

Structure Evolution of CaCO3 Precipitates Formed during the Bacillus cereus Induced Biomineralization
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
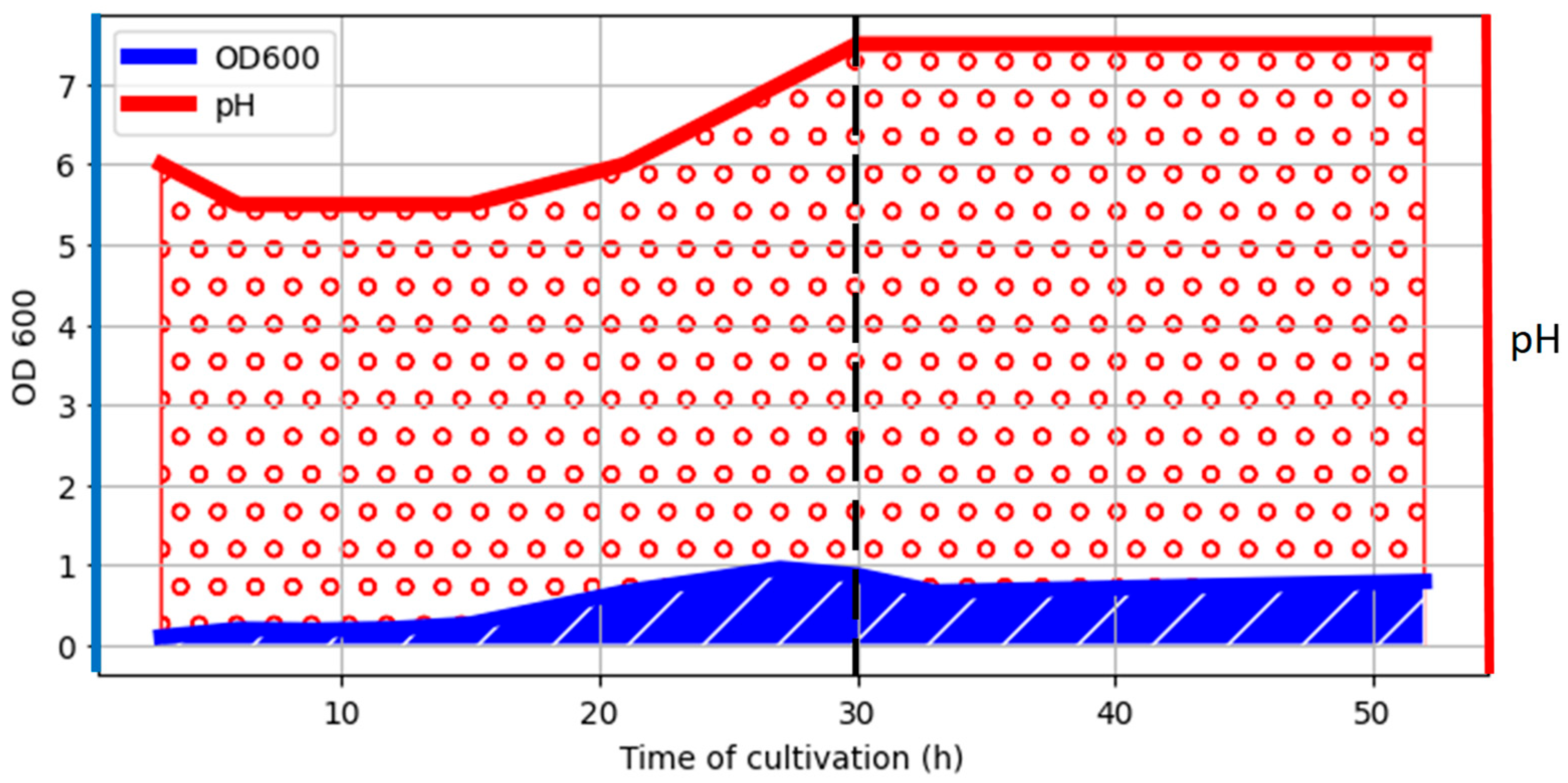
2.1. Bacterium Growth and Sample Preparations
2.2. Cell-Free Experiment
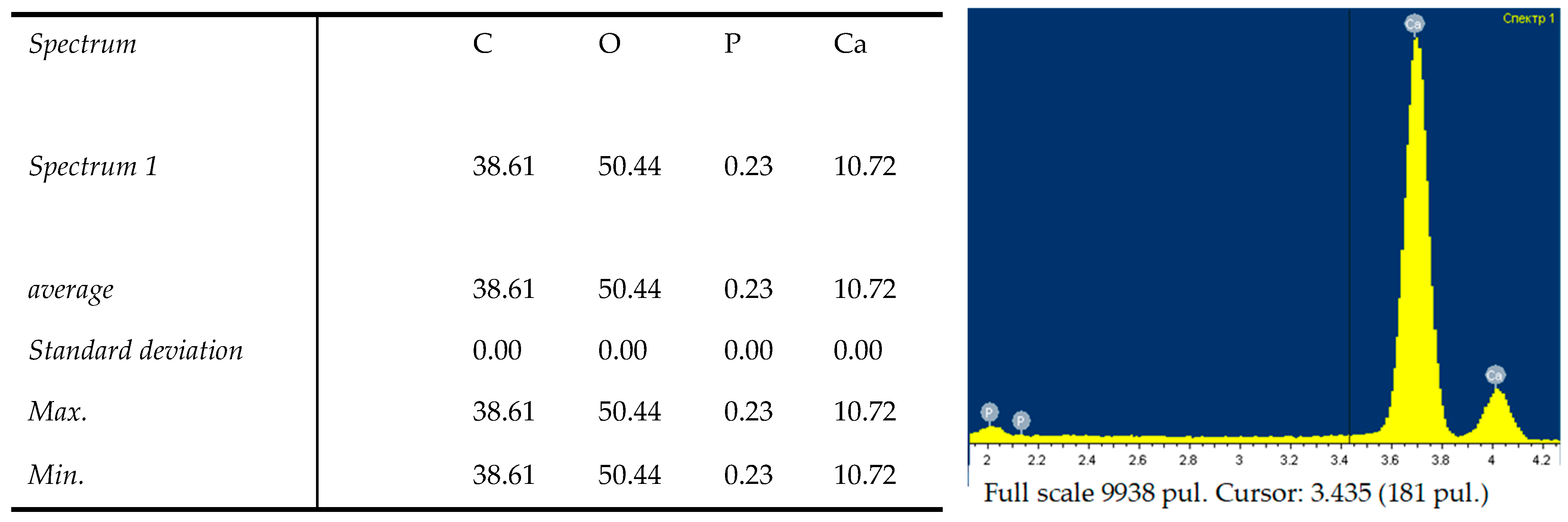
2.3. Scanning Electron Microscopy (SEM) and Energy-Dispersive X-ray (EDX) Analyses
2.4. XRD and FTIR Analyses
2.5. SAXS and SANS Analyses
3. Results and Discussion
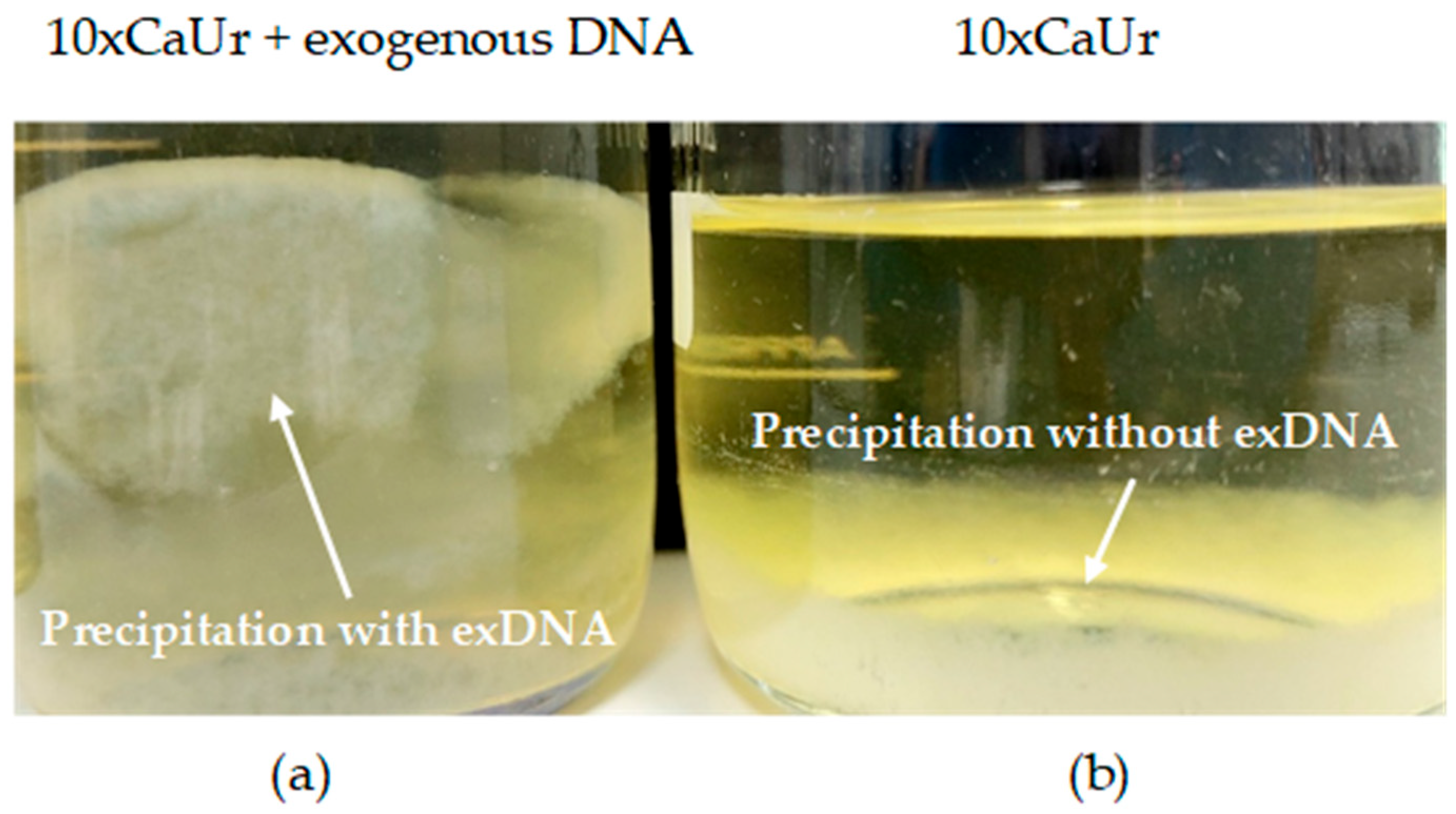
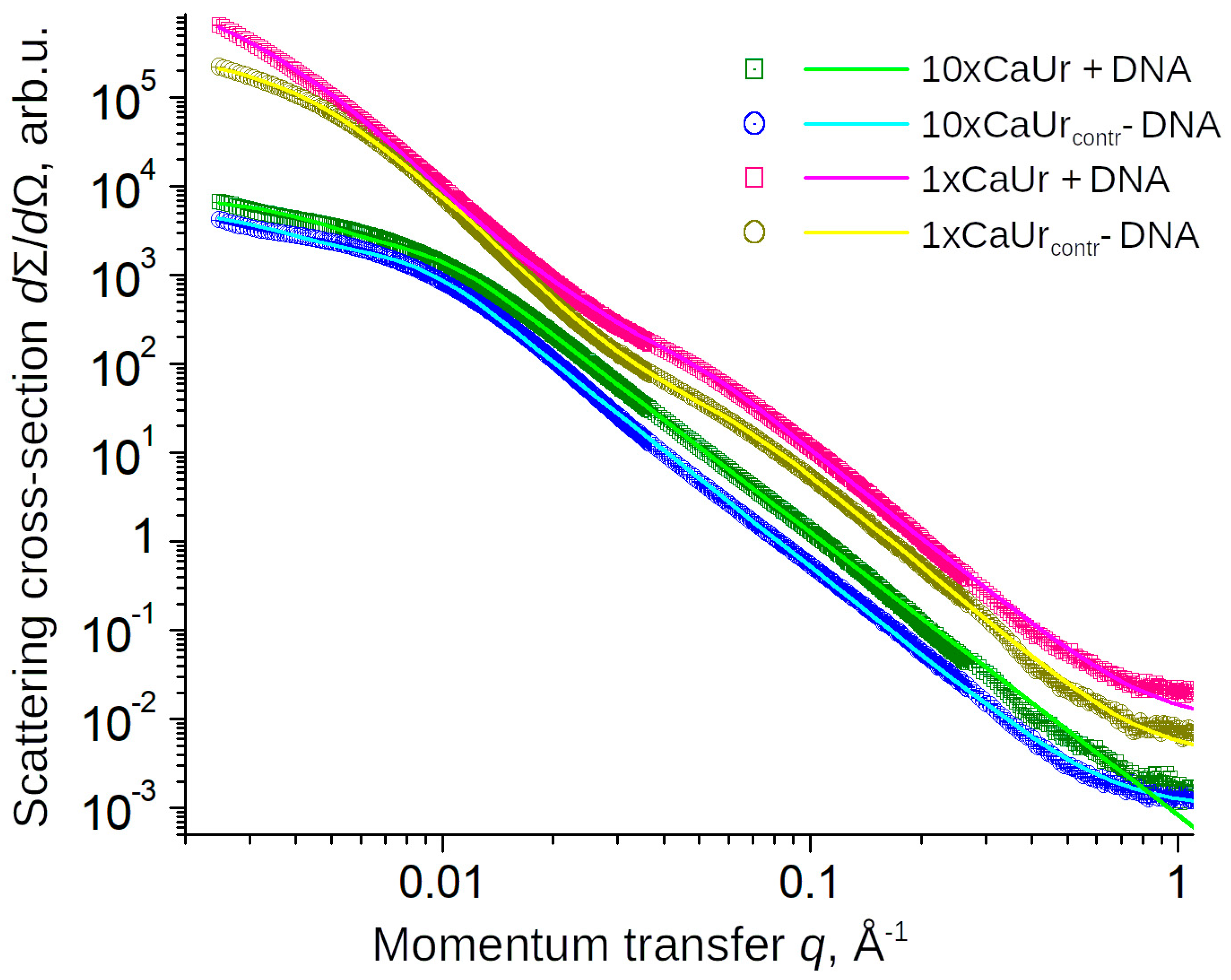
3.1. Cell-Free System
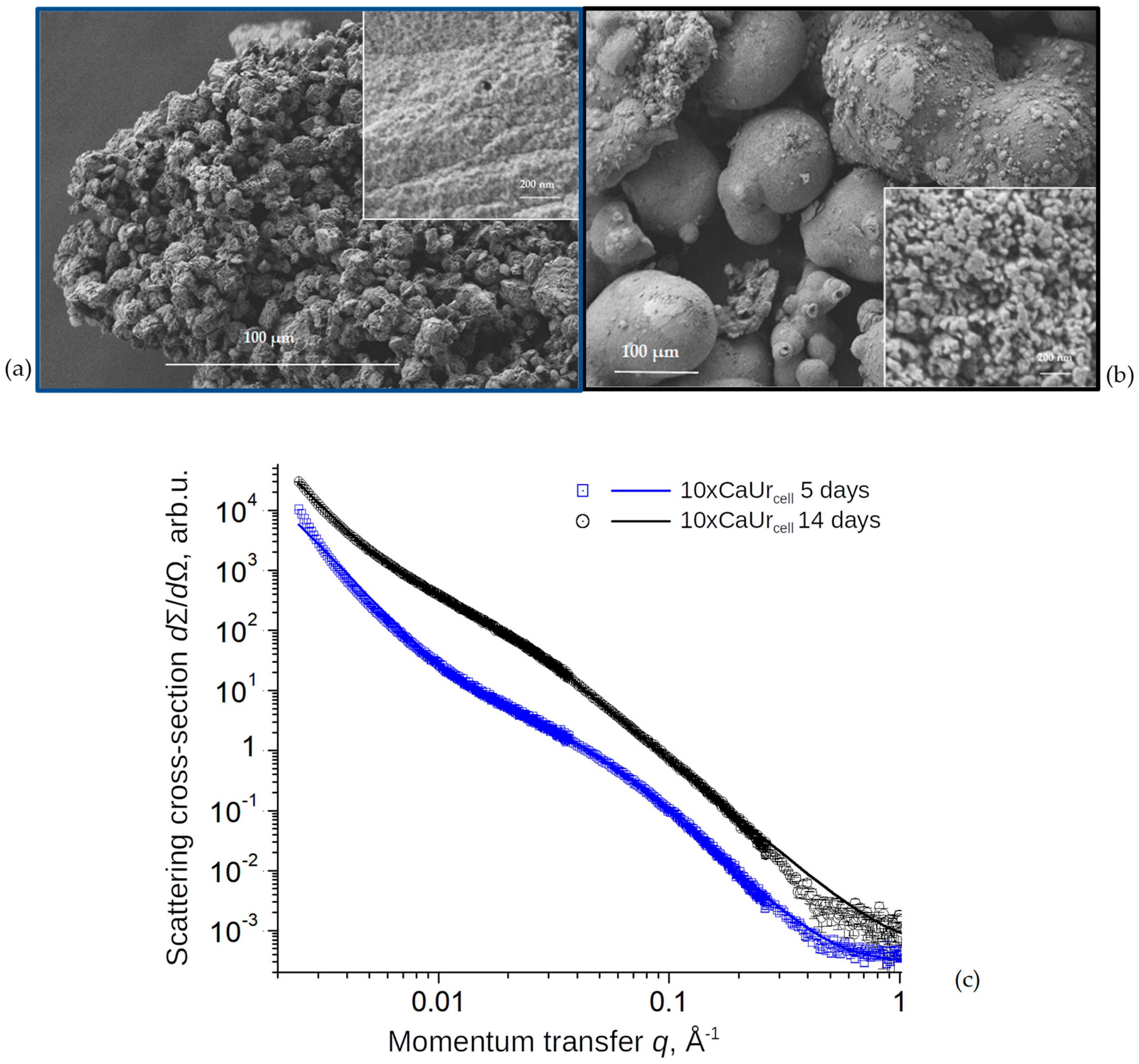
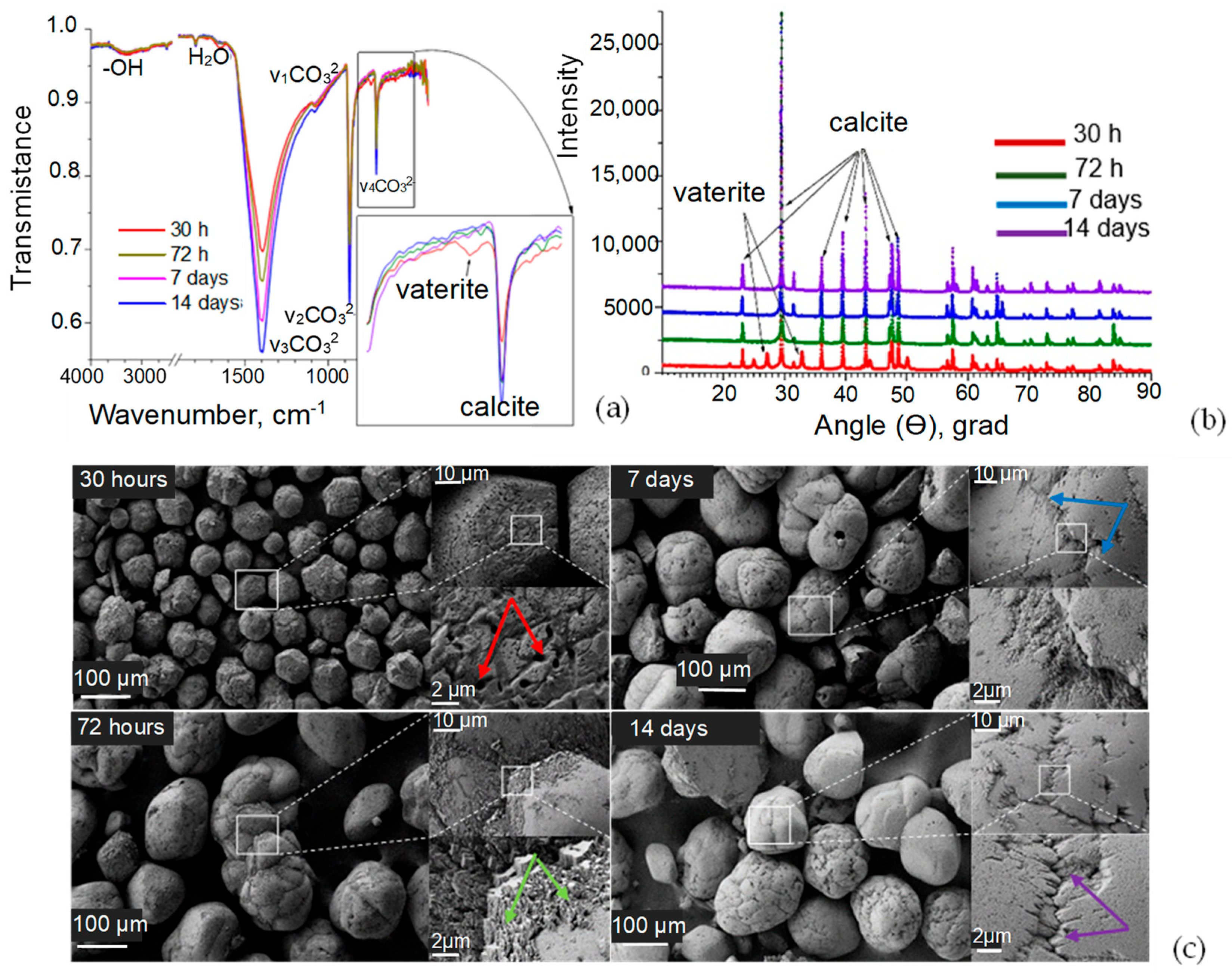
3.2. Structure of Precipitates Formed in Cell System
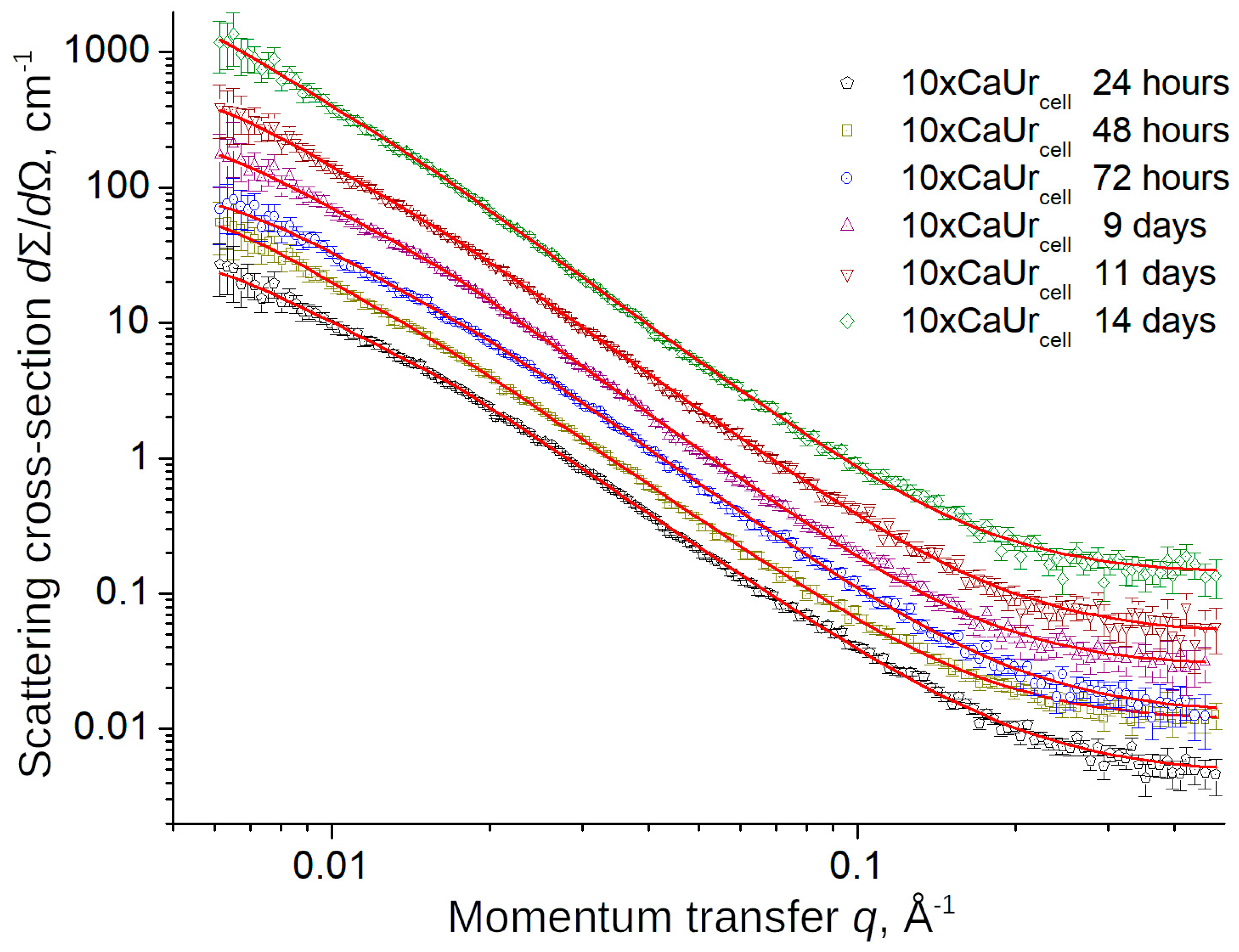
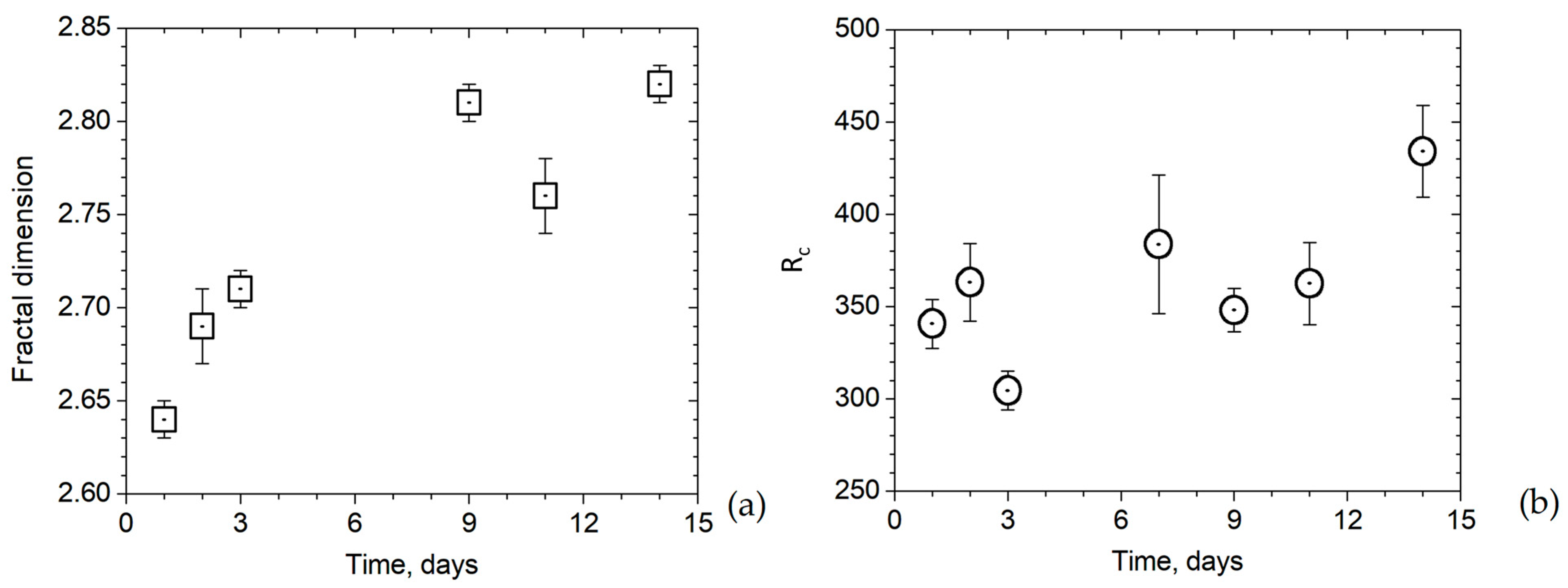
3.3. Evolution of Structure of CaCO3 Precipitates in Cell System
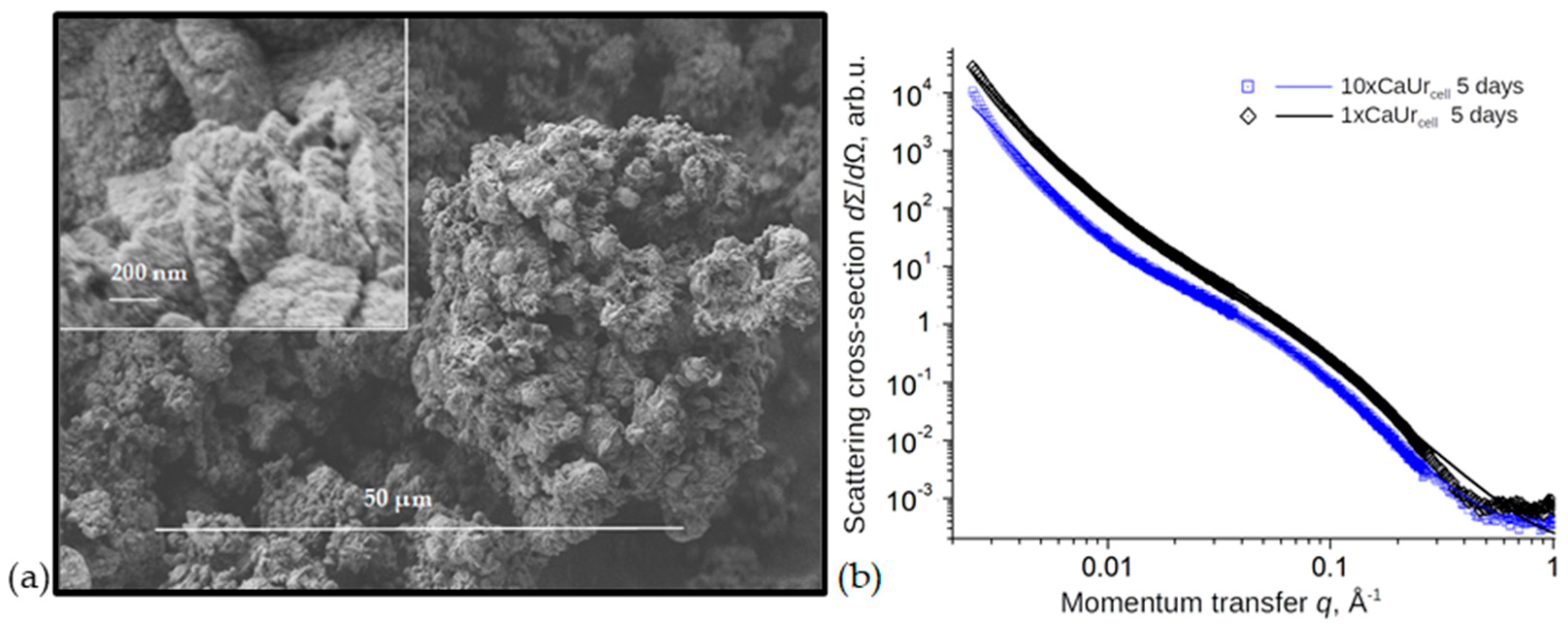
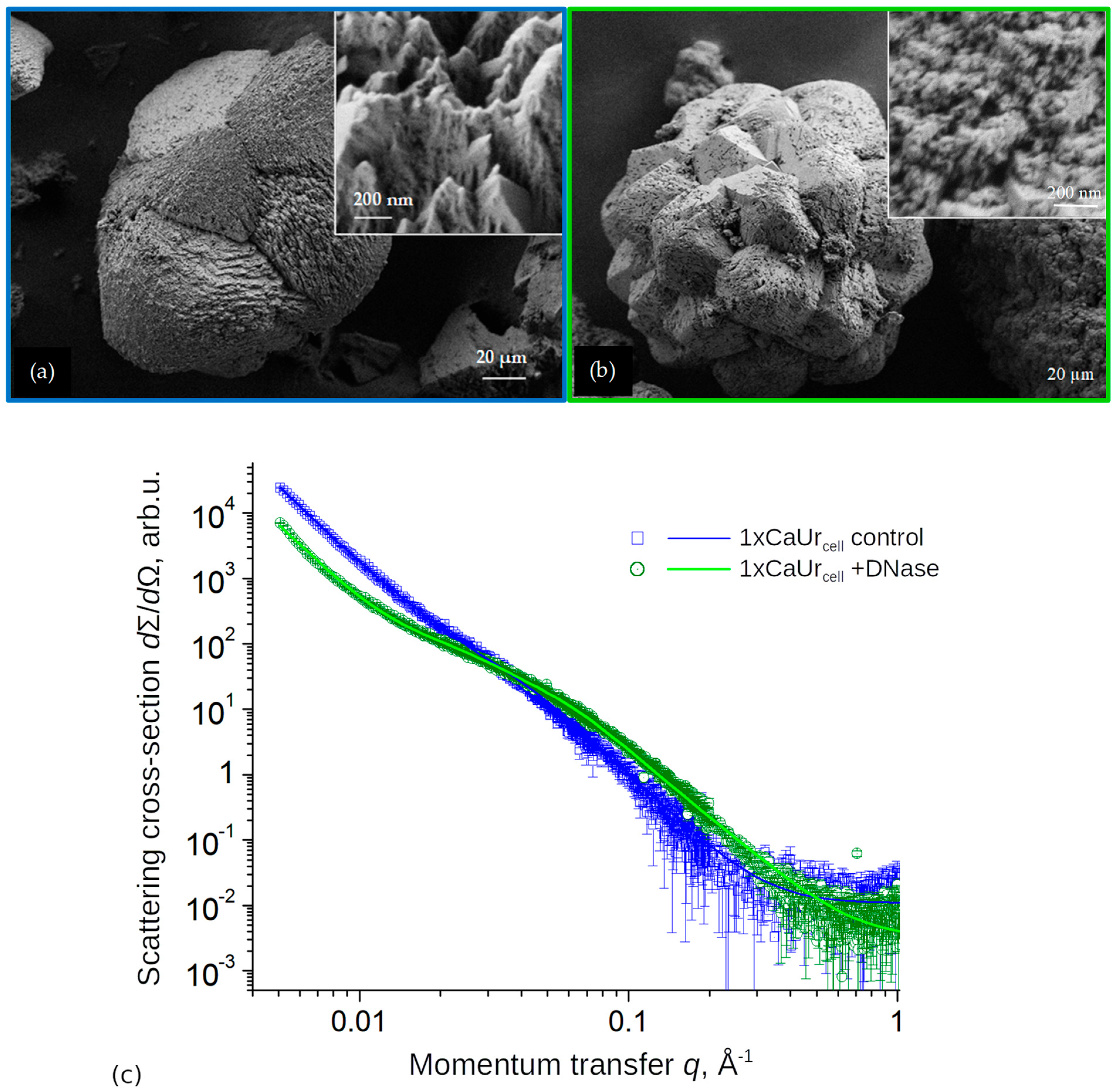
3.4. Effects of DNA Removal during Biomineralization on CaCO3 Structure and Morphology
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A



References
- Seifan, M.; Berenjian, A. Application of Microbially Induced Calcium Carbonate Precipitation in Designing Bio Self-Healing Concrete. World J. Microbiol. Biotechnol. 2018, 34, 168. [Google Scholar] [CrossRef] [PubMed]
- Whiffin, V.S.; van Paassen, L.A.; Harkes, M.P. Microbial Carbonate Precipitation as a Soil Improvement Technique. Geomicrobiol. J. 2007, 24, 417–423. [Google Scholar] [CrossRef]
- Dhami, N.K.; Reddy, M.S.; Mukherjee, A. Biomineralization of Calcium Carbonates and Their Engineered Applications: A Review. Front. Microbiol. 2013, 4, 314. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Park, W. Current Challenges and Future Directions for Bacterial Self-Healing Concrete. Appl. Microbiol. Biotechnol. 2018, 102, 3059–3070. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Goyal, S.; Mukherjee, A.; Reddy, M.S. Microbial Healing of Cracks in Concrete: A Review. J. Ind. Microbiol. Biotechnol. 2017, 44, 1511–1525. [Google Scholar] [CrossRef]
