Palmitate but Not Oleate Exerts a Negative Effect on Oxygen Utilization in Myoblasts of Patients with the m.3243A>G Mutation: A Pilot Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Myoblasts
2.2. Metabolic Function Measurements with the Seahorse XF96 Cell Analyzer
2.2.1. Mito Stress Test
The Effect of FFAs on the Mitochondrial Respiration
2.2.2. Fatty Acid Oxidation
2.3. Statistical Analysis
2.4. Ethical Statement
3. Results
3.1. Metabolic Function Measurements with the Seahorse XF96 Cell Analyzer
3.1.1. Mito Stress Test
3.1.2. The Effect of FFAs on Mitochondrial Respiration
Differences between Patients and the Controls
The Effect of Palmitate
The Effect of Oleate
3.1.3. Fatty Acid Oxidation (FAO)
The Difference between Patients and the Controls
The Difference between Palmitate and Oleate
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonnefont, J.P.; Demaugre, F.; Prip-Buus, C.; Saudubray, J.M.; Brivet, M.; Abadi, N.; Thuillier, L. Carnitine palmitoyltransferase deficiencies. Mol. Genet. Metab. 1999, 68, 424–440. [Google Scholar] [CrossRef] [PubMed]
- Henique, C.; Mansouri, A.; Fumey, G.; Lenoir, V.; Girard, J.; Bouillaud, F.; Prip-Buus, C.; Cohen, I. Increased mitochondrial fatty acid oxidation is sufficient to protect skeletal muscle cells from palmitate-induced apoptosis. J. Biol. Chem. 2010, 285, 36818–36827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turpin, S.M.; Lancaster, G.I.; Darby, I.; Febbraio, M.A.; Watt, M.J. Apoptosis in skeletal muscle myotubes is induced by ceramides and is positively related to insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E1341–E1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, T.; Eyre, E.; Rodríguez-Calvo, R.; Palomer, X.; Sánchez, R.M.; Merlos, M.; Laguna, J.C.; Vázquez-Carrera, M. Oleate reverses palmitate-induced insulin resistance and inflammation in skeletal muscle cells. J. Biol. Chem. 2008, 283, 11107–11116. [Google Scholar] [CrossRef] [Green Version]
- Riccardi, G.; Giacco, R.; Rivellese, A. Dietary fat, insulin sensitivity and the metabolic syndrome. Clin. Nutr. 2004, 23, 447–456. [Google Scholar] [CrossRef]
