
Growth and Antifungal Resistance of the Pathogenic Yeast, Candida Albicans, in the Microgravity Environment of the International Space Station: An Aggregate of Multiple Flight Experiences


 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strain and Growth Conditions
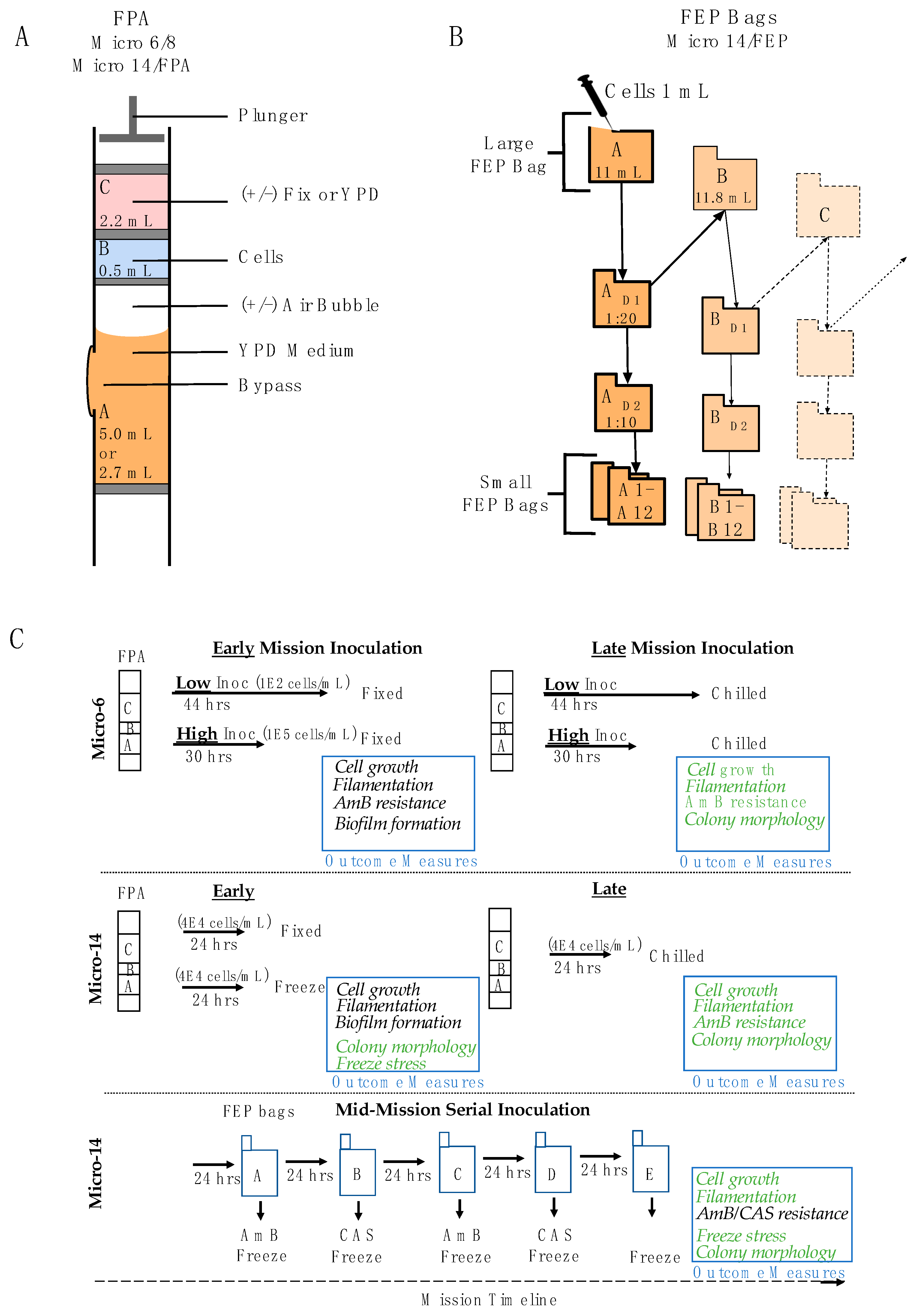
2.2. Missions: Common Procedures
2.3. Mission-Specific Growth Conditions and Procedures
2.3.1. Micro-6 (and the Reflight Micro-8), Space X CRS-1 and -4, Respectively
2.3.2. Micro-14/FPA, Space X CRS-16
2.3.3. Micro-14/FEP, Space X CRS-17
2.4. Ground-Based Analyses
2.4.1. Cell Growth and Morphology
2.4.2. Biofilm formation
2.4.3. Antimicrobial Resistance
2.4.4. Statistical Analysis
3. Results
3.1. Mission Timelines
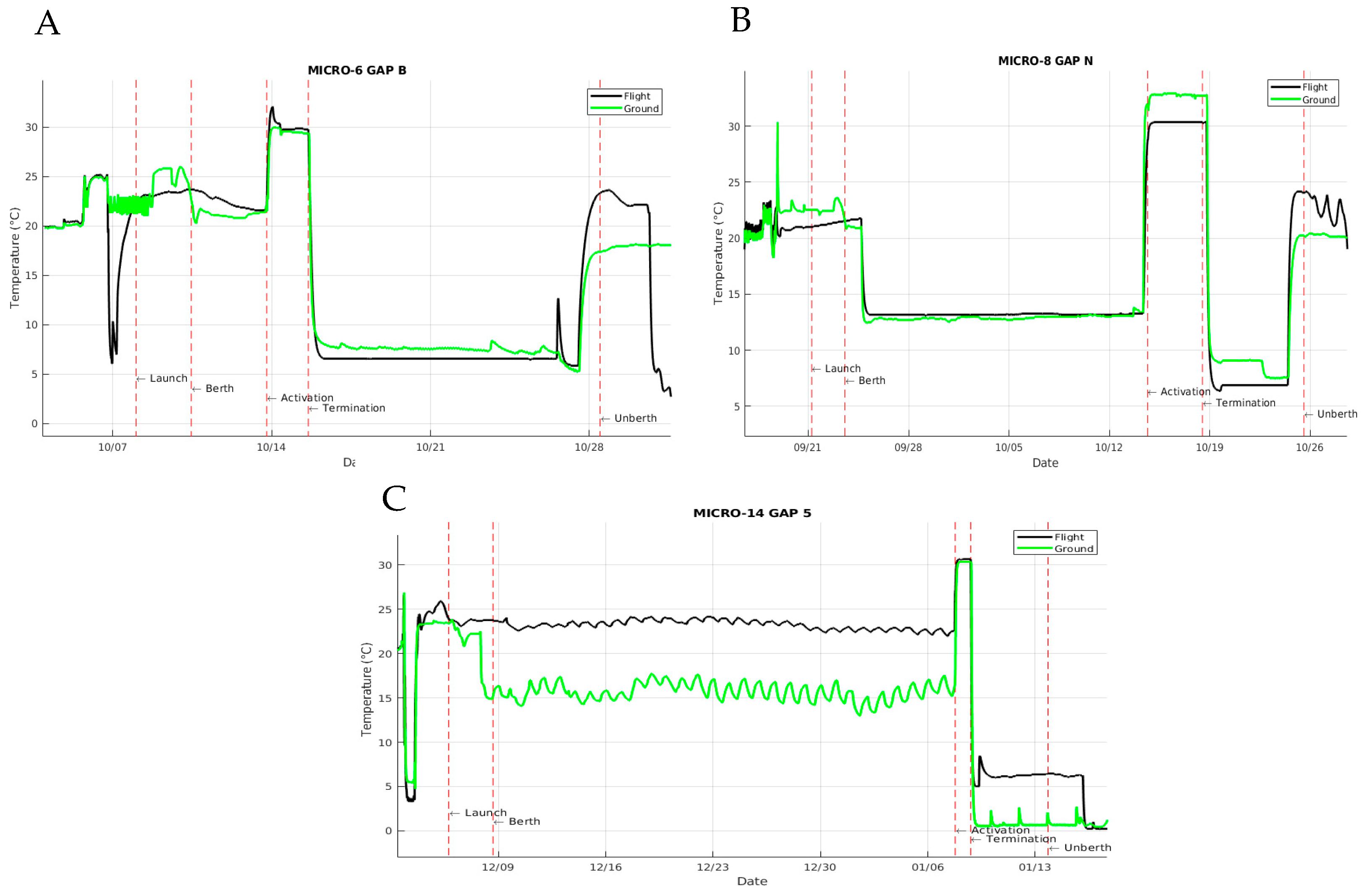
3.2. Sample Temperature Transitions throughout Mission
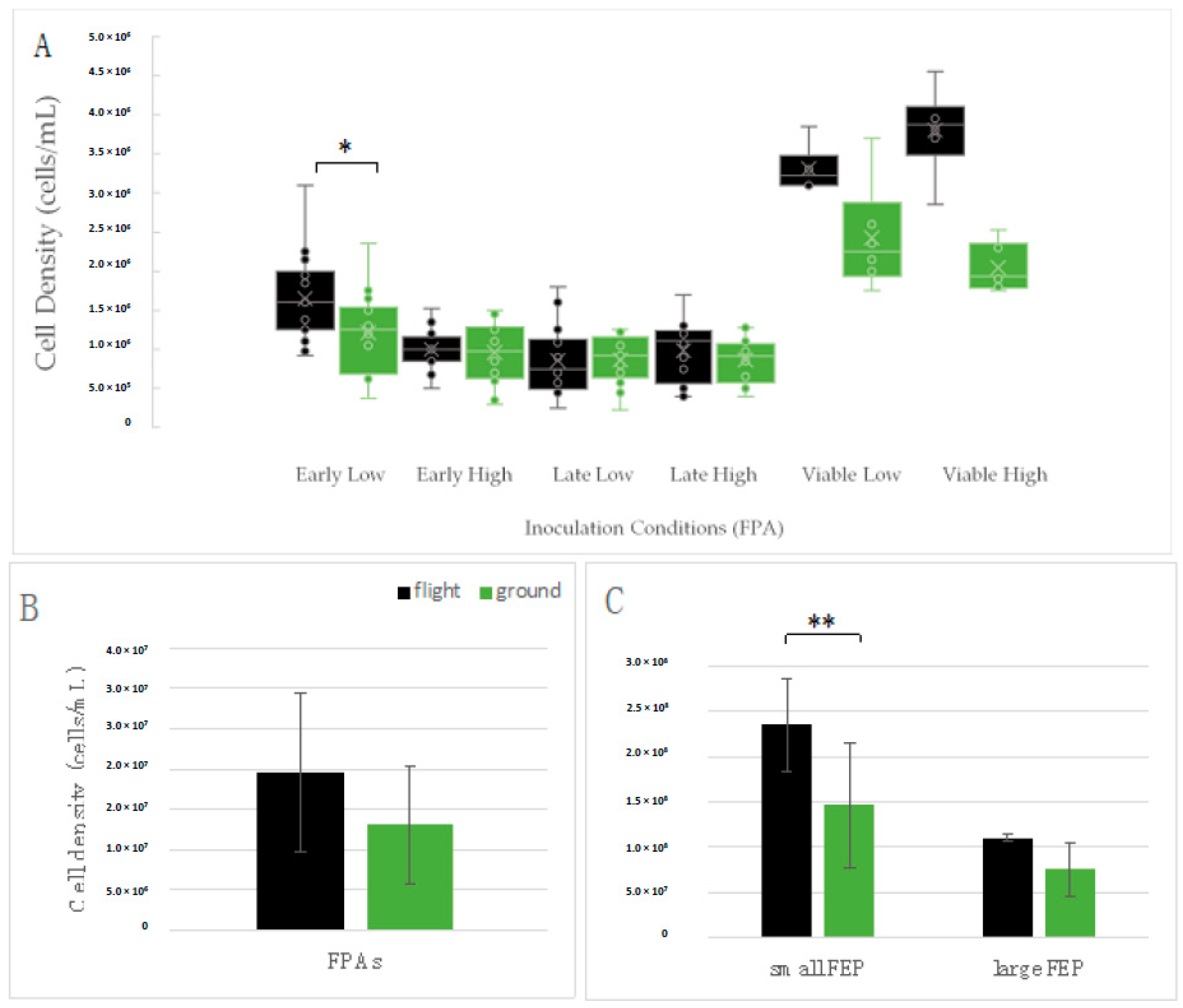
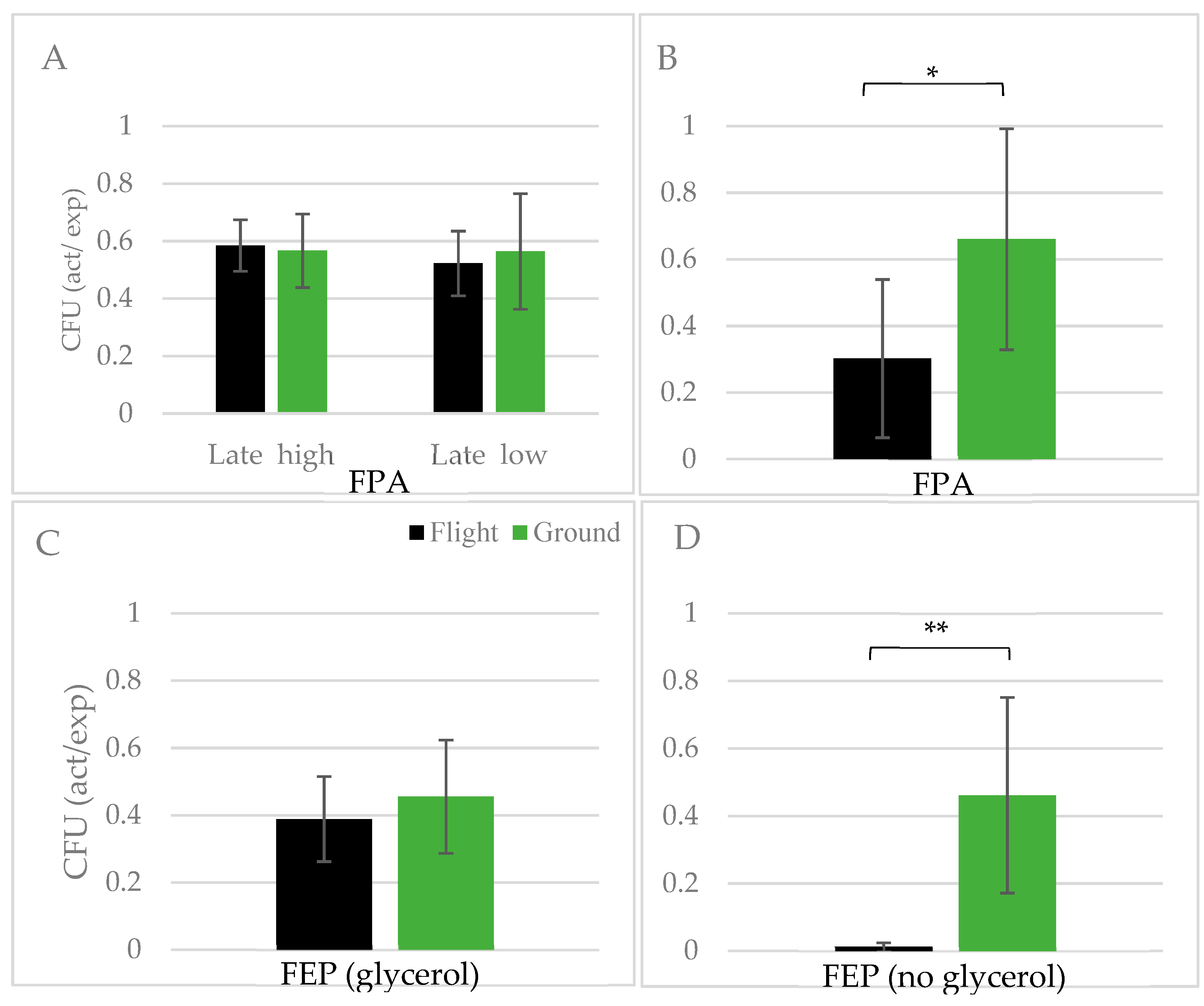
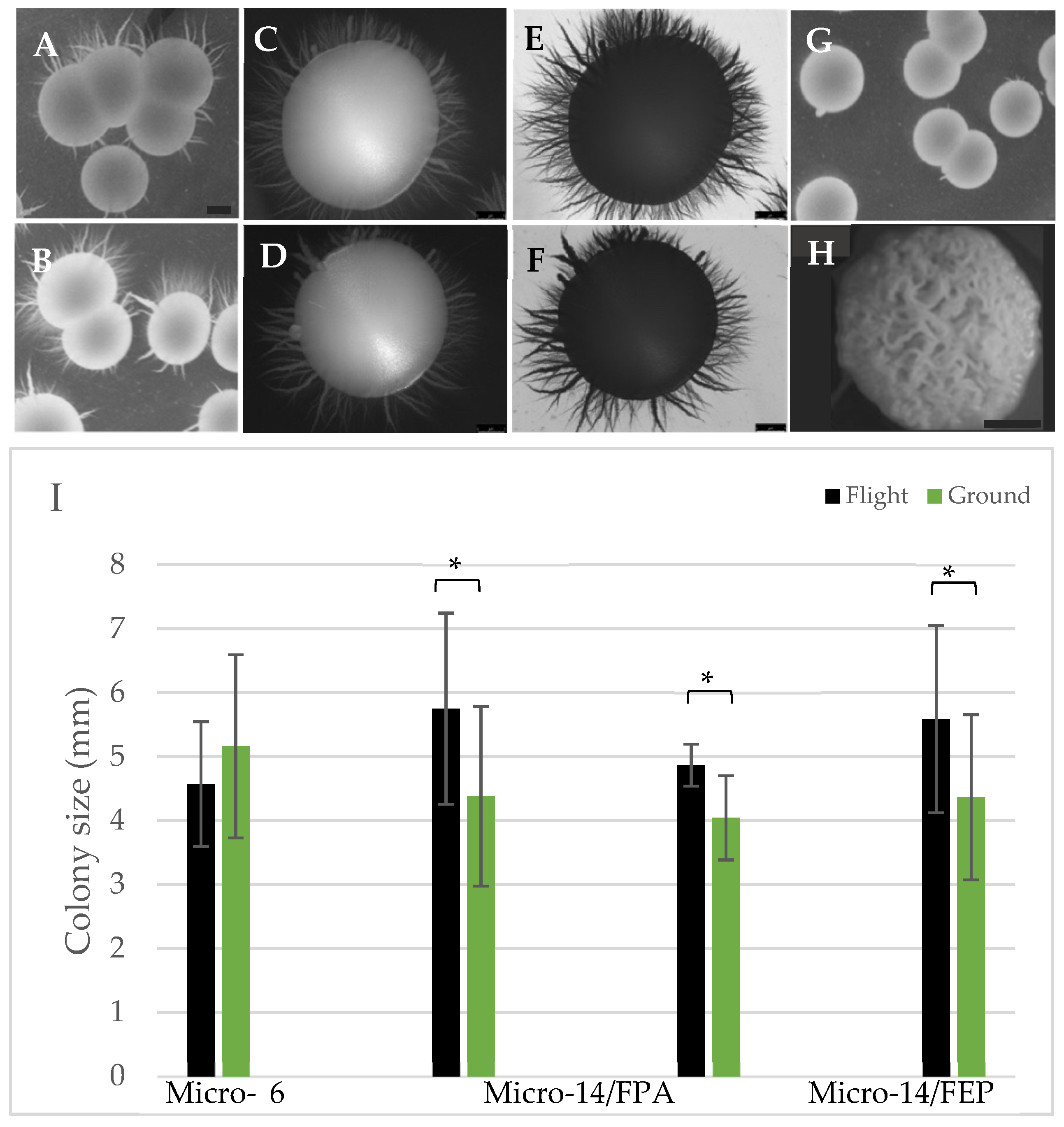
3.3. Yeast Growth
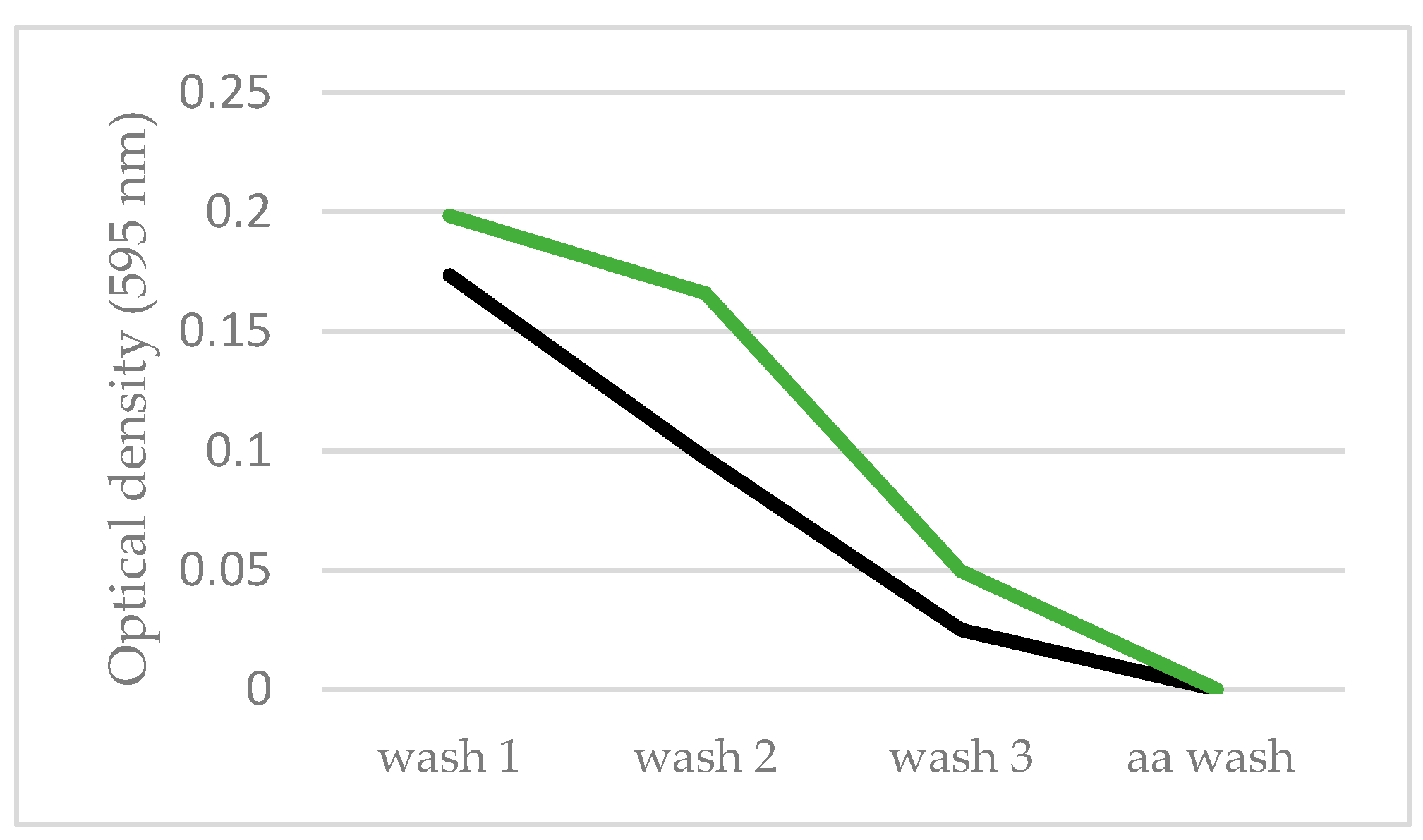
3.4. Biofilm Formation
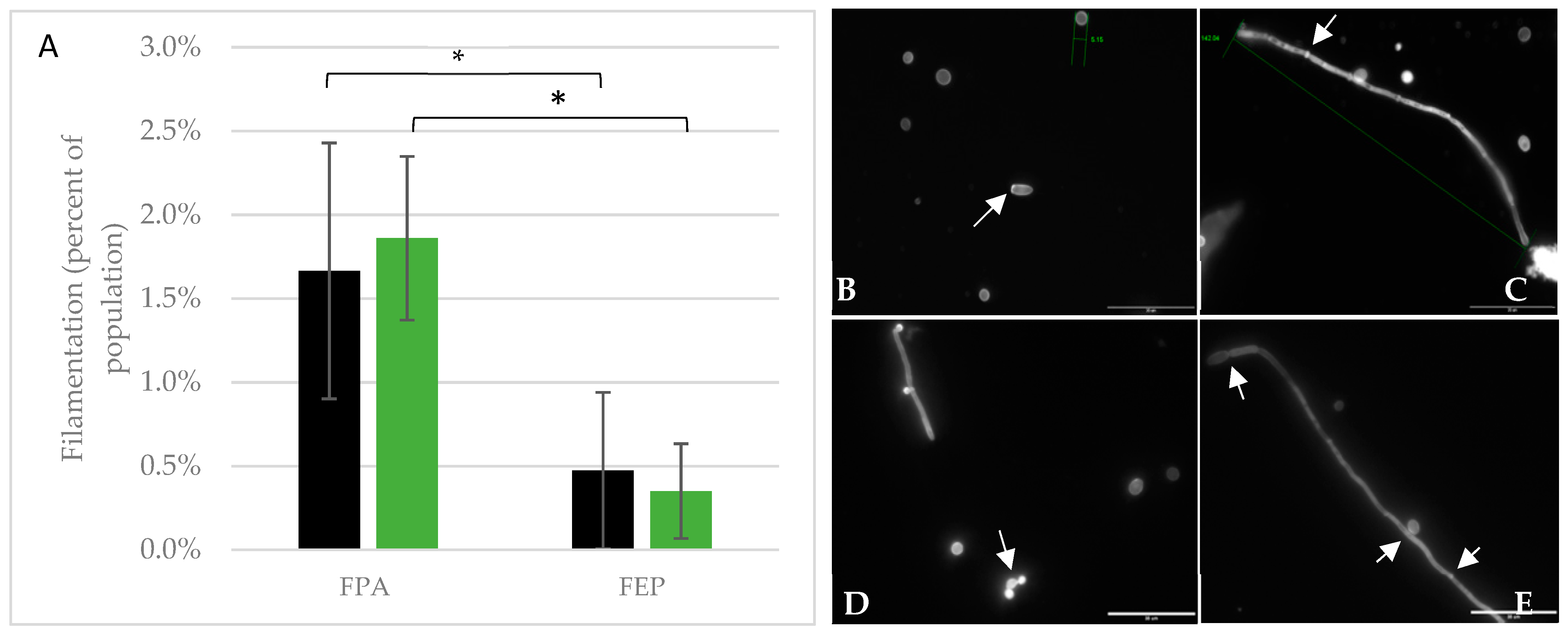
3.5. Cell Morphology and Filamentation
3.6. Characteristics of C. albicans Cultured Post Flight
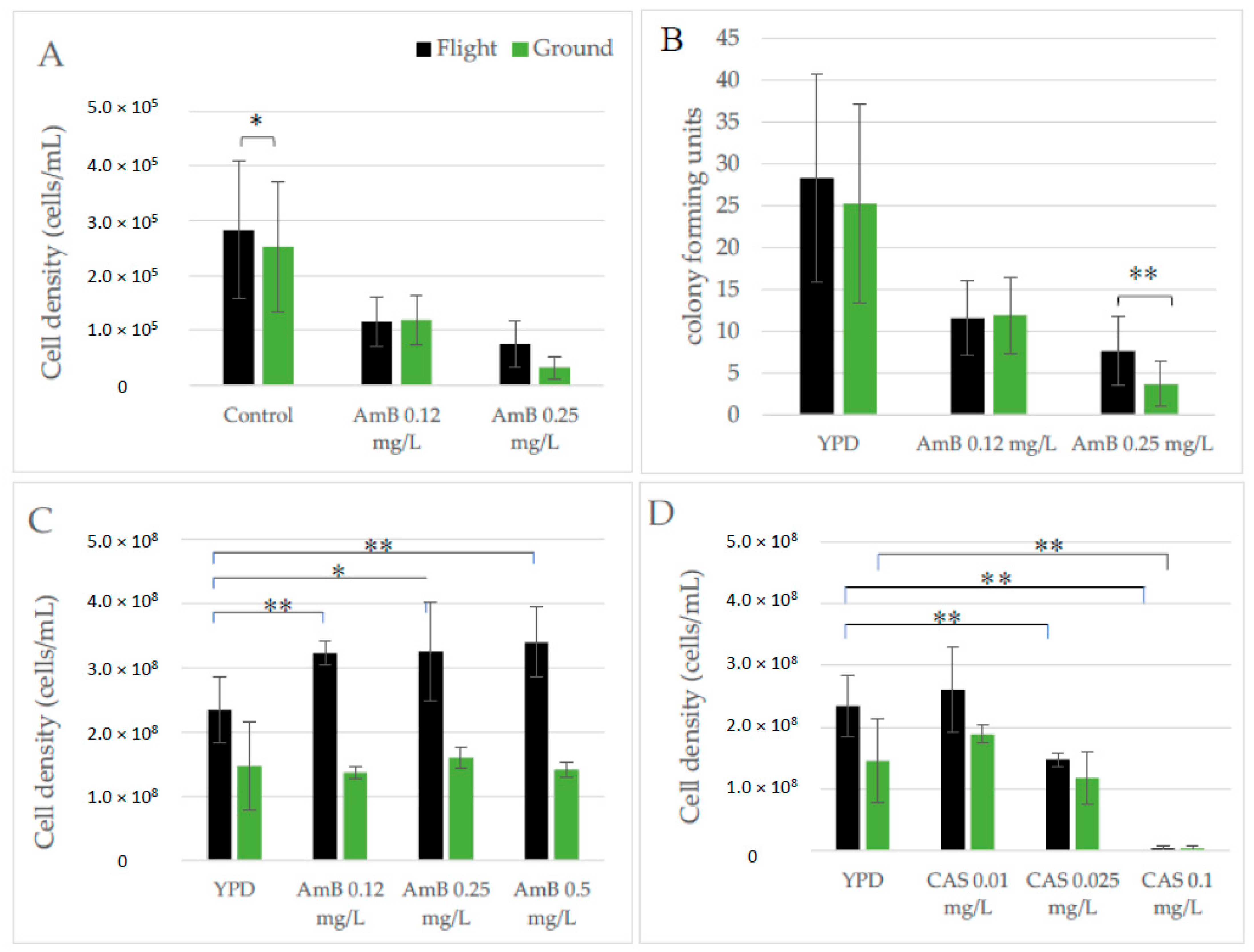
3.7. Susceptibility to Antifungal Agents
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romo, J.A.; Kumamoto, C.A. On Commensalism of Candida. J. Fungi 2020, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neville, B.A.; d’Enfert, C.; Bougnoux, M.E. Candida albicans commensalism in the gastrointestinal tract. FEMS Yeast Res. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Odds, F.C. CandidaInfections: An Overview. CRC Crit. Rev. Microbiol. 1987, 15, 1–5. [Google Scholar] [CrossRef]
- Nobile, C.J.; Johnson, A.D. Candida albicansBiofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of Invasive Candidiasis: A Persistent Public Health Problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [Green Version]