- Ivanov, V.; Chu, J. Applications of Microorganisms to Geotechnical Engineering for Bioclogging and Biocementation of Soil in Situ. Rev. Environ. Sci. Biotechnol. 2008, 7, 139–153. [Google Scholar] [CrossRef]
- Achal, V.; Mukherjee, A.; Kumari, D.; Zhang, Q. Biomineralization for Sustainable Construction—A Review of Processes and Applications. Earth Sci. Rev. 2015, 148, 1–17. [Google Scholar] [CrossRef]
- Yuan, F.; Huang, Z.; Yang, T.; Wang, G.; Li, P.; Yang, B.; Li, J. Pathogenesis of Proteus mirabilis in Catheter-Associated Urinary Tract Infections. Urol. Int. 2021, 105, 354–361. [Google Scholar] [CrossRef]
- Jones, S.M.; Yerly, J.; Hu, Y.; Ceri, H.; Martinuzzi, R. Structure of Proteus Mirabilis Biofilms Grown in Artificial Urine and Standard Laboratory Media. FEMS Microbiol. Lett. 2007, 268, 16–21. [Google Scholar] [CrossRef]
- Tomer, N.; Garden, E.; Small, A.; Palese, M. Ureteral Stent Encrustation: Epidemiology, Pathophysiology, Management and Current Technology. J. Urol. 2021, 205, 68–77. [Google Scholar] [CrossRef]
- Akcalı, A.; Lang, N.P. Dental Calculus: The Calcified Biofilm and Its Role in Disease Development. Periodontology 2000 2018, 76, 109–115. [Google Scholar] [CrossRef]
- Hoffmann, T.D.; Paine, K.; Gebhard, S. Genetic Optimisation of Bacteria-Induced Calcite Precipitation in Bacillus Subtilis. Microb. Cell Fact. 2021, 20, 214. [Google Scholar] [CrossRef]
- Decho, A.W. Overview of Biopolymer-Induced Mineralization: What Goes on in Biofilms? Ecol. Eng. 2010, 36, 137–144. [Google Scholar] [CrossRef]