- Jarmuszkiewicz, W.; Woyda-Ploszczyca, A.; Koziel, A.; Majerczak, J.; Zoladz, J.A. Temperature controls oxidative phosphorylation and reactive oxygen species production through uncoupling in rat skeletal muscle mitochondria. Free Radic. Biol. Med. 2015, 83, 12–20. [Google Scholar] [CrossRef]
- Tardo-Dino, P.-E.; Touron, J.; Baugé, S.; Bourdon, S.; Koulmann, N.; Malgoyre, A. The effect of a physiological increase in temperature on mitochondrial fatty acid oxidation in rat myofibers. J. Appl. Physiol. 2019, 127, 312–319. [Google Scholar] [CrossRef]
- Montgomery, M.K.; Turner, N. Mitochondrial dysfunction and insulin resistance: An update. Endocr. Connect. 2015, 4, R1–R15. [Google Scholar] [CrossRef] [Green Version]
- Bruce, C.R.; Risis, S.; Babb, J.R.; Yang, C.; Kowalski, G.M.; Selathurai, A.; Lee-Young, R.S.; Weir, J.M.; Yoshioka, K.; Takuwa, Y. Overexpression of sphingosine kinase 1 prevents ceramide accumulation and ameliorates muscle insulin resistance in high-fat diet–fed mice. Diabetes 2012, 61, 3148–3155. [Google Scholar] [CrossRef] [Green Version]
- Samuel, V.T.; Petersen, K.F.; Shulman, G.I. Lipid-induced insulin resistance: Unravelling the mechanism. Lancet 2010, 375, 2267–2277. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.-S.; Kao, S.-H.; Ho, C.-S.; Wei, Y.-H.; Hung, P.-L.; Hsu, M.-H.; Wu, T.-Y.; Wang, T.-J.; Jian, Y.-R.; Lee, T.-H. Inflexibility of AMPK-mediated metabolic reprogramming in mitochondrial disease. Oncotarget 2017, 8, 73627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motlagh Scholle, L.; Schieffers, H.; Al-Robaiy, S.; Thaele, A.; Dehghani, F.; Lehmann Urban, D.; Zierz, S. The Effect of Resveratrol on Mitochondrial Function in Myoblasts of Patients with the Common m. 3243A> G Mutation. Biomolecules 2020, 10, 1103. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, K.; Yamano, A.; Nagashima, M.; Mori, Y.; Anzai, K.; Akehi, Y.; Nomiyama, R.; Asano, T.; Urae, A.; Ono, J. Mitochondrial gene mutations in the tRNALeu (UUR) region and diabetes: Prevalence and clinical phenotypes in Japan. Clin. Chem. 2001, 47, 1641–1648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, M.; Angelini, C.; Bignami, F.; Bodin, A.-M.; Crimi, M.; Di Donato, J.