- Crucian, B.E.; Chouker, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune System Dysregulation During Spaceflight: Potential Countermeasures for Deep Space Exploration Missions. Front Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
- Wächtler, B.; Citiulo, F.; Jablonowski, N.; Förster, S.; Dalle, F.; Schaller, M.; Wilson, D.; Hube, B. Candida albicans-Epithelial Interactions: Dissecting the Roles of Active Penetration, Induced Endocytosis and Host Factors on the Infection Process. PLoS ONE 2012, 7, e36952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soll, D.R. Candida commensalism and virulence: The evolution of phenotypic plasticity. ACTA Trop. 2002, 81, 101–110. [Google Scholar] [CrossRef]
- Alonso-Valle, H.; Acha, O.; García-Palomo, J.D.; Fariñas-Álvarez, C.; Fernández-Mazarrasa, C.; Fariñas, M.C. Candidemia in a Tertiary Care Hospital: Epidemiology and Factors Influencing Mortality. Eur. J. Clin. Microbiol. Infect. Dis. 2003, 22, 254–257. [Google Scholar] [CrossRef]
- Pappas, P.G.; Rex, J.H.; Lee, J.; Hamill, R.J.; Larsen, R.A.; Powderly, W.; Kauffman, C.A.; Hyslop, N.; Mangino, J.E.; Chapman, S.; et al. A Prospective Observational Study of Candidemia: Epidemiology, Therapy, and Influences on Mortality in Hospitalized Adult and Pediatric Patients. Clin. Infect. Dis. 2003, 37, 634–643. [Google Scholar] [CrossRef] [Green Version]
- Costa-de-Oliveira, S.; Rodrigues, A.G. Candida albicans antifungal resistance and tolerance in bloodstream infections: The triad yeast-host-antifungal. Microorganisms 2020, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden Killers: Human Fungal Infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [Green Version]
- Santos, G.C.D.O.; Vasconcelos, C.C.; Lopes, A.J.O.; Cartágenes, M.D.S.D.S.; Filho, A.K.D.B.; Nascimento, F.R.F.D.; Ramos, R.M.; Pires, E.R.R.B.; De Andrade, M.S.; Rocha, F.M.G.; et al. Candida Infections and Therapeutic Strategies: Mechanisms of Action for Traditional and Alternative Agents. Front Microbiol. 2018, 9, 1351. [Google Scholar] [CrossRef]