- Li, X.; Chopp, D.L.; Russin, W.A.; Brannon, P.T.; Parsek, M.R.; Packman, A.I. Spatial Patterns of Carbonate Biomineralization in Biofilms. Appl. Environ. Microbiol. 2015, 81, 7403–7410. [Google Scholar] [CrossRef]
- Al Disi, Z.A.; Zouari, N.; Dittrich, M.; Jaoua, S.; Al-Kuwari, H.A.S.; Bontognali, T.R.R. Characterization of the Extracellular Polymeric Substances (EPS) of Virgibacillus Strains Capable of Mediating the Formation of High Mg-Calcite and Protodolomite. Mar. Chem. 2019, 216, 103693. [Google Scholar] [CrossRef]
- Ivanova, L.A.; Egorov, V.V.; Zabrodskaya, Y.A.; Shaldzhyan, A.A.; Baranchikov, A.Y.; Tsvigun, N.V.; Lykholay, A.N.; Yapryntsev, A.D.; Lebedev, D.V.; Kulminskaya, A.A. Matrix Is Everywhere: Extracellular DNA Is a Link between Biofilm and Mineralization in Bacillus Cereus Planktonic Lifestyle. NPJ Biofilms Microbiomes 2023, 9, 9. [Google Scholar] [CrossRef]
- Zehner, J.; Røyne, A.; Sikorski, P. Calcite Seed-Assisted Microbial Induced Carbonate Precipitation (MICP). PLoS ONE 2021, 16, e0240763. [Google Scholar] [CrossRef]
- Cheng, M.; Sun, S.; Wu, P. Microdynamic Changes of Moisture-Induced Crystallization of Amorphous Calcium Carbonate Revealed via in Situ FTIR Spectroscopy. Phys. Chem. Chem. Phys. 2019, 21, 21882–21889. [Google Scholar] [CrossRef]
- Hammes, F.; Verstraete, W. Key Roles of PH and Calcium Metabolism in Microbial Carbonate Precipitation. Rev. Environ. Sci. Biotechnol. 2002, 1, 3–7. [Google Scholar] [CrossRef]
- Anbu, P.; Kang, C.-H.; Shin, Y.-J.; So, J.-S. Formations of Calcium Carbonate Minerals by Bacteria and Its Multiple Applications. Springerplus 2016, 5, 250. [Google Scholar] [CrossRef]
- Seifan, M.; Samani, A.K.; Berenjian, A. Induced Calcium Carbonate Precipitation Using Bacillus Species. Appl. Microbiol. Biotechnol. 2016, 100, 9895–9906. [Google Scholar] [CrossRef]
