
Swimming Training Does Not Affect the Recovery of Femoral Midshaft Structural and Mechanical Properties in Growing Diabetic Rats Treated with Insulin

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Body Weight and Blood Glucose
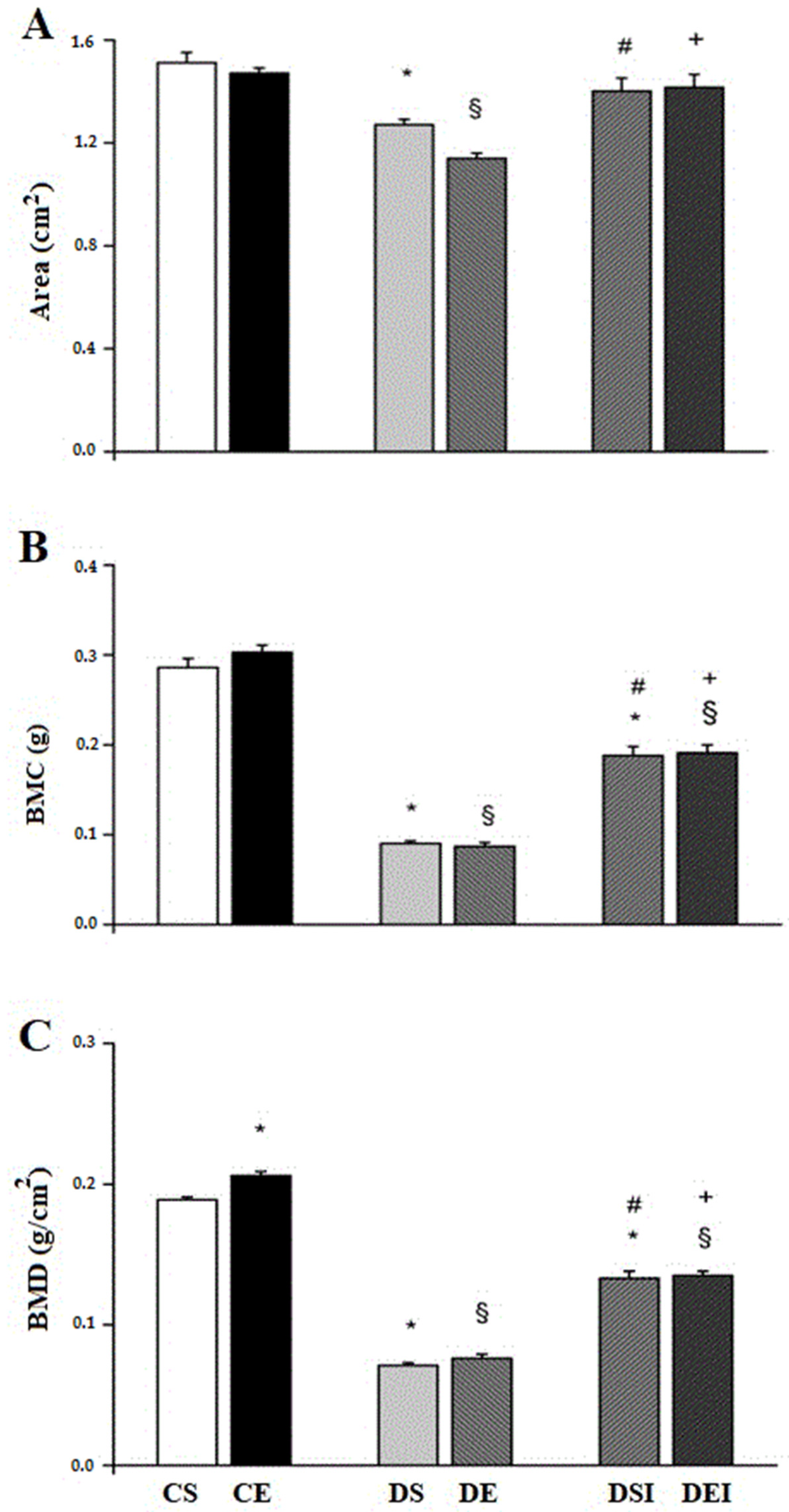
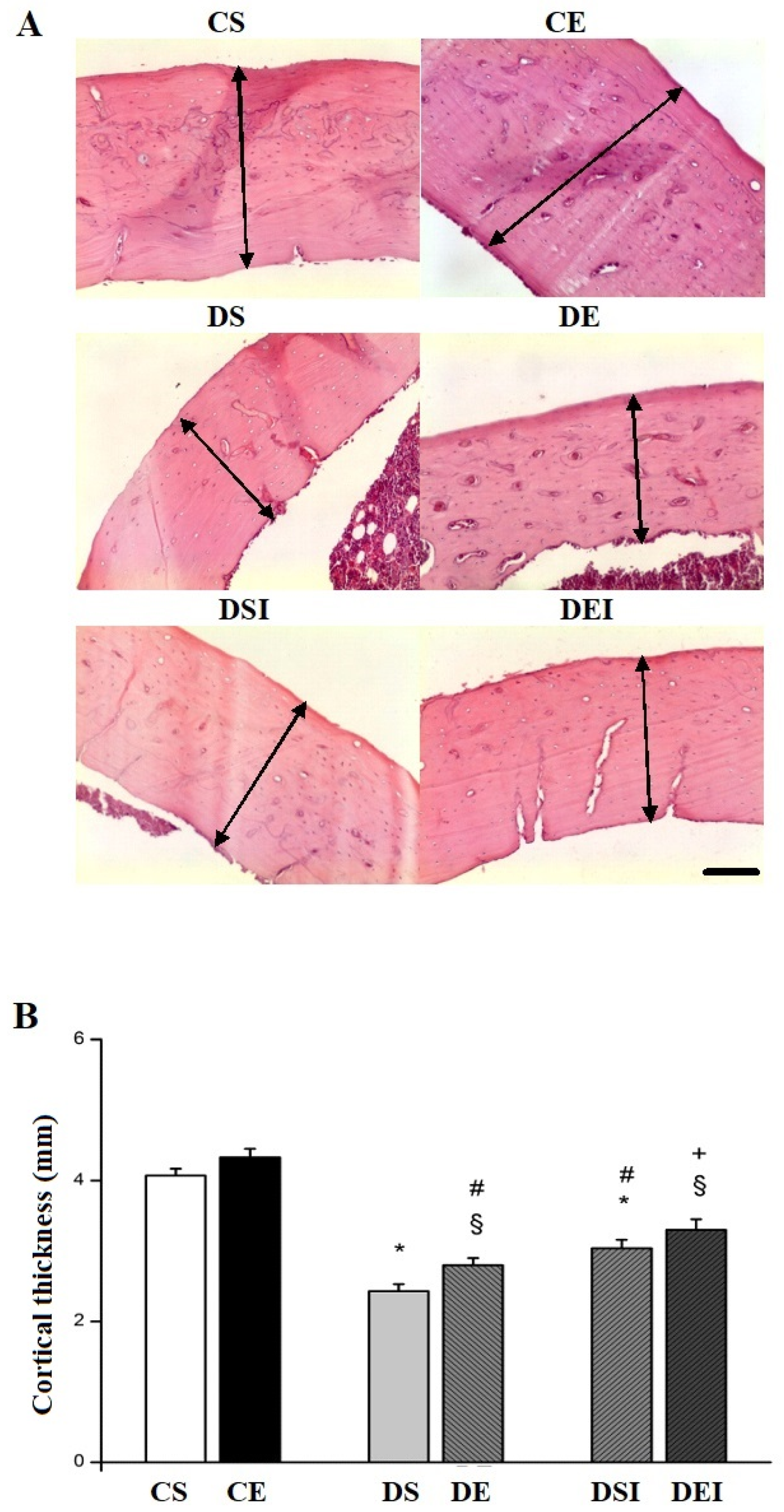
3.2. Femoral Midshaft Structural Properties
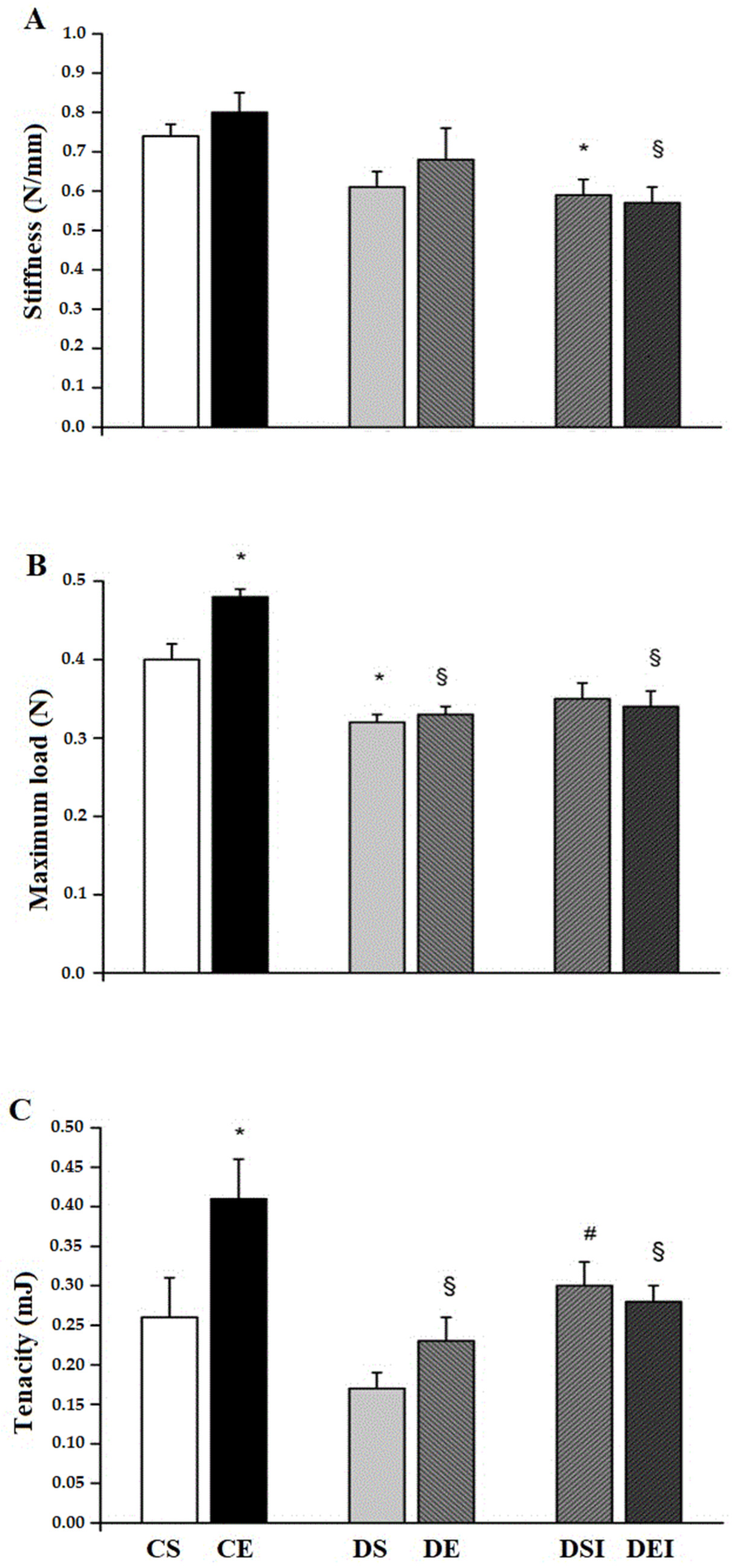
3.3. Femoral Midshaft Mechanical Properties
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Napoli, N.; Chandran, M.; Pierroz, D.D.; Abrahamsen, B.; Schwartz, A.V.; Ferrari, S.L. Mechanisms of diabetes mellitus-induced bone fragility. Nat. Rev. Endocrinol. 2017, 13, 208–219. [Google Scholar] [CrossRef]
- Sellmeyer, D.E.; Civitelli, R.; Hofbauer, L.C.; Khosla, S.; Lecka-Czernik, B.; Schwartz, A.V. Skeletal metabolism, fracture risk, and fracture outcomes in type 1 and type 2 diabetes. Diabetes 2016, 65, 1757–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestergaard, P.; Rejnmark, L.; Mosekilde, L. Diabetes and its complications and their relationship with risk of fractures in type 1 and 2 diabetes. Calcif. Tissue Int. 2009, 84, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Fujii, K.; Mori, Y.; Marumo, K. Role of collagen enzymatic and glycation induced cross-links as a determinant of bone quality in spontaneously diabetic BN/Kob rats. Osteoporos Int. 2006, 17, 1514–1523. [Google Scholar] [CrossRef]
