1° visual angle. Each trial started with a cue and a fixation cross presented for 100 ms. The cue was a black arrow (0.8° wide, 0.4° high) presented above a central fixation cross (0.2° × 0.2°) and pointed to either the LVF or RVF. After the cue, a memory sample array was presented for 500 ms, based on previous VWM studies [13,29,55,64]. Two of these studies used letters as stimuli and presented these for 500 ms [29,55], and the other two demonstrated better performance in VWM task with a presentation time of 500 ms than with a presentation time of 100 ms [13,64]. Each memory sample array consisted of a fixation cross and bilateral sets of three, four or five capital letters C, F, M, P, S, T, V, and K (approx. 0.34° visual degree wide and 0.4° visual degree high). Letters were randomly arranged in the left and right visual half-fields within imaginary circles with a radius of 2°, 3.8°, 5.5° visual degrees. The stimuli eccentricity was 1.6° visual degree for inner and 4.3° visual degree for outer letters. Participants were instructed to memorize the letters in the cued visual half-field. After the memory sample array, a fixation cross appeared for 1000 ms and was followed by a memory test array. The memory test array was identical to the memory sample display in 50% of the trials. One letter, on the cued side, was changed relative to the corresponding memory sample in the remaining 50% of the trials. The positions of letters were balanced between RVFs and LVFs; changes were also distributed equally on both sides and with respect to the cross of fixation. The memory test display was presented for 2000 ms or until response. Participants were instructed to press ‘F’ with a left index finger, if the letter change was detected and ‘J’ with a right index finger, if no change was detected. If no response was given, the next trial started. The duration of intertrial intervals varied between 2100–2900 ms.
The task consisted of 240 trials (80 trials consisting of three-letter stimuli, 80 trials with four letters, and 80 trials with five letters in each visual half-field). After 120 trials, participants took a self-paced break. Ten practice trials were performed before the main experiment. E-PRIME 2.0 software (Psychology Software Tools, USA) was used for stimuli presentation and data collection. The VWM task took approximately 30–35 min to complete.
2.6. General Procedure
The experiments were performed in a sound-proof and light-controlled room. All participants filled in the demographic and Edinburgh [61] questionnaires, received detailed tasks instructions. The TLDT was performed first, followed by approximately 15 min break and the VWM task. The whole session lasted approximately 90 min.
2.7. Data Analysis
The number of correct answers (ACC, %) and the response time of correct responses (RT, ms) were assessed for RVF and LVF. Sided and absolute asymmetry indices (AIs) were calculated. The scores of the sided AI take the direction of laterality bias into account and were calculated using formula: [(RVF − LVF)/(RVF + LVF)] × 100. Sided AIs range from −100 to 100. For clarity, AIs calculated from RT were inverted so that positive values always indicated RVF/LH advantage. Absolute AIs were calculated as absolute value of sided AIs and showed asymmetry magnitude, irrespective of whether the laterality bias was to the left or right, resulting in values between 0 and 100 [36]. Smaller AIs around 0 indicate less asymmetries whereas larger AIs around 100 indicate pronounced asymmetries.
Similar to Hausmann et al. (2019), we excluded data of participants based on individual VWM task performance by calculating thresholds above/below chance based on binomial tests [65]. Before exclusion there were 104 participants (63 women, 41 men). The number of trials for one visual half-field was 120. Accordingly, the number of 72 correctly identified targets (60%) was used as an upper threshold (for the visual half-field with the higher performance) and 48 correctly identified targets (40%) was used as a lower threshold (for the visual half-field with the lower performance). Eighty-two participants demonstrated RVF advantage, twenty LVF advantage and two performed equally for RVF vs. LVF. Data of five (all with RVF advantage) participants were excluded because performance for stimuli presented in their dominant VHF was not significantly above chance level. Consequently, data of 99 participants (59 women, 40 men) were analyzed.
For the main analyses, mixed ANCOVA and Bonferroni test were used for post hoc. The sphericity assumption for repeated measures was tested with Mauchly’s test. If sphericity assumption was violated, Greenhouse–Geisser correction was applied.
Pearson correlation analyses were performed to evaluate the relationships between performance parameters from VWM and TLDT. For relations between lateralization and performance, both linear and quadratic (only for sided AIs) regressions were calculated, as previous studies suggested U-shaped relationship between the degree of lateralization and performance when sided AIs were considered [51,53].
Data were analyzed using IBM SPSS20. This study’s design and its analysis were not pre-registered.
3. Results
3.1. VWM Performance Accuracy
The performance accuracy was calculated using a formula: performance accuracy (%, ACC) = (((p (hit) + p (correct rejection))/2) − p (false alarm)) × 100, where: p (hit) = hit number/20; p (false alarm) = false alarm number/20; p (correct rejection) = correct rejection/20; 20—number of trials per condition.
ACC of the VWM task was subjected to a 2 (LVF vs. RVF) × 3 (three vs. four vs. five letters) × 2 (male vs. female) mixed ANCOVA. Handedness and AIs calculated from accuracy in TLDT were included as covariates. The analysis revealed significant RVF advantage, F (1, 94) = 38.59, p < 0.0001, ƞp2 = 0.29, indicating more accurate performance when stimuli were presented in the RVF (76.3 ± 10.3 %, M ± SD) as compared to LVF (67.9 ± 11.9 %). The effect of the number of letters that participants needed to remember was significant too, F (2, 188) = 157.47, p < 0.0001, ƞp2 = 0.63. As expected, performance accuracy decreased with increasing number of stimuli (all p < 0.0001) (Figure 2). Post hoc tests revealed the significant RVF advantage in all three VWM conditions (RVF: three letters: 91.5 ± 8.1 %, four: 77.8 ± 14.2 %, five: 59.7 ± 17.5 %; LVF: three letters: 84.1 ± 11.2 %, four: 67.2 ± 17.6 %, five: 52.5 ± 17.6 %, all p < 0.05 (Figure 2)). No other main effect or interaction approached significance, all F ≤ 2.2, p ≥ 0.113.