- Segovia-Campos, I.; Martignier, A.; Filella, M.; Jaquet, J.; Ariztegui, D. Micropearls and Other Intracellular Inclusions of Amorphous Calcium Carbonate: An Unsuspected Biomineralization Capacity Shared by Diverse Microorganisms. Environ. Microbiol. 2022, 24, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, J.H.E.; Checa, A.G.; Gale, J.D.; Gebauer, D.; Sainz-Díaz, C.I. Calcium Carbonate Polyamorphism and Its Role in Biomineralization: How Many Amorphous Calcium Carbonates Are There? Angew. Chem. Int. Ed. 2012, 51, 11960–11970. [Google Scholar] [CrossRef] [PubMed]
- Cantaert, B.; Kuo, D.; Matsumura, S.; Nishimura, T.; Sakamoto, T.; Kato, T. Use of Amorphous Calcium Carbonate for the Design of New Materials. Chempluschem 2017, 82, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Zhao, Y.; Guan, Q.; Yang, S.; Wang, W.; Yan, B.; Meng, Y.; Li, S.; Tang, P.; Mao, L.; et al. Amorphous Calcium Carbonate Cluster Nanospheres in Water-Deficient Organic Solvents. Angew. Chem. Int. Ed. 2022, 61, e202211254. [Google Scholar] [CrossRef]
- Grünewald, T.A.; Checchia, S.; Dicko, H.; Le Moullac, G.; Sham Koua, M.; Vidal-Dupiol, J.; Duboisset, J.; Nouet, J.; Grauby, O.; Di Michiel, M.; et al. Structure of an Amorphous Calcium Carbonate Phase Involved in the Formation of Pinctada Margaritifera Shells. Proc. Natl. Acad. Sci. USA 2022, 119, e2212616119. [Google Scholar] [CrossRef]
- Fernandez-Martinez, A.; Kalkan, B.; Clark, S.M.; Waychunas, G.A. Pressure-Induced Polyamorphism and Formation of ‘Aragonitic’ Amorphous Calcium Carbonate. Angew. Chem. 2013, 125, 8512–8515. [Google Scholar] [CrossRef]
- Clark, S.M.; Colas, B.; Jacob, D.E.; Neuefeind, J.C.; Wang, H.-W.; Page, K.L.; Soper, A.K.; Schodder, P.I.; Duchstein, P.; Zubiri, B.A.; et al. The Nano- and Meso-Scale Structure of Amorphous Calcium Carbonate. Sci. Rep. 2022, 12, 6870. [Google Scholar] [CrossRef]
- Soper, A.K. Empirical Potential Monte Carlo Simulation of Fluid Structure. Chem. Phys. 1996, 202, 295–306. [Google Scholar] [CrossRef]