- Saito, M.; Marumo, K. Collagen cross-links as a determinant of bone quality: A possible explanation for bone fragility in aging, osteoporosis, and diabetes mellitus. Osteoporos Int. 2010, 21, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Nyman, J.S.; Even, J.L.; Jo, C.; Herbert, E.G.; Murry, M.R.; Cockrell, G.E.; Wahl, E.C.; Bunn, R.C.; Lupkin, C.K., Jr.; Fowlkes, J.L.; et al. Increasing duration of type 1 diabetes perturbs the strength-structure relationship and increases brittleness of bone. Bone 2011, 48, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, N.; Pruzzob, R.; Eyzaguirrec, F.; Iñiguez, G.; López, P.; Mohr, J.; Perez-Bravo, F.; Cassorla, F.; Codner, E. Bone mass and sex steroids in postmenarcheal adolescents and adult women with type 1 diabetes mellitus. J. Diabetes Complicat. 2011, 25, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.C.H.; Zernicke, R.F.; Barnard, R.J. Effects of severe diabetes and insulin on the femoral neck of the immature rat. J. Orthop. Res. 1993, 11, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, L.C.; Brueck, C.C.; Singh, S.K.; Dobnig, H. Osteoporosis in patients with diabetes mellitus. J. Bone Min. Res. 2007, 22, 1317–1328. [Google Scholar] [CrossRef]
- Gomes, G.J.; Del Carlo, R.J.; da Silva, M.F.; Cunha, D.N.Q.; da Silva, E.; Silva, K.A.; Carneiro-Junior, M.A.; Prímola-Gomes, T.N.; Natali, A.J. Swimming training potentiates the recovery of femoral neck strength in young diabetic rats under insulin therapy. Clinics 2019, 74, e829. [Google Scholar] [CrossRef]