-H.; Felice, A.; Jaeger, C.; Karcagi, V.; LeCam, Y. The EuroBioBank Network: 10 years of hands-on experience of collaborative, transnational biobanking for rare diseases. Eur. J. Hum. Genet. 2015, 23, 1116–1123. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.M.; Caputo, C.; Medici, F.; Pambakian, A.L.; Dornhorst, A.; Meeran, K.; Williams, G.R.; Khoo, B. MELAS syndrome, diabetes and thyroid disease: The role of mitochondrial oxidative stress. Clin. Endocrinol. 2009, 70, 340–341. [Google Scholar] [CrossRef] [PubMed]
- Janssen, G.M.; Hensbergen, P.J.; van Bussel, F.J.; Balog, C.I.; Maassen, J.A.; Deelder, A.M.; Raap, A.K. The A3243G tRNALeu (UUR) mutation induces mitochondrial dysfunction and variable disease expression without dominant negative acting translational defects in complex IV subunits at UUR codons. Hum. Mol. Genet. 2007, 16, 2472–2481. [Google Scholar] [CrossRef] [Green Version]
- Maassen, J.A.; M‘t Hart, L.; van Essen, E.; Heine, R.J.; Nijpels, G.; Tafrechi, R.S.J.; Raap, A.K.; Janssen, G.M.; Lemkes, H.H. Mitochondrial diabetes: Molecular mechanisms and clinical presentation. Diabetes 2004, 53, S103–S109. [Google Scholar] [CrossRef] [Green Version]
- Boden, G. Effects of free fatty acids (FFA) on glucose metabolism: Significance for insulin resistance and type 2 diabetes. Exp. Clin. Endocrinol. Diabetes 2003, 111, 121–124. [Google Scholar] [CrossRef]
- Lim, J.-H.; Gerhart-Hines, Z.; Dominy, J.E.; Lee, Y.; Kim, S.; Tabata, M.; Xiang, Y.K.; Puigserver, P. Oleic acid stimulates complete oxidation of fatty acids through protein kinase A-dependent activation of SIRT1-PGC1α complex. J. Biol. Chem. 2013, 288, 7117–7126. [Google Scholar] [CrossRef] [Green Version]
- Salvado, L.; Coll, T.; Gomez-Foix, A.; Salmeron, E.; Barroso, E.; Palomer, X.; Vazquez-Carrera, M. Oleate prevents saturated-fatty-acid-induced ER stress, inflammation and insulin resistance in skeletal muscle cells through an AMPK-dependent mechanism. Diabetologia 2013, 56, 1372–1382. [Google Scholar] [CrossRef]