- Tobler, D.J.; Rodriguez Blanco, J.D.; Sørensen, H.O.; Stipp, S.L.S.; Dideriksen, K. Effect of PH on Amorphous Calcium Carbonate Structure and Transformation. Cryst. Growth Des. 2016, 16, 4500–4508. [Google Scholar] [CrossRef]
- Yang, G.; Li, F.; Wang, Y.; Ji, C.; Huang, L.; Su, Z.; Li, X.; Zhang, C. The Effect of Bacillus Cereus LV-1 on the Crystallization and Polymorphs of Calcium Carbonate. RSC Adv. 2022, 12, 26908–26921. [Google Scholar] [CrossRef]
- Wang, H.-W.; Daemen, L.L.; Cheshire, M.C.; Kidder, M.K.; Stack, A.G.; Allard, L.F.; Neuefeind, J.; Olds, D.; Liu, J.; Page, K. Synthesis and Structure of Synthetically Pure and Deuterated Amorphous (Basic) Calcium Carbonates. Chem. Commun. 2017, 53, 2942–2945. [Google Scholar] [CrossRef]
- Liu, R.; Huang, S.; Zhang, X.; Song, Y.; He, G.; Wang, Z.; Lian, B. Bio-Mineralisation, Characterization, and Stability of Calcium Carbonate Containing Organic Matter. RSC Adv. 2021, 11, 14415–14425. [Google Scholar] [CrossRef]
- Steciuk, G.; Palatinus, L.; Rohlíček, J.; Ouhenia, S.; Chateigner, D. Stacking Sequence Variations in Vaterite Resolved by Precession Electron Diffraction Tomography Using a Unified Superspace Model. Sci. Rep. 2019, 9, 9156. [Google Scholar] [CrossRef]
- Lyu, J.; Li, F.; Zhang, C.; Gower, L.; Wasman, S.; Sun, J.; Yang, G.; Chen, J.; Gu, L.; Tang, X.; et al. From the inside out: Elemental Compositions and Mineral Phases Provide Insights into Bacterial Calcification. Chem. Geol. 2021, 559, 119974. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, M.; Wang, H.; Tang, D.; Huang, J.; Sun, Y. Study on the Remediation of Cd Pollution by the Biomineralization of Urease-Producing Bacteria. Int. J. Environ. Res. Public Health 2019, 16, 268. [Google Scholar] [CrossRef]