3.2. VWM Response Time
RT (ms) of correct responses of the VWM task was subjected to the same mixed ANCOVA as used for ACC. Handedness and AIs calculated from RT in TLDT were included as covariates. In line with performance accuracy, the analysis revealed significant RVF advantage; F (1, 94) = 14.17, p < 0.0001, ƞ2p = 0.14, i.e., faster responses to stimuli presented in RVF (887 ± 156 ms) as compared to LVF (916 ± 161 ms). Moreover, the effect of the condition (number of letters) was significant, F (1.33, 124.9) = 90.40, p < 0.0001, ƞ2p = 0.49. As expected, the more letters participants needed to remember, the longer response time was, and all conditions differed significantly from each other: stimuli presented in RVF: three letters: 807 ± 158 ms, four − 902 ± 163 ms, five: 950 ± 171 ms; in LVF: three letters: 843 ± 164 ms, four: 931 ± 167 ms, five: 972 ± 174 ms (all p < 0.0001, Figure 3). The analysis also revealed tendency of interaction between the visual half-field and number of letters: F (1.99, 124.9) = 2.42, p = 0.092, ƞ2p = 0.03. Post hoc showed that difference between visual half-fields decreased with increase in memory load (three stimuli t (98) = −6.58, p < 0.0001; four stimuli t (98) = −4.07, p < 0.0001; five stimuli t (98) = −2.76, p = 0.007). No other main effect or interaction approached significance, all F ≤ 2.42, p ≥ 0.092.
3.3. VWM Capacity
Following formula was used for working memory capacity calculation: memory capacity = (((p (hit) − p (false alarm))/(1 − p (false alarm))) × set size, where: p (hit) = hit number/20; p (false alarm) = false alarm number/20; 20—number of trials per condition [10]. The memory capacity can potentially range between 0 and 5, indicating a number of stimuli participant can maintain in VWM.
Memory capacity was subjected to a 2 (LVF vs. RVF) × 2 (male vs. female) mixed ANCOVA with handedness and AIs calculated from ACC in TLDT as covariates. The ANCOVA revealed the significant visual half-field effect; F (1, 94) = 6.02, p = 0.016, ƞ2p = 0.06, indicating higher memory capacity when stimuli were presented in the RVF (3.05 ± 0.50) as compared to LVF (2.93 ± 0.53). Moreover, there was a significant interaction between sex and visual half-field; F (1, 94) = 4.43, p = 0.038, ƞ2p = 0.05. Post hoc t-tests revealed that women’s memory capacity was higher when words were presented in the RVF (3.1 ± 0.49) than in the LVF (2.87 ± 0.56, t (58) = 3.96, p < 0.0001), while men’s memory capacity did not differ between visual half-fields (RVF 3.01 ± 0.52, LVF 3.01 ± 0.48, t (39) = −0.06, p = 0.96) (Figure 4.). All other main effects and interactions did not approach significance, all F ≤ 1.50, p ≥ 0.22.
3.4. VWM Asymmetry Indices
Frequency analysis revealed that 81% participants showed RVF advantage (positive-sided AI values) for performance accuracy (X2 (1, 99) = 39.23, p < 0.0001), 71% for RT (X2 (1, 99) = 18.68, p < 0.0001) and only 56% for memory capacity (X2 (1, 99) = 1.22, p = 0.27) (Figure 5). These results are in line with the ANOVA and indicated that mean AIs, showing better performance in RVF condition, were not simply based on outliers.
3.5. TLDT Performance
In line with the previous TLDT studies [36,37], there was significant RVF/LH advantage F (1, 88) = 6.52, p = 0.01, ƞ2p = 0.07 in ACC. ACC was higher (Δ = 8.7 %) when words were presented to the RVF/LH (61.7 ± 13.9) than to the LVF/RH (53.0 ± 14.3). Nevertheless, it is worth noticing that the effect of visual half-field on ACC was much smaller in the present study that in both previous (ƞ2p = 0.44 [37] and ƞ2p = 0.26 [36]). Moreover, in contrast to the same previous studies [36,37], the visual half-field effect did not occur for RTs (F (1, 88) = 1.01, p = 0.317) in the present study (RT: RVF/LH 692 ± 129, LVF/RH 712 ± 109). Sex/gender has no significant effect on ACC or RT (all p ≤ 0.15). Moreover, in line with previous studies [36,37], the visual half-field effect did not interact significantly with sex/gender.
3.6. Relationship between TLDT and VWM Performance
Pearson correlation analysis, performed to evaluate relationships between TLDT AI and VWM AI, did not show relationship between sided AI of two tasks (r(89) = 0.02, p = 0.82) (Figure 6). In addition, the VWM AI was calculated separately for three, four, and five letter conditions and correlated with TLDT AI but did not reveal significant relationships (respectively: r(89) = −0.05, p = 0.61, r(89) = 0.18, p = 0.09, r(89) = −0.08, p = 0.48).
3.7. VWM Performance Comparison between Participants with Typical and Atypical TLDT Asymmetry
Several previous studies [35,47] presented an interesting idea about the effect of typical (left) vs. atypical (right) language dominance on other cognitive functions, including WM. The present study aimed to investigate the idea that language lateralization (as assessed by TLDT) might impact on the VWM of letters more directly by comparing VWM performance of participants with typical (left dominant) and atypical (non-left dominant) language lateralization. The analysis of asymmetry indices calculated for TLDT revealed that 63 (69.2%) participants demonstrated typical (RVF/LH) advantage, 24 (26.4%) participants demonstrated atypical (LVF/RH) advantage, and 4 (4.4%) showed no laterality bias in TLDT accuracy. From a subgroup of typical language lateralization, 24 participants were selected to match the 24 participants with atypical language lateralization as measured with TLDT. However, simple comparisons between both groups did not reveal any significant differences in VWM accuracy, response times or AIs between these two groups (all t ≤ 1.7, p ≥ 0.1).