- Oliveira, A.F.; Cunha, D.A.; Ladriere, L.; Igoillo-Esteve, M.; Bugliani, M.; Marchetti, P.; Cnop, M. In vitro use of free fatty acids bound to albumin: A comparison of protocols. Biotechniques 2015, 58, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsabeeh, N.; Chausse, B.; Kakimoto, P.A.; Kowaltowski, A.J.; Shirihai, O. Cell culture models of fatty acid overload: Problems and solutions. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2018, 1863, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Theunissen, T.E.J.; Gerards, M.; Hellebrekers, D.; van Tienen, F.H.; Kamps, R.; Sallevelt, S.; Hartog, E.; Scholte, H.R.; Verdijk, R.M.; Schoonderwoerd, K.; et al. Selection and characterization of palmitic acid responsive patients with an oxphos complex I defect. Front. Mol. Neurosci. 2017, 10, 336. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Keung, W.; Samokhvalov, V.; Wang, W.; Lopaschuk, G.D. Role of fatty acid uptake and fatty acid β-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2010, 1801, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Brookheart, R.T.; Michel, C.I.; Schaffer, J.E. As a matter of fat. Cell Metab. 2009, 10, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Gaster, M.; Rustan, A.C.; Beck-Nielsen, H. Differential utilization of saturated palmitate and unsaturated oleate: Evidence from cultured myotubes. Diabetes 2005, 54, 648–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montell, E.; Turini, M.; Marotta, M.; Roberts, M.; Noé, V.; Ciudad, C.J.; Macé, K.; Gómez-Foix, A.M. DAG accumulation from saturated fatty acids desensitizes insulin stimulation of glucose uptake in muscle cells. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E229–E237. [Google Scholar] [CrossRef] [Green Version]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef] [Green Version]
- Yuzefovych, L.; Wilson, G.; Rachek, L. Different effects of oleate vs. palmitate on mitochondrial function, apoptosis, and insulin signaling in L6 skeletal muscle cells: Role of oxidative stress. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E1096–E1105. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Gender | Age at Biopsy | |
|---|---|---|
| Patients | ||
| P 1 | M | 43 |
| P 2 | M | 42 |
| P 3 | F | 70 |
| P 4 | M | 34 |
| P 5 | F | 40 |
| Controls | ||
| C 1 | F | 50 |
| C 2 | M | 53 |
| C 3 | F | 40 |