- Mesa-Arango, A.C.; Scorzoni, L.; Zaragoza, O. It only takes one to do many jobs: Amphotericin B as antifungal and immunomodulatory drug. Front. Microbiol. 2012, 3, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, A.; Holz, R. Aqueous pores created in thin lipid membranes by the polyene antibiotics nystatin and amphotericin B. Membranes 1973, 2, 377–408. [Google Scholar] [PubMed]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Executive Summary: Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2016, 62, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Ernst, J.F. Transcription factors in Candida albicans-environmental control of morphogenesis. Microbiology 2000, 146, 1763–1774. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, R.; Cowen, L. Coupling temperature sensing and development: Hsp90 regulates morphogenetic signalling in Candida albicans. Virulence 2010, 1, 45–48. [Google Scholar] [CrossRef] [Green Version]
- Kullas, A.L.; Martin, S.J.; Davis, D. Adaptation to environmental pH: Integrating the Rim101 and calcineurin signal transduction pathways. Mol. Microbiol. 2007, 66, 858–871. [Google Scholar] [CrossRef]
- Lu, Y.; Su, C.; Solis, N.V.; Filler, S.G.; Liu, H. Synergistic Regulation of Hyphal Elongation by Hypoxia, CO2, and Nutrient Conditions Controls the Virulence of Candida albicans. Cell Host Microbe 2013, 14, 499–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; Pierson, D.L. Microbial Responses to Microgravity and Other Low-Shear Environments. Microbiol. Mol. Biol. Rev. 2004, 68, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, J.W.; Ott, C.M.; Zu Bentrup, K.H.; Ramamurthy, R.; Quick, L.; Porwollik, S.; Cheng, P.; McClelland, M.; Tsaprailis, G.; Radabaugh, T.; et al. Space flight alters bacterial gene expression and virulence and reveals a role for global regulator Hfq. Proc. Natl. Acad. Sci. USA 2007, 104, 16299–16304. [Google Scholar] [CrossRef] [Green Version]