3.8. Asymmetry–Performance Relationships in VWM
The present study investigated asymmetry–performance relationships for the whole sample and for women and men separately. The evaluation of sex/gender effect was encouraged by studies showing sex/gender differences in asymmetry–performance relationships [48,51]. Regression analyses (linear and quadratic) were performed for both sided and absolute AIs, with mean ACC, RT, and memory capacity (both collapsed for RVF and LVF) (Table 1). The regression analyses revealed that correlation coefficients were consistently negative across all measures (although not all correlations were statistically significant), indicating that larger asymmetries in VWM were related to lower VWM performance. The correlation coefficients were even larger for quadratic regression, which might indicate optimum performance when VWM asymmetry was rather small. This effect was only significant in women, not men.
4. Discussion
The results of our study showed an overall RVF advantage in a bilateral change-detection WM task with letters as stimuli, suggesting a dominance of the language-dominant left hemisphere in VWM. The RVF advantage was independent of participant’s sex/gender as well as handedness. Moreover, language lateralization as determined by TLDT was not related to VWM performance. In addition, smaller VWM asymmetry was related to more accurate VWM performance and higher memory capacity.
The RVF advantage in VWM is in agreement with other studies showing an RVF advantage for digit recall [26] and word processing [18,19]. It is also important to note that single letters have previously been shown to produce a secure RVF advantage in VHF tasks [66]. However, it is unlikely that the RVF advantage is simply due to the verbal nature of the task and the well-known left hemisphere’s dominance in processing verbal information [34,36,37] because the lexical decision asymmetry, determined by TLDT, was not related to VWM performance. Moreover, we did not find differences between participants showing typical (RVF/LH) and atypical (LVF/RH) language lateralization, as determined by TLDT, in VWM accuracy, response times, and asymmetry indices. This indicates that participants who showed larger RVF/LH advantage in lexical decision task were not necessarily the same participants who were more lateralized in the same direction in the VWM task. The findings of the previous electrophysiological WM study [29] showed that letters, despite their verbal associations, elicited the visual (not verbal) working memory-related ERP component. Similarly, the missing relationship between performance asymmetry in VWM and TLDT asymmetry in the present study suggests that letters were maintained/processed in visual, but not verbal working memory. In a fMRI study of visual memory, Sheremata et al. [23] showed that during VWM processing left hemisphere IPS was strongly biased for contralateral targets, but right hemisphere IPS exhibited effects for stimuli placed in both visual half-fields. Moreover, Mesulam [67] proposed the model of hemispheric specialization of visuospatial attention, which is an integral part of VWM (see e.g., [68,69]), suggesting that the LH is concerned almost exclusively with attention to the contralateral right hemifield, whereas the RH is capable of directing attention to both sides of space. Therefore, stimuli from RVF could be processed in VWM more efficiently because of projections to contra- as well as ipsilateral hemispheres.
Nevertheless, the RVF advantage in the presented study could potentially be explained by the left-to-right reading habit in our sample. It has been demonstrated that the higher resolution of perception and the allocation of visual attention in the RVF in skilled left-to-right readers [70,71] is absent in Hebrew (right-to-left) and bilingual readers [70] and in subjects with disturbed reading [71]. Therefore, the effect of habitual reading direction on visual perception and VWM cannot be fully ruled out.
4.1. Asymmetry–Performance Relationships
The investigation of asymmetry–performance relationship revealed a significant negative correlation between asymmetry indices and VWM task performance. The negative correlation was strongest between asymmetry indices and performance accuracy and was best described by an inverted U-shape curve (quadratic regression), i.e., the accuracy was lower when asymmetries were larger. The relationship between asymmetry indices and memory capacity or response time was weaker and mostly not significant but indicated the same direction interaction—lower asymmetry was related to faster responses and higher memory capacity. To the best of our knowledge there are no previous studies evaluating the asymmetry–performance relationship in a change detection VWM task. Nevertheless, our results are consistent with those showing a negative relationship between the degrees of lateralization and performance on various verbal tasks involving semantic language decision, vocabulary and verbal fluency [49], remembering of words [50], translingual decisions [36], and word-matching [51]. However, there are also data showing opposite direction asymmetry–performance relationships across different cognitive functions, e.g., better reading performance [43], higher verbal IQ scores, better visuospatial skills [44,45], and higher accuracy in a dichotic listening task [46] in more lateralized participants. Thus, it seems that the asymmetry–performance relationship could be both negative and positive and depends on methodological aspects (cognitive functions, tasks, methods of lateralization determination, etc.) as well as individual characteristics (age, handedness, sex, etc.).
4.2. Sex Effect
In line with previous studies (e.g., [36,72,73]), no consistent sex/gender difference in the degree of performance lateralization was found, with one exception—women’s, but not men’s memory capacity was higher when words were presented in the RVF than in the LVF. Absence of the sex/gender difference was opposite to two previous studies showing the small effect of interaction between sex and visual half-field in a word-matching task (RVF advantage in women, but not men) [51] and in a digits recall task (RVF advantage in men but not women) [26]. In the present study and in both previous studies, the sex/gender effect on performance lateralization was small and should be interpreted with caution. Nevertheless, the results of present study also revealed the sex/gender effect in asymmetry–performance analysis. Here, the negative relationship between asymmetry and performance occurring in a whole sample became even stronger when evaluated in women and much weaker and non-significant when evaluated in men. It could be that this is an accidental finding and lower correlation in the male group may be simply explained by the lower men number in our sample. Nevertheless, it is interesting that in a word-matching study [51] (where there were also fewer men than women), negative correlation between asymmetry and performance accuracy was also lower in men as compared to women. It has been suggested that the asymmetry–performance relationship might depend on cooperation between the hemispheres via corpus callosum [72,74]. Therefore, the microstructural organizational differences of the corpus callosum between men and women [75] might, at least partly, explain not only sex/gender differences in functional brain asymmetries [60,76], but also the slightly different asymmetry–performance relationship.