| C 4 | M | 35 |
| C 5 | F | 49 |
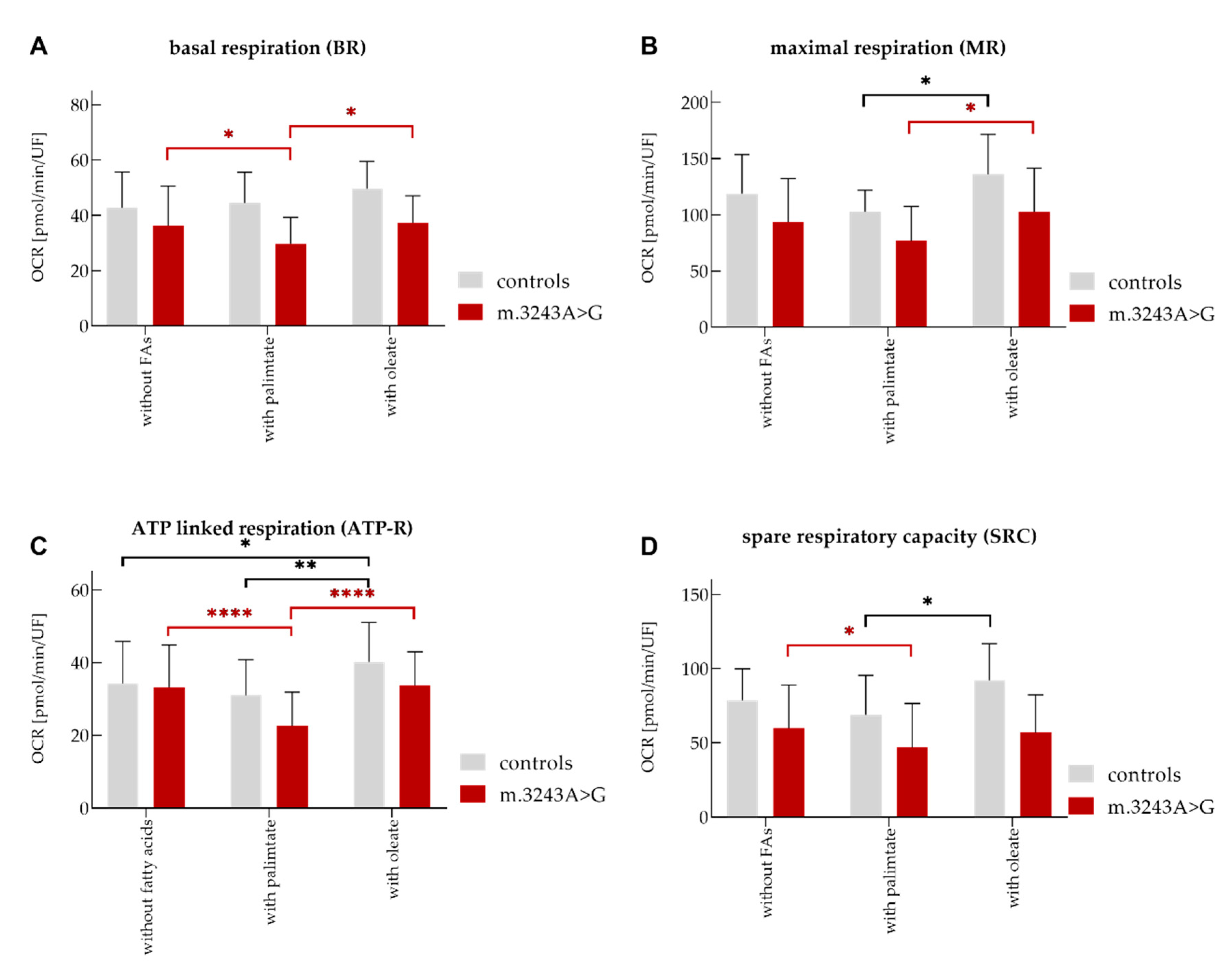
| Mito Stress Test | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| +Palmitate | +Oleate | ||||||||
| Controls (Mean) | Patients (Mean) | p-Value | Controls (Mean) | Patients (Mean) | p-Value | Controls (Mean) | Patients (Mean) | p-Value | |
| Basal | 42.7 | 36.3 | 0.004 | 44.5 | 29.7 | <0.0001 | 49.6 | 37.3 | 0.0006 |
| MR | 118.9 | 93.6 | <0.0001 | 102.6 | 76.7 | 136 | 102.7 | 0.002 | |
| SRC | 78.6 | 59.9 | 0.0003 | 68.8 | 46.8 | 0.006 | 92.3 | 56.9 | <0.0001 |
| ATP | 34.2 | 33.2 | 30.9 | 22.6 | 0.009 | 40.2 | 33.7 | 0.04 | |
| Controls | ||||||||
| –FAs (Mean) | Pa (Mean) | p. –FAs/Pa (Mean) | O (Mean) | p. –FAs/O (Mean) | Pa (Mean) | O (Mean) | p. Pa/O (Mean) | |
| Basal | 42.7 | 44.5 | 49.6 | 44.5 | 49.6 | |||
| MR | 118.9 | 102.6 | 136 | 102.6 | 136 | 0.01 | ||
| SRC | 78.6 | 68.9 | 92.3 | 68.9 | 92.3 | 0.01 | ||
| ATP | 34.2 | 30.9 | 40.2 | 0.02 | 30.9 | 40.2 | 0.003 | |
| Patients | ||||||||
| –FAs (Mean) | Pa (Mean) | p. –FAs/Pa (Mean) | O (Mean) | p. –FAs/O (Mean) | Pa (Mean) | O (Mean) | p. Pa/O (Mean) | |
| Basal | 36.3 | 29.7 | 0.03 | 37.3 | 29.7 | 37.3 | 0.04 | |
| MR | 93.6 | 76.7 | 102.7 | 76.7 | 102.7 | 0.02 | ||
| SRC | 59.9 | 46.8 | 0.04 | 56.9 | 46.8 | 56.9 | ||
| ATP | 33.2 | 22.6 | <0.0001 | 33.7 | 22.6 | 33.7 | <0.0001 | |
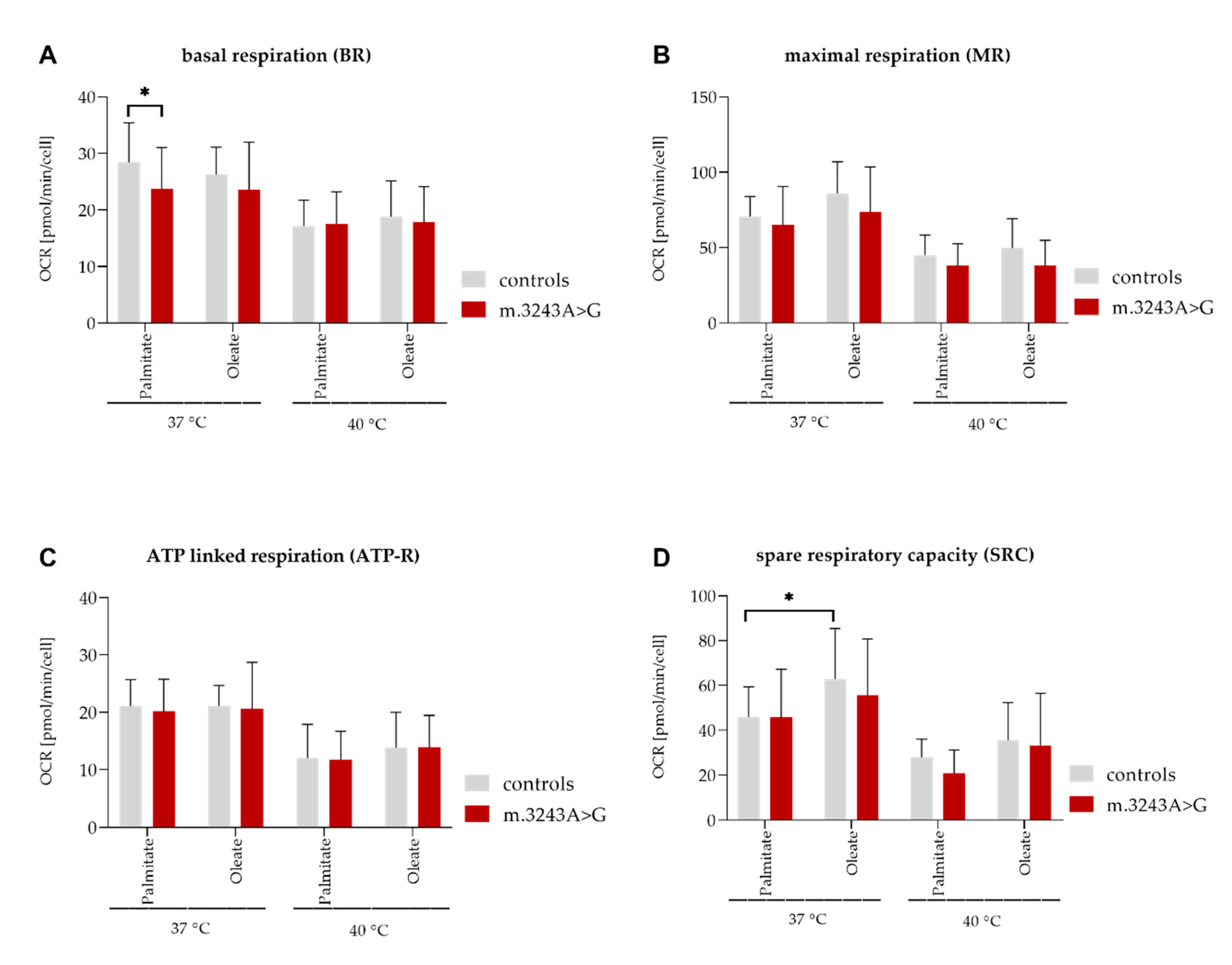
| FAO 37 °C | ||||||
| Palmitate as Substrate | Oleate as Substrate | |||||
| Controls (Mean) | Patients (Mean) | p-Value | Controls (Mean) | Patients (Mean) | p-Value | |
| Basal | 28.4 | 23.7 | 0.04 | 26.3 | 23.5 | |
| MR | 70.7 | 64.9 | 85.9 | 73.7 | ||
| SRC | 46 | 45.7 | 62.8 | 55.5 | ||
| ATP | 21.1 | 20.2 | 21.1 | 20.6 | ||
| FAO 40 °C | ||||||
| Palmitate as Substrate | Oleate as Substrate | |||||
| Controls (Mean) | Patients (Mean) | p-Value | Controls (Mean) | Patients (Mean) | p-Value | |
| Basal | 17.2 | 17.5 | 18.8 | 17.9 | ||
| MR | 45.1 | 38 | 49.5 | 44.3 | ||
| SRC | 28 | 20.8 | 35.5 | 33.3 | ||
| ATP | 12 | 11.7 | 13.9 | 13.9 | ||
| Controls | ||||||
| Palmitate as Substrate | Oleate as Substrate | |||||
| 37 °C (Mean) | 40 °C (Mean) | p-Value | 37 °C (Mean) | 40 °C (Mean) | p-Value | |