- Noble, S.M.; Gianetti, B.A.; Witchley, J.N. Candida albicans cell-type switching and functional plasticity in the mammalian host. Nat. Rev. Genet. 2017, 15, 96–108. [Google Scholar] [CrossRef] [Green Version]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Desai, P.R.; Van Wijlick, L.; Kurtz, D.; Juchimiuk, M.; Ernst, J.F. Hypoxia and Temperature Regulated Morphogenesis in Candida albicans. PLoS Genet. 2015, 11, e1005447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawser, S.P.; Douglas, L.J. Resistance of Candida albicans biofilms to antifungal agents in vitro. Antimicrob. Agents Chemother. 1995, 39, 2128–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathé, L.; Van Dijck, P. Recent insights into Candida albicans biofilm resistance mechanisms. Curr. Genet. 2013, 59, 251–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheehan, K.B.; McInnerney, K.; Purevdorj-Gage, B.; Altenburg, S.D.; Hyman, E.L. Yeast genomic expression patterns in response to low-shear modeled microgravity. BMC Genom. 2007, 8, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altenburg, S.D.; Nielsen-Preiss, S.M.; Hyman, L.E. Increased Filamentous Growth of Candida albicans in Simulated Microgravity. Genom. Proteom. Bioinform. 2008, 6, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.; De Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.; Lebert, M.; et al. Ground-Based Facilities for Simulation of Microgravity: Organism-Specific Recommendations for Their Use, and Recommended Terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Searles, S.C.; Woolley, C.M.; Petersen, R.A.; Hyman, L.E.; Nielsen-Preiss, S.M. Modeled Microgravity Increases Filamentation, Biofilm Formation, Phenotypic Switching, and Antimicrobial Resistance inCandida albicans. Astrobiology 2011, 11, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Guadarrama, S.P.E.; Broadway, S.; Pyle, B. Pseudomonas aeruginosa growth and production of exotoxin A in static and modeled microgravity environments. Gravit. Space Biol. 2005, 18, 85–86. [Google Scholar]