4.3. Limitations
There are a few methodological aspects that are important to consider when interpreting the results of the present study. First of all, this refers to the relatively long stimuli presentation time (500 ms), which is longer than the tachistoscopic stimulus presentation of 150–500 ms typical in VHF studies. However, the longer presentation time in VWM tasks is needed to ensure that the performance is above chance level [13,64], and 500 ms presentations were used in the only previous VWM change detection study that also used letters as stimuli and demonstrated that letters are stored visually but not verbally in WM [29]. Furthermore, the laterality bias has been demonstrated in studies where presentation times are several seconds long such as the emotional chimeric faces tasks and the greyscales task [77,78,79]. Nevertheless, future studies should use shorter stimulus presentations or control for eye movement with eye-tracking devices for visual half-field presentations longer than 200 ms. Second, the response format of the TLDT in the present study was identical to two previous studies [36,37], which used three manual response (i.e., left hand response for targets in the LVF, right hand response for targets in the RVF, and bimanual response when no target was presented). For consistency, the same response format was applied for the VWM change detection task with the only difference being that no bimanual response was required, which bares the risk of manual stimulus–response compatibility effects. However, we consider this risk to be relatively small as (a) no compatibility effects were found when piloting the tasks, and (b) the present study included left- and right-handers and no handedness effects were found. Third, laterality has been shown to be sex/gender sensitive [60,76,80] and although sex/gender differences were not in the focus of the present study, sex/gender of participants was recorded and included in the statistical analyses. Although sex/gender could have been better balanced across participants, the current results suggest that a much larger sample size would have been necessary to find significant sex/gender differences. Moreover, previous studies suggested that an appropriate analyses of sex/gender differences would require an analysis of participants’ hormonal status (e.g., [81,82,83,84,85]). Forth, the evidence of a non-existing correlation between TLDT and VWM performance should be taken with caution; a lack of association between two tasks could be partly confounded by low test-retest reliability of the tasks in this sample [86], as previous laterality studies have shown dynamic fluctuations in laterality patters due to, for example, fluctuations in mood and sex (e.g., [60,87]). Finally, the participants of the current study were white younger adults, with most of them being university students living in Lithuania. Therefore, absence of full demographic data regarding age, race/ethnicity, and socioeconomic status may limit generalizability.
5. Conclusions
In summary, the current study is one of the few studies using letters as stimuli in a lateralized change-detection VWM task. The results of the present study suggest RVF bias in VWM, which is independent of sex, handedness, and language processing. In addition, and to the best of our knowledge, this is the first study showing the negative asymmetry–performance relationship in visual WM. However, a deeper understanding of at which stage of the VWM process lateralization emerges would require a separation of the entire VWM process into sensory, cognitive (attention, memory encoding, and retrieval related) and motor subprocesses; for example, with ERP.
Author Contributions
R.G. (Ramune Griksiene): conceptualization; methodology; supervision; writing—original draft. R.G. (Rimante Gaizauskaite): methodology; software; investigation; data curation; formal analysis; visualization; writing—review and editing. I.P.: investigation; data curation; formal analysis. M.H.: conceptualization; methodology; software; supervision; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
Research reported in this publication was supported by European Social Fund (project No. 09.3.3-LMT-K-712-22-0170) under grant agreement with the Research Council of Lithuania.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nyberg, L.; Eriksson, J. Working Memory: Maintenance, Updating, and the Realization of Intentions. Cold Spring Harb. Perspect. Biol. 2015, 8, a021816. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Vogel, E.; Mayr, U.; Awh, E. Quantity, Not Quality: The Relationship between Fluid Intelligence and Working Memory Capacity. Psychon. Bull. Rev. 2010, 17, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unsworth, N.; Fukuda, K.; Awh, E.; Vogel, E.K. Working Memory Delay Activity Predicts Individual Differences in Cognitive Abilities. J. Cogn. Neurosci. 2015, 27, 853–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healy, A.F.; Jr, L.E.B. Foreign Language Learning: Psycholinguistic Studies on Training and Retention; Psychology Press: London, UK, 2013; ISBN 978-1-134-80766-6. [Google Scholar]
- Luck, S.J.; Vogel, E.K. The Capacity of Visual Working Memory for Features and Conjunctions. Nature 1997, 390, 279–281. [Google Scholar] [CrossRef]
- Phillips, W.A. On the Distinction between Sensory Storage and Short-Term Visual Memory. Percept. Psychophys. 1974, 16, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Allon, A.S.; Vixman, G.; Luria, R. Gestalt Grouping Cues Can Improve Filtering Performance in Visual Working Memory. Psychol. Res. 2019, 83, 1656–1672. [Google Scholar] [CrossRef] [PubMed]