- Khanjani, M.; Westenberg, D.J.; Kumar, A.; Ma, H. Tuning Polymorphs and Morphology of Microbially Induced Calcium Carbonate: Controlling Factors and Underlying Mechanisms. ACS Omega 2021, 6, 11988–12003. [Google Scholar] [CrossRef]
- Wei, S.; Cui, H.; Jiang, Z.; Liu, H.; He, H.; Fang, N. Biomineralization Processes of Calcite Induced by Bacteria Isolated from Marine Sediments. Braz. J. Microbiol. 2015, 46, 455–464. [Google Scholar] [CrossRef]
- Golovkina, D.A.; Zhurishkina, E.V.; Ivanova, L.A.; Baranchikov, A.E.; Sokolov, A.Y.; Bobrov, K.S.; Masharsky, A.E.; Tsvigun, N.V.; Kopitsa, G.P.; Kulminskaya, A.A. Calcifying Bacteria Flexibility in Induction of CaCO3 Mineralization. Life 2020, 10, 317. [Google Scholar] [CrossRef]
- Marvasi, M.; Gallagher, K.L.; Martinez, L.C.; Molina Pagan, W.C.; Rodríguez Santiago, R.E.; Castilloveitía Vega, G.; Visscher, P.T. Importance of B4 Medium in Determining Organomineralization Potential of Bacterial Environmental Isolates. Geomicrobiol. J. 2012, 29, 916–924. [Google Scholar] [CrossRef]
- Klepetsanis, P.G.; Kladi, A.; Ostvold, T.; Kontoyiannis, C.G.; Koutsoukos, P.G.; Amjad, Z.; Reddy, M.M. The Inhibition of Calcium Carbonate Formation in Aqueous Supersaturated Solutions, Spontaneous Precipitation and Seeded Crystal Growth. In Advances in Crystal Growth Inhibition Technologies; Kluwer Academic Publishers: Boston, MA, USA, 2022; pp. 123–137. [Google Scholar]
- Krishnapriya, S.; Venkatesh Babu, D.L.; Prince Arulraj, G. Isolation and Identification of Bacteria to Improve the Strength of Concrete. Microbiol. Res. 2015, 174, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Beaucage, G. Approximations Leading to a Unified Exponential/Power-Law Approach to Small-Angle Scattering. J. Appl. Crystallogr. 1995, 28, 717–728. [Google Scholar] [CrossRef]