- Hammond, T.G.; Hammond, J.M. Optimized suspension culture: The rotating-wall vessel. Am. J. Physiol. Physiol. 2001, 281, F12–F25. [Google Scholar] [CrossRef]
- Hall, D.; Hoshino, M. Effects of macromolecular crowding on intracellular diffusion from a single particle perspective. Biophys. Rev. 2010, 2, 39–53. [Google Scholar] [CrossRef] [Green Version]
- Crabbé, A.; Nielsen-Preiss, S.M.; Woolley, C.M.; Barrila, J.; Buchanan, K.; McCracken, J.; Inglis, D.O.; Searles, S.C.; Nelman-Gonzalez, M.A.; Ott, C.M.; et al. Spaceflight Enhances Cell Aggregation and Random Budding in Candida albicans. PLoS ONE 2013, 8, e80677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zea, L.; Larsen, M.; Estante, F.; Qvortrup, K.; Moeller, R.; De Oliveira, S.D.; Stodieck, L.; Klaus, D. Phenotypic Changes Exhibited by E. coli Cultured in Space. Front. Microbiol. 2017, 8, 1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawal, A.; Kirtley, M.L.; van Lier, C.; Erova, T.E.; Kozlova, E.V.; Sha, J.; Chopra, A.K.; Rosenzweig, J.A. The effects of modeled microgravity on growth kinetics, antibiotic susceptibility, cold growth, and the virulence potential of a Yersinia pestis ymoA-deficient mutant and its isogenic parental strain. Astrobiology 2013, 13, 821–832. [Google Scholar] [CrossRef] [Green Version]
- Hammond, T.G.; Stodieck, L.; Birdsall, H.H.; Becker, J.L.; Koenig, P.; Hammond, J.S.; Gunter, M.A.; Allen, P.L. Effects of Microgravity on the Virulence of Listeria monocytogenes, Enterococcus faecalis, Candida albicans, and Methicillin-Resistant Staphylococcus aureus. Astrobiology 2013, 13, 1081–1090. [Google Scholar] [CrossRef]
- Woolley, C.; Nielsen-Preiss, S.; Hyman, L. The response of Candida albicans to simulated microgravity. Gravit. Space Biol. 2010, 23, 93–94. [Google Scholar]
- Gulati, M.; Lohse, M.B.; Ennis, C.L.; Gonzalez, R.E.; Perry, A.M.; Bapat, P.; Arevalo, A.V.; Rodriguez, D.L.; Nobile, C.J. In VitroCulturing and Screening ofCandida albicansBiofilms. Curr. Protoc. Microbiol. 2018, 50, e60. [Google Scholar] [CrossRef]