| Basal | 28.4 | 17.2 | <0.0001 | 26.3 | 18.8 | 0.005 |
| MR | 70.7 | 45.1 | 0.006 | 85.9 | 49.5 | <0.0001 |
| SRC | 46 | 28 | 0.05 | 62.8 | 35.5 | 0.0002 |
| ATP | 21.1 | 12 | <0.0001 | 21.1 | 13.9 | 0.001 |
| Patients | ||||||
| Palmitate as Substrate | Oleate as Substrate | |||||
| 37 °C (Mean) | 40 °C (Mean) | p-Value | 37 °C (Mean) | 40 °C (Mean) | p-Value | |
| Basal | 23.7 | 17.5 | 0.02 | 23.5 | 17.9 | 0.04 |
| MR | 64.9 | 38 | 0.0006 | 73.7 | 44.3 | 0.0003 |
| SRC | 45.7 | 20.8 | 0.0006 | 55.5 | 33.3 | 0.003 |
| ATP | 20.2 | 11.7 | <0.0001 | 20.6 | 13.9 | 0.004 |
| FAO 37 °C | ||||||
| Controls | Patients | |||||
| Pa (Mean) | O (Mean) | p-Value | Pa (Mean) | O (Mean) | p-Value | |
| Basal | 28.4 | 26.3 | 23.7 | 23.5 | ||
| MR | 70.7 | 85.9 | 64.9 | 73.7 | ||
| SRC | 46 | 62.8 | 0.04 | 45.7 | 55.5 | |
| ATP | 21.1 | 21.1 | 20.2 | 20.6 | ||
| FAO 40 °C | ||||||
| Controls | Patients | |||||
| Pa (Mean) | O (Mean) | p-Value | Pa (Mean) | O (Mean) | p-Value | |
| Basal | 17.2 | 18.8 | 17.5 | 17.9 | ||
| MR | 45.1 | 49.5 | 38 | 44.3 | ||
| SRC | 28 | 35.5 | 20.8 | 33.3 | ||
| ATP | 12 | 13.9 | 11.7 | 13.9 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Motlagh Scholle, L.; Schieffers, H.; Al-Robaiy, S.; Thaele, A.; Lehmann Urban, D.; Zierz, S. Palmitate but Not Oleate Exerts a Negative Effect on Oxygen Utilization in Myoblasts of Patients with the m.3243A>G Mutation: A Pilot Study. Life 2020, 10, 204. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090204
Motlagh Scholle L, Schieffers H, Al-Robaiy S, Thaele A, Lehmann Urban D, Zierz S. Palmitate but Not Oleate Exerts a Negative Effect on Oxygen Utilization in Myoblasts of Patients with the m.3243A>G Mutation: A Pilot Study. Life. 2020; 10(9):204. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090204
Chicago/Turabian StyleMotlagh Scholle, Leila, Helena Schieffers, Samiya Al-Robaiy, Annemarie Thaele, Diana Lehmann Urban, and Stephan Zierz. 2020. "Palmitate but Not Oleate Exerts a Negative Effect on Oxygen Utilization in Myoblasts of Patients with the m.3243A>G Mutation: A Pilot Study" Life 10, no. 9: 204. https://0-doi-org.brum.beds.ac.uk/10.3390/life10090204