- Simon, P.; Pompe, W.; Gruner, D.; Sturm, E.; Ostermann, K.; Matys, S.; Vogel, M.; Rödel, G. Nested Formation of Calcium Carbonate Polymorphs in a Bacterial Surface Membrane with a Graded Nanoconfinement: An Evolutionary Strategy to Ensure Bacterial Survival. ACS Biomater. Sci. Eng. 2022, 8, 526–539. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, K. IK-Spektry i Spektry KR Neorganicheskikh i Koordinatsionnykh Soedinenii [Infrared and Raman Spectra of Inorganic and Coordination Compounds*]; Mir: Moscow, Russia, 1991. [Google Scholar]
- Tammer, M.G. Sokrates: Infrared and Raman Characteristic Group Frequencies: Tables and Charts. Colloid Polym. Sci. 2004, 283, 235. [Google Scholar] [CrossRef]
- Jones, G.C.; Jackson, B. Infrared Transmission Spectra of Carbonate Minerals; Springer: Dordrecht, The Netherlands, 1993; ISBN 978-94-010-4940-5. [Google Scholar]
- Zhong, C.; Chu, C.C. On the Origin of Amorphous Cores in Biomimetic CaCO3 Spherulites: New Insights into Spherulitic Crystallization. Cryst. Growth Des. 2010, 10, 5043–5049. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, C.; Jimenez-Lopez, C.; Rodriguez-Navarro, A.; Gonzalez-Muñoz, M.T.; Rodriguez-Gallego, M. Bacterially Mediated Mineralization of Vaterite. Geochim. Cosmochim. Acta 2007, 71, 1197–1213. [Google Scholar] [CrossRef]
- Beniash, E.; Aizenberg, J.; Addadi, L.; Weiner, S. Amorphous Calcium Carbonate Transforms into Calcite during Sea Urchin Larval Spicule Growth. Proc. R Soc. Lond. B Biol. Sci. 1997, 264, 461–465. [Google Scholar] [CrossRef]
- Jayaraman, A.; Subramanyam, G.; Sindhu, S.; Ajikumar, P.K.; Valiyaveettil, S. Biomimetic Synthesis of Calcium Carbonate Thin Films Using Hydroxylated Poly(Methyl Methacrylate) (PMMA) Template. Cryst. Growth Des. 2007, 7, 142–146. [Google Scholar] [CrossRef]
- Yan, G.W.; Huang, J.H.; Zhang, J.F.; Qian, C.J. Aggregation of Hollow CaCO3 Spheres by Calcite Nanoflakes. Mater. Res. Bull. 2008, 43, 2069–2077. [Google Scholar] [CrossRef]