- Silva, K.A.; Del Carlo, R.J.; Matta, S.L.P.; Louzada, M.J.Q.; Rodrigues, A.C.; Silva, M.F.; Drummond, L.R.; Castro, C.A.; Silva, C.H.O.; Natali, A.J. Effects of swimming training on the femoral neck strength in growing rats with untreated streptozotocin-induced diabetes. Int. Sport Med J. 2014, 15, 445–457. [Google Scholar]
- Hamrick, M.W.; Samaddar, T.; Pennington, C.; McCormick, J. Increased muscle mass with myostatin deficiency improves gains in bone strength with exercise. J. Bone Miner. Res. 2006, 21, 477–483. [Google Scholar] [CrossRef]
- Erdal, N.; Gurgul, S.; Demirel, C.; Yildiz, A. The effect of insulin therapy on biomechanical deterioration of bone in streptozotocin (STZ)-induced type 1 diabetes mellitus in rats. Diabetes Res. Clin. Pract. 2012, 97, 461–467. [Google Scholar] [CrossRef]
- Beck, B.R.; Dalyb, R.M.; Singhc, M.A.F.; Taaffed, D.R. Exercise and Sports Science Australia (ESSA) position statement on exercise prescription for the prevention and management of osteoporosis. J. Sci. Med. Sport 2017, 20, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.I.; Sone, T.; Ohnaru, K.; Tanaka, K.; Fukunaga, M. Effect of swimming exercise on three-dimensional trabecular bone microarchitecture in ovariectomized rats. J. Appl. Physiol. 