- Awh, E.; Barton, B.; Vogel, E.K. Visual Working Memory Represents a Fixed Number of Items Regardless of Complexity. Psychol. Sci. 2007, 18, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Li, Y.; Gan, S.; Du, F. The Reliability of Estimating Visual Working Memory Capacity. Sci. Rep. 2019, 9, 1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouder, J.N.; Morey, R.D.; Morey, C.C.; Cowan, N. How to Measure Working Memory Capacity in the Change Detection Paradigm. Psychon. Bull. Rev. 2011, 18, 324–330. [Google Scholar] [CrossRef] [Green Version]
- Vogel, E.K.; Machizawa, M.G. Neural Activity Predicts Individual Differences in Visual Working Memory Capacity. Nature 2004, 428, 748–751. [Google Scholar] [CrossRef]
- Machizawa, M.G.; Driver, J. Principal Component Analysis of Behavioural Individual Differences Suggests That Particular Aspects of Visual Working Memory May Relate to Specific Aspects of Attention. Neuropsychologia 2011, 49, 1518–1526. [Google Scholar] [CrossRef] [PubMed]
- Sander, M.C.; Werkle-Bergner, M.; Lindenberger, U. Contralateral Delay Activity Reveals Life-Span Age Differences in Top-down Modulation of Working Memory Contents. Cereb. Cortex. 2011, 21, 2809–2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Chun, M.M. Dissociable Neural Mechanisms Supporting Visual Short-Term Memory for Objects. Nature 2006, 440, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Clapp, W.; Kirk, I.J.; Hausmann, M. Effects of Memory Load on Hemispheric Asymmetries of Colour Memory. Laterality 2007, 12, 139–153. [Google Scholar] [CrossRef]
- Cutini, S.; Scarpa, F.; Scatturin, P.; Jolicœur, P.; Pluchino, P.; Zorzi, M.; Dell’Acqua, R. A Hemodynamic Correlate of Lateralized Visual Short-Term Memories. Neuropsychologia 2011, 49, 1611–1621. [Google Scholar] [CrossRef]
- Diamantopoulou, S.; Poom, L.; Klaver, P.; Talsma, D. Visual Working Memory Capacity and Stimulus Categories: A Behavioral and Electrophysiological Investigation. Exp. Brain Res. 2011, 209, 501–513. [Google Scholar] [CrossRef] [Green Version]
- Evans, K.M.; Federmeier, K.D. The Memory That’s Right and the Memory That’s Left: Event-Related Potentials Reveal Hemispheric Asymmetries in the Encoding and Retention of Verbal Information. Neuropsychologia 2007, 45, 1777–1790. [Google Scholar] [CrossRef] [Green Version]
- Evans, K.M.; Federmeier, K.D. Left and Right Memory Revisited: Electrophysiological Investigations of Hemispheric Asymmetries at Retrieval. Neuropsychologia 2009, 47, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Machizawa, M.G.; Driver, J.; Watanabe, T. Gray Matter Volume in Different Cortical Structures Dissociably Relates to Individual Differences in Capacity and Precision of Visual Working Memory. Cereb. Cortex 2020, 30, 4759–4770. [Google Scholar] [CrossRef]
- McCollough, A.W.; Machizawa, M.G.; Vogel, E.K. Electrophysiological Measures of Maintaining Representations in Visual Working Memory. Cortex 2007, 43, 77–94. [Google Scholar] [CrossRef]
- Robitaille, N.; Grimault, S.; Jolicœur, P. Bilateral Parietal and Contralateral Responses during Maintenance of Unilaterally Encoded Objects in Visual Short-Term Memory: Evidence from Magnetoencephalography. Psychophysiology 2009, 46, 1090–1099. [Google Scholar] [CrossRef]
- Sheremata, S.L.; Bettencourt, K.C.; Somers, D.C. Hemispheric Asymmetry in Visuotopic Posterior Parietal Cortex Emerges with Visual Short-Term Memory Load. J. Neurosci. 2010, 30, 12581–12588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheremata, S.; Shomstein, S. Hemifield Asymmetries Differentiate VSTM for Single- and Multiple-Feature Objects. Atten. Percept. Psychophys. 2014, 76, 1609–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voytek, B.; Knight, R.T. Prefrontal Cortex and Basal Ganglia Contributions to Visual Working Memory. Proc. Natl. Acad. Sci. USA 2010, 107, 18167–18172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kail, R.V.; Siegel, A.W. Sex and Hemispheric Differences in the Recall of Verbal and Spatial Information. Cortex 1978, 14, 557–563. [Google Scholar] [CrossRef]
- Jonides, J.; Smith, E.E.; Koeppe, R.A.; Awh, E.; Minoshima, S.; Mintun, M.A. Spatial Working Memory in Humans as Revealed by PET. Nature 1993, 363, 623–625. [Google Scholar] [CrossRef] [Green Version]
- De Renzi, E.; Faglioni, P.; Previdi, P. Spatial Memory and Hemispheric Locus of Lesion. Cortex J. Devoted Study Nerv. Syst. Behav. 1977, 13, 424–433. [Google Scholar] [CrossRef]
- Rajsic, J.; Burton, J.A.; Woodman, G.F. Contralateral Delay Activity Tracks the Storage of Visually Presented Letters and Words. Psychophysiology 2019, 56, e13282. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.E.; Swearingen, J.; Corbly, C.R.; Curry, T.E.; Kelly, T.H. Influence of Estradiol on Functional Brain Organization for Working Memory. Neuroimage 2012, 59, 2923–2931. [Google Scholar] [CrossRef] [Green Version]
- Petrides, M.; Alivisatos, B.; Meyer, E.; Evans, A.C. Functional Activation of the Human Frontal Cortex during the Performance of Verbal Working Memory Tasks. Proc. Natl. Acad. Sci. USA 1993, 90, 878–882. [Google Scholar] [CrossRef]
- Sternberg, S. Memory-Scanning: Mental Processes Revealed by Reaction-Time Experiments. Am. Sci. 1969, 57, 421–457. [Google Scholar] [PubMed]
- Baddeley, A. Working Memory: Looking Back and Looking Forward. Nat. Rev. Neurosci. 2003, 4, 829–839. [Google Scholar] [CrossRef]