- Touil, H.; Boucherit-Otmani, Z.; Boucherit, K. In vitro activity of antifungal combinations against planktonic and sessile cells of Candida albicans isolated from medical devices in an intensive care department. J. Mycol. Med. 2018, 28, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Simitsopoulou, M.; Peshkova, P.; Tasina, E.; Katragkou, A.; Kyrpitzi, D.; Velegraki, A.; Walsh, T.J.; Roilides, E. Species-Specific and Drug-Specific Differences in Susceptibility of Candida Biofilms to Echinocandins: Characterization of Less Common Bloodstream Isolates. Antimicrob. Agents Chemother. 2013, 57, 2562–2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pina-Vaz, C.; Sansonetty, F.; Rodrigues, A.G.; Costa-Oliveira, S.; Tavares, C.; Martinez-De-Oliveira, J. Cytometric approach for a rapid evaluation of susceptibility of Candida strains to antifungals. Clin. Microbiol. Infect. 2001, 7, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pore, R.S. Antibiotic susceptibility testing ofCandida albicans by flow cytometry. Curr. Microbiol. 1990, 20, 323–328. [Google Scholar] [CrossRef]
- Huang, B.; Liu, N.; Rong, X.; Ruan, J. Effects of simulated microgravity and spaceflight on morphological differentiation and secondary metabolism of Streptomyces coelicolor A3(2). Appl. Microbiol. Biotechnol. 2015, 99, 4409–4422. [Google Scholar] [CrossRef] [PubMed]
- Budd, S. Best Practices in Cryopreservation. 2016: American Type Culture Collection. Soc. Probl. 2020. [Google Scholar] [CrossRef]
- Bond, C. Cryopreservation of Yeast Cultures. Cryopreservation and Freeze-Drying Protocols. Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Buschelman, B.; Jones, R.N.; Pfaller, M.; Koontz, F.P.; Doern, G.V. Colony morphology of Candida spp. as a guide to species identification. Diagn. Microbiol. Infect. Dis. 1999, 35, 89–91. [Google Scholar] [CrossRef]
- Daniels, K.J.; Pujol, C.; Srikantha, T.; Soll, D.R. The “Finger,” a Unique Multicellular Morphology of Candida albicans Induced by CO2and Dependent upon the Ras1-Cyclic AMP Pathway. Eukaryot. Cell 2012, 11, 1257–1267. [Google Scholar] [CrossRef] [Green Version]