2015, 119, 990–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.-S.; Kim, S.-H.; Kim, J.-C. Effects of swimming exercise on high-fat diet-induced low bone mineral density and trabecular bone microstructure in rats. J. Exerc. Nutr. Biochem. 2017, 21, 48–55. [Google Scholar] [CrossRef]
- Gomez-Bruton, A.; Gonzalez-Aguero, A.; Gomez-Cabello, A.; Casajus, J.A.; Vicente-Rodriguez, G. Is bone tissue really affected by swimming? A systematic review. PLoS ONE 2013, 8, e70119. [Google Scholar]
- Portier, H.; Benaitreau, D.; Pallu, S. Does physical exercise always improve bone quality in rats? Life 2020, 10, 217. [Google Scholar] [CrossRef]
- Borer, K.T. Physical Activity in the prevention and amelioration of osteoporosis. Sports Med. 2005, 35, 779–830. [Google Scholar] [CrossRef]
- Mosekilde, L.; Thomsen, J.S.; Orhii, P.B.; McCarter, R.J.; Mejia, W.; Kalu, D.N. Additive effect of voluntary exercise and growth hormone treatment on bone strength assessed at four different skeletal sites in an aged rat model. Bone 1999, 24, 71–80. [Google Scholar] [CrossRef]
- Duarte, V.M.G.; Ramos, A.M.O.; Rezende, L.A.; Macedo, U.B.O.; Brandão-Neto, J.; Almeida, M.G.; Rezende, A.A. Osteopenia: A bone disorder associated with diabetes mellitus. J. Bone Miner. Metab. 2005, 23, 58–68. [Google Scholar] [CrossRef]