- Bishop, D.V.M. Cerebral Asymmetry and Language Development: Cause, Correlate, or Consequence? Science 2013, 340, 1230531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axmacher, N.; Bialleck, K.A.; Weber, B.; Helmstaedter, C.; Elger, C.E.; Fell, J. Working Memory Representation in Atypical Language Dominance. Hum. Brain Mapp. 2008, 30, 2032–2043. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, M.; Brysbaert, M.; van der Haegen, L.; Lewald, J.; Specht, K.; Hirnstein, M.; Willemin, J.; Barton, J.; Buchilly, D.; Chmetz, F.; et al. Language Lateralisation Measured across Linguistic and National Boundaries. Cortex 2019, 111, 134–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willemin, J.; Hausmann, M.; Brysbaert, M.; Dael, N.; Chmetz, F.; Fioravera, A.; Gieruc, K.; Mohr, C. Stability of Right Visual Field Advantage in an International Lateralized Lexical Decision Task Irrespective of Participants’ Sex, Handedness or Bilingualism. Laterality Asymmetries Body Brain Cogn. 2016, 21, 502–524. [Google Scholar] [CrossRef] [Green Version]
- Bourne, V.J. The Divided Visual Field Paradigm: Methodological Considerations. Laterality 2006, 11, 373–393. [Google Scholar] [CrossRef] [PubMed]
- Krach, S.; Chen, L.M.; Hartje, W. Comparison between Visual Half-Field Performance and Cerebral Blood Flow Changes as Indicators of Language Dominance. Laterality 2006, 11, 122–140. [Google Scholar] [CrossRef]
- Esteves, M.; Lopes, S.S.; Almeida, A.; Sousa, N.; Leite-Almeida, H. Unmasking the Relevance of Hemispheric Asymmetries—Break on through (to the Other Side). Prog. Neurobiol. 2020, 192, 101823. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Ströckens, F.; Ocklenburg, S. Brain Lateralization: A Comparative Perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef]
- Rogers, L.J. Brain Lateralization and Cognitive Capacity. Animals 2021, 11, 1996. [Google Scholar] [CrossRef]
- Chiarello, C.; Welcome, S.E.; Halderman, L.K.; Leonard, C.M. Does Degree of Asymmetry Relate to Performance? An Investigation of Word Recognition and Reading in Consistent and Mixed Handers. Brain Cogn. 2009, 69, 521–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everts, R.; Lidzba, K.; Wilke, M.; Kiefer, C.; Mordasini, M.; Schroth, G.; Perrig, W.; Steinlin, M. Strengthening of Laterality of Verbal and Visuospatial Functions during Childhood and Adolescence. Hum. Brain Mapp. 2009, 30, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Gotts, S.J.; Jo, H.J.; Wallace, G.L.; Saad, Z.S.; Cox, R.W.; Martin, A. Two Distinct Forms of Functional Lateralization in the Human Brain. Proc. Natl. Acad. Sci. USA 2013, 110, E3435–E3444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirnstein, M.; Hugdahl, K.; Hausmann, M. How Brain Asymmetry Relates to Performance–a Large-Scale Dichotic Listening Study. Front. Psychol. 2014, 4, 997. [Google Scholar] [CrossRef] [Green Version]
- Mellet, E.; Zago, L.; Jobard, G.; Crivello, F.; Petit, L.; Joliot, M.; Mazoyer, B.; Tzourio-Mazoyer, N. Weak Language Lateralization Affects Both Verbal and Spatial Skills: An FMRI Study in 297 Subjects. Neuropsychologia 2014, 65, 56–62. [Google Scholar] [CrossRef]
- Plessen, K.J.; Hugdahl, K.; Bansal, R.; Hao, X.; Peterson, B.S. Sex, Age, and Cognitive Correlates of Asymmetries in Thickness of the Cortical Mantle Across the Life Span. J. Neurosci. 2014, 34, 6294–6302. [Google Scholar] [CrossRef] [Green Version]
- Bartha-Doering, L.; Kollndorfer, K.; Kasprian, G.; Novak, A.; Schuler, A.-L.; Fischmeister, F.P.S.; Alexopoulos, J.; Gaillard, W.D.; Prayer, D.; Seidl, R.; et al. Weaker Semantic Language Lateralization Associated with Better Semantic Language Performance in Healthy Right-Handed Children. Brain Behav. 2018, 8, e01072. [Google Scholar] [CrossRef] [Green Version]
- Catani, M.; Allin, M.P.G.; Husain, M.; Pugliese, L.; Mesulam, M.M.; Murray, R.M.; Jones, D.K. Symmetries in Human Brain Language Pathways Correlate with Verbal Recall. Proc. Natl. Acad. Sci. USA 2007, 104, 17163–17168. [Google Scholar] [CrossRef] [Green Version]
- Hirnstein, M.; Leask, S.; Rose, J.; Hausmann, M. Disentangling the Relationship between Hemispheric Asymmetry and Cognitive Performance. Brain Cogn. 2010, 73, 119–127. [Google Scholar] [CrossRef]
- Làdavas, E.; Umiltà, C. Do Laterality Measures Relate to Speed of Response in Central Vision? Brain Cogn. 1983, 2, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Boles, D.B.; Barth, J.M.; Merrill, E.C. Asymmetry and Performance: Toward a Neurodevelopmental Theory. Brain Cogn. 2008, 66, 124–139. [Google Scholar] [CrossRef]
- Nagel, B.J.; Herting, M.M.; Maxwell, E.C.; Bruno, R.; Fair, D. Hemispheric Lateralization of Verbal and Spatial Working Memory during Adolescence. Brain Cogn. 2013, 82, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter-Lorenz, P.A.; Jonides, J.; Smith, E.E.; Hartley, A.; Miller, A.; Marshuetz, C.; Koeppe, R.A. Age Differences in the Frontal Lateralization of Verbal and Spatial Working Memory Revealed by PET. J. Cogn. Neurosci. 2000, 12, 174–187. [Google Scholar] [CrossRef]
- Boles, D.B. A Large-Sample Study of Sex Differences in Functional Cerebral Lateralization. J. Clin. Exp. Neuropsychol. 2005, 27, 759–768. [Google Scholar] [CrossRef]