- Brown-Thomsen, J. Variability in Candida albicans (Robin Berkhout) Studies on morphology and biochemical activity. Hereditas 1968, 60, 355–398. [Google Scholar]
- Ahrens, J.P.; Daneo-Moore, M.R.; Buckley, H.R. Effects of culture density on the kinetics of germ tube formation in Candida albicans. J. Gen. Microbiol. 1983, 129, 3001–3006. [Google Scholar] [CrossRef] [Green Version]
- Zea, L.; Prasad, N.; Levy, S.E.; Stodieck, L.; Jones, A.; Shrestha, S.; Klaus, D. A Molecular Genetic Basis Explaining Altered Bacterial Behavior in Space. PLoS ONE 2016, 11, e0164359. [Google Scholar] [CrossRef] [PubMed]
- Baginski, M.; Sternal, K.; Czub, J.; Borowski, E. Molecular modelling of membrane activity of amphotericin B, a polyene macrolide antifungal antibiotic. ACTA Biochim. Pol. 2005, 52, 655–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.A.; De Sordi, L.; Maccallum, D.M.; Topal, H.; Eaton, R.; Bloor, J.W.; Robinson, G.K.; Levin, L.R.; Buck, J.; Wang, Y.; et al. CO2 Acts as a Signalling Molecule in Populations of the Fungal Pathogen Candida albicans. PLoS Pathog. 2010, 6, e1001193. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SpX CRS-1 | SpX CRS-4 | SpX CRS-16 | SpX CRS-17 | |
|---|---|---|---|---|
| Micro-6 | Micro-8 | Micro-14/FPA | Micro-14/FEP | |
| Scrub delay (days) | 0 | 1 | 1 | 4 |
| Launch date | 8 October 2012 | 21 September 2014 | 5 December 2018 | 4 May 2019 |
| Launch to dock (h) | 70.5 | 79.5 | 69.5 | 42 |
| Dock to sample access (h) ^ | 79 | 25 | 30 | 18 |
| Length of mission (days, launch to splashdown) | 21 | 35 | 39 | 31 |
| Experiment completion toundock (h) | 48 | 27 | 120 * | 264 # |
| Splashdown to home lab (h) | 70 | 68 | 84.5 | 66 |
| Sample | Flight-Generation Time (min) | Ground-Generation Time (min) |
|---|---|---|
| FPA-Early Low (Micro-6) | 155 | 159 |
| FPA-Early High (Micro-6) | 303 | 303 |
| FPA-Late Low (Micro-6) | 163 | 163 |