- Mantilaka, M.M.M.G.P.G.; Pitawala, H.M.T.G.A.; Rajapakse, R.M.G.; Karunaratne, D.G.G.P.; Upul Wijayantha, K.G. Formation of Hollow Bone-like Morphology of Calcium Carbonate on Surfactant/Polymer Templates. J. Cryst. Growth 2014, 392, 52–59. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample * | Experiment | Added CaCl2, g/L | Yield of CaCO3 Precipitates, mg per 100 mL | Observed Precipitation Time, min |
|---|---|---|---|---|
| 10xCaUr | Supernatant with urea (25 g/L) + exDNA | 25 | 17.9 | 5 |
| 10xCaUrcontr | Supernatant with urea (25 g/L) | 25 | 17.6 | 30 |
| 1xCaUr | Supernatant with urea (2.5 g/L) + exDNA | 2.5 | 8.3 | 8 |
| 1xCaUrcontr | Supernatant with urea (2.5 g/L) | 2.5 | 8.2 | 40 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, L.A.; Golovkina, D.A.; Zhurishkina, E.V.; Gorshkova, Y.E.; Yapryntsev, A.D.; Baranchikov, A.E.; Tsvigun, N.V.; Kopitsa, G.P.; Kulminskaya, A.A.; Lebedev, D.V. Structure Evolution of CaCO3 Precipitates Formed during the Bacillus cereus Induced Biomineralization. Minerals 2023, 13, 740. https://0-doi-org.brum.beds.ac.uk/10.3390/min13060740
Ivanova LA, Golovkina DA, Zhurishkina EV, Gorshkova YE, Yapryntsev AD, Baranchikov AE, Tsvigun NV, Kopitsa GP, Kulminskaya AA, Lebedev DV. Structure Evolution of CaCO3 Precipitates Formed during the Bacillus cereus Induced Biomineralization. Minerals. 2023; 13(6):740. https://0-doi-org.brum.beds.ac.uk/10.3390/min13060740
Chicago/Turabian StyleIvanova, Lyubov A., Darya A. Golovkina, Elena V. Zhurishkina, Yulia E. Gorshkova, Alexey D. Yapryntsev, Alexander E. Baranchikov, Natalia V. Tsvigun, Gennady P. Kopitsa, Anna A. Kulminskaya, and Dmitry V. Lebedev. 2023. "Structure Evolution of CaCO3 Precipitates Formed during the Bacillus cereus Induced Biomineralization" Minerals 13, no. 6: 740. https://0-doi-org.brum.beds.ac.uk/10.3390/min13060740