- Leask, S.J.; Crow, T.J. Word Acquisition Reflects Lateralization of Hand Skill. Trends Cogn. Sci. 2001, 5, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- World Medical Association. World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human Subjects. JAMA 2013, 310, 2191–2194. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, M. Why Sex Hormones Matter for Neuroscience: A Very Short Review on Sex, Sex Hormones, and Functional Brain Asymmetries. J. Neurosci. Res. 2017, 95, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldfield, R.C. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Forster, K.I.; Forster, J.C. DMDX: A Windows Display Program with Millisecond Accuracy. Behav. Res. Methods Instrum. Comput. 2003, 35, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keuleers, E.; Brysbaert, M. Wuggy: A Multilingual Pseudoword Generator. Behav. Res. Methods 2010, 42, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Sheremata, S.L.; Somers, D.C. Role of Encoding Duration on Visual-Short Term Memory Capacity. J. Vis. 2008, 8, 1173. [Google Scholar] [CrossRef]
- Bortz, J.; Gustav, A.; Lienert; Boehnke, K. Verteilungsfreie Methoden in der Biostatistik; Springer-Lehrbuch; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 978-3-540-74706-2. [Google Scholar]
- Beaumont, J.G. Divided Visual Field Studies of Cerebral Organization; Academic Press: London, UK; New York, NY, USA, 1982; ISBN 978-0-12-084080-9. [Google Scholar]
- Mesulam, M. Functional Anatomy of Attention and Neglect: From Neurons to Networks. In The Cognitive and Neural Bases of Spatial Neglect; Oxford University Press: Oxford, UK, 2002; ISBN 978-0-19-850833-5. [Google Scholar]
- Desimone, R.; Duncan, J. Neural Mechanisms of Selective Visual Attention. Annu. Rev. Neurosci. 1995, 18, 193–222. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhao, C.; Guo, J.; Kong, Y.; Li, H.; Du, B.; Ding, Y.; Song, Y. Visual Working Memory Guides Spatial Attention: Evidence from Alpha Oscillations and Sustained Potentials. Neuropsychologia 2021, 151, 107719. [Google Scholar] [CrossRef]
- Kermani, M.; Verghese, A.; Vidyasagar, T.R. Attentional Asymmetry between Visual Hemifields Is Related to Habitual Direction of Reading and Its Implications for Debate on Cause and Effects of Dyslexia. Dyslexia 2018, 24, 33–43. [Google Scholar] [CrossRef]
- Rima, S.; Kerbyson, G.; Jones, E.; Schmid, M.C. Advantage of Detecting Visual Events in the Right Hemifield Is Affected by Reading Skill. Vis. Res. 2020, 169, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Hirnstein, M.; Hausmann, M.; Güntürkün, O. The Evolutionary Origins of Functional Cerebral Asymmetries in Humans: Does Lateralization Enhance Parallel Processing? Behav. Brain Res. 2008, 187, 297–303. [Google Scholar] [CrossRef]
- Jäncke, L.; Steinmetz, H. Interhemispheric Transfer Time and Corpus Callosum Size. Neuroreport 1994, 5, 2385–2388. [Google Scholar] [CrossRef] [PubMed]
- Lust, J.M.; Geuze, R.H.; Groothuis, A.G.G.; Bouma, A. Functional Cerebral Lateralization and Dual-Task Efficiency—Testing the Function of Human Brain Lateralization Using FTCD. Behav. Brain Res. 2011, 217, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Menzler, K.; Belke, M.; Wehrmann, E.; Krakow, K.; Lengler, U.; Jansen, A.; Hamer, H.M.; Oertel, W.H.; Rosenow, F.; Knake, S. Men and Women Are Different: Diffusion Tensor Imaging Reveals Sexual Dimorphism in the Microstructure of the Thalamus, Corpus Callosum and Cingulum. NeuroImage 2011, 54, 2557–2562. [Google Scholar] [CrossRef]
- Draca, S. Gender-Specific Functional Cerebral Asymmetries and Unilateral Cerebral Lesion Sequelae. Rev. Neurosci. 2010, 21, 421–425. [Google Scholar] [CrossRef]
- Innes, B.R.; Burt, D.M.; Birch, Y.K.; Hausmann, M. A Leftward Bias However You Look at It: Revisiting the Emotional Chimeric Face Task as a Tool for Measuring Emotion Lateralization. Laterality 2016, 21, 643–661. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, M.E.; Roberts, G.R. Can Free-Viewing Perceptual Asymmetries Be Explained by Scanning, Pre-Motor or Attentional Biases? Cortex 2002, 38, 113–136. [Google Scholar] [CrossRef] [PubMed]
- Smekal, V.; Burt, D.M.; Kentridge, R.W.; Hausmann, M. Emotion Lateralization in a Graduated Emotional Chimeric Face Task: An Online Study. Neuropsychology 2022, 36, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Hodgetts, S.; Hausmann, M. Sex/Gender Differences in Brain Lateralisation and Connectivity; Current Topics in Behavioral Neurosciences; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Bayer, U.; Kessler, N.; Güntürkün, O.; Hausmann, M. Interhemispheric Interaction during the Menstrual Cycle. Neuropsychologia 2008, 46, 2415–2422. [Google Scholar] [CrossRef]
- Bayer, U.; Erdmann, G. The Influence of Sex Hormones on Functional Cerebral Asymmetries in Postmenopausal Women. Brain Cogn. 2008, 67, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; De Tommaso, M.; Cianci, A.; Colacurci, N.; Rella, L.; Loiudice, L.; Cicinelli, M.V.; Livrea, P. Oral Contraceptive Therapy Modulates Hemispheric Asymmetry in Spatial Attention. Contraception 2011, 84, 634–636. [Google Scholar] [CrossRef]
- Hausmann, M. Hemispheric Asymmetry in Spatial Attention across the Menstrual Cycle. Neuropsychologia 2005, 43, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.; Hausmann, M.; Stoffers, B.; Vohn, R.; Kellermann, T.; Sturm, W. Estradiol Modulates Functional Brain Organization during the Menstrual Cycle: An Analysis of Interhemispheric Inhibition. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 13401–13410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouder, J.N.; Haaf, J.M. A Psychometrics of Individual Differences in Experimental Tasks. Psychon. Bull. Rev. 2019, 26, 452–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausmann, M.; Hodgetts, S.; Eerola, T. Music-Induced Changes in Functional Cerebral Asymmetries. Brain Cogn. 2016, 104, 58–71. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic figures of: (A) the translingual lexical decision task (TLDT); (B) the visual working memory task (VWM).