| FPA-Late High (Micro-6) | 300 | 310 |
| FPA-Low (Micro-14/FPA) | 113 | 129 |
| FPA-High (Micro-14/FPA) | 127 | 126 |
| FEP-Large (Micro-14/FEP) | 95 | 99 |
| FEP-Small (Micro-14/FEP) | 113 | 118 |
| Outcome Measure | FPA (Micro-6) | FPA (Micro-14) | FEP (Micro-14) |
|---|---|---|---|
| Cell Growth | F > T * | Trend F > T | F > T |
| Cell Density | Low | Intermediate | High |
| Filamentation | Low, no change F vs. T | Low, no change F vs. T | Very low, no change F vs. T |
| Colony Morphology | No change | No change | No change |
| Colony Size | No change | F > T | F > T |
| AmB Resistance | F > T | F > T | F > T |
| CAS Resistance | NA | NA | F = T |
| Biofilm | None | Trend T > F | NA |
| Freeze Stress Resistance | NA | T > F | T > F |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, S.; White, K.; Preiss, K.; Peart, D.; Gianoulias, K.; Juel, R.; Sutton, J.; McKinney, J.; Bender, J.; Pinc, G.; et al. Growth and Antifungal Resistance of the Pathogenic Yeast, Candida Albicans, in the Microgravity Environment of the International Space Station: An Aggregate of Multiple Flight Experiences. Life 2021, 11, 283. https://0-doi-org.brum.beds.ac.uk/10.3390/life11040283
Nielsen S, White K, Preiss K, Peart D, Gianoulias K, Juel R, Sutton J, McKinney J, Bender J, Pinc G, et al. Growth and Antifungal Resistance of the Pathogenic Yeast, Candida Albicans, in the Microgravity Environment of the International Space Station: An Aggregate of Multiple Flight Experiences. Life. 2021; 11(4):283. https://0-doi-org.brum.beds.ac.uk/10.3390/life11040283
Chicago/Turabian StyleNielsen, Sheila, Kenna White, Kyle Preiss, Dakota Peart, Kathryn Gianoulias, Rachel Juel, James Sutton, James McKinney, Jaclyn Bender, Gabrielle Pinc, and et al. 2021. "Growth and Antifungal Resistance of the Pathogenic Yeast, Candida Albicans, in the Microgravity Environment of the International Space Station: An Aggregate of Multiple Flight Experiences" Life 11, no. 4: 283. https://0-doi-org.brum.beds.ac.uk/10.3390/life11040283