Figure 1.
Schematic figures of: (A) the translingual lexical decision task (TLDT); (B) the visual working memory task (VWM).

Figure 2.
Mean and standard deviations of performance accuracy in three, stimuli set sizes related, conditions of the VWM task. * p < 0.05, *** p < 0.0001.
Figure 2.
Mean and standard deviations of performance accuracy in three, stimuli set sizes related, conditions of the VWM task. * p < 0.05, *** p < 0.0001.

Figure 3.
Mean and standard deviations of response time in three, stimuli set size related, conditions of the VWM task. * p < 0.05, *** p < 0.0001.
Figure 3.
Mean and standard deviations of response time in three, stimuli set size related, conditions of the VWM task. * p < 0.05, *** p < 0.0001.

Figure 4.
Men’s and women’s memory capacity (means (in black dotted line) and participant values (in grey)) for right (RVF) vs. left (LVF) visual half-field. *** p < 0.001.
Figure 4.
Men’s and women’s memory capacity (means (in black dotted line) and participant values (in grey)) for right (RVF) vs. left (LVF) visual half-field. *** p < 0.001.

Figure 5.
Distribution of the sided (directional) asymmetry indices (AIsd) calculated from: (A) performance accuracy, (B) response time (inverted), and (C) memory capacity.
Figure 5.
Distribution of the sided (directional) asymmetry indices (AIsd) calculated from: (A) performance accuracy, (B) response time (inverted), and (C) memory capacity.

Figure 6.
Relationship between the sided (directional) asymmetry indices (AI) of VWM and TLDT tasks.
Figure 6.
Relationship between the sided (directional) asymmetry indices (AI) of VWM and TLDT tasks.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The linear and quadratic regressions between sided and absolute asymmetry indices (AIs) and overall (collapsed for LVF and RVF) VWM performance accuracy, response time, and memory capacity for all participants and across both sexes. Significant relationships (p ≤ 0.01, after correction for multiple comparisons) are shown in bold. Negative r values in quadratic regressions indicate inverted U-shape relationships. Optimal AI indicate which AIs were associated with optimal performance.
Table 1.
The linear and quadratic regressions between sided and absolute asymmetry indices (AIs) and overall (collapsed for LVF and RVF) VWM performance accuracy, response time, and memory capacity for all participants and across both sexes. Significant relationships (p ≤ 0.01, after correction for multiple comparisons) are shown in bold. Negative r values in quadratic regressions indicate inverted U-shape relationships. Optimal AI indicate which AIs were associated with optimal performance.
| Accuracy r (Optimal AI) | Memory Capacity r (Optimal AI) | Response Time r (Optimal AI) | |
|---|---|---|---|
| Sided (directional)AIs | |||
| Linear regression | |||
| All participants | −0.29 ** | −0.17 # | 0.005 |
| Women | −0.40 ** | −0.26 # | 0.004 |
| Men | −0.13 | −0.06 | 0.027 |
| Quadratic regression | |||
| All participants | −0.41 (1.61) *** | −0.25 (1.82) * | −0.13 (−1.09) |
| Women | −0.51 (2.36) *** | −0.26 (−6.67) | −0.18 (−2.55) |
| Men | −0.29 (2.38) | −0.28 (4.88) | −0.07 (−1.09) |
| Absolute AIs | |||
| Linear regression | |||
| All participants | −0.39 *** | −0.25 ** | −0.10 |
| Women | −0.49 *** | −0.29 * | −0.12 |
| Men | −0.21 | −0.20 | −0.07 |
Note: AIs—asymmetry indices, #—p ≤ 0.1, *—p ≤ 0.05, **—p ≤ 0.01, ***—p ≤ 0.001.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grikšienė, R.; Gaizauskaite, R.; Pretkelyte, I.; Hausmann, M. Lateral Bias in Visual Working Memory. Symmetry 2022, 14, 2509. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14122509
AMA Style
Grikšienė R, Gaizauskaite R, Pretkelyte I, Hausmann M. Lateral Bias in Visual Working Memory. Symmetry. 2022; 14(12):2509. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14122509
Chicago/Turabian StyleGrikšienė, Ramunė, Rimante Gaizauskaite, Indre Pretkelyte, and Markus Hausmann. 2022. "Lateral Bias in Visual Working Memory" Symmetry 14, no. 12: 2509. https://0-doi-org.brum.beds.ac.uk/10.3390/sym14122509
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.