- McCarthy, A.D.; Uemura, T.; Etcheverry, S.B.; Cortizo, A.M. Advanced glycation endproducts interefere with integrin-mediated osteoblastic attachment to a type-I collagen matrix. Int. J. Biochem. Cell Biol. 2004, 36, 840–848. [Google Scholar] [CrossRef]
- Weiss, R.E.; Reddy, A.H. Influence of experimental diabetes and insulin on matrix-induced cartilage and bone differentiation. Am. J. Physiol. Endocrinol. Metab. 1980, 238, 200–207. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, X.; Wang, W.; Liu, J. Insulin stimulates osteoblast proliferation and differentiation through ERK and PI3K in MG-63 cells. Cell Biochem. Funct. 2010, 28, 334–341. [Google Scholar] [CrossRef]
- Maïmoun, L.; Sultan, C. Effect of physical activity on calcium homeostasis and calciotropic hormones: A review. Calcif. Tissue Int. 2009, 85, 277–286. [Google Scholar] [CrossRef]
- Boivin, G.; Bala, Y.; Doublier, A.; Farlay, D.; Ste-Marie, L.G.; Meunier, P.J.; Delmas, P.D. The role of mineralization and organic matrix in the microhardness of bone tissue from controls and osteoporotic patients. Bone 2008, 43, 532–538. [Google Scholar] [CrossRef]
- Kohn, D.H.; Sahar, N.D.; Wallace, J.M.; Golcuk, K.; Morris, M.D. Exercise alters mineral and matrix composition in the absence of adding new bone. Cells Tissues Organs. 2009, 189, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Robling, A.G.; Castillo, A.B.; Turner, C.H. Biomechanical and molecular regulation of bone remodeling. Annu. Rev. Biomed. Eng. 2006, 8, 455–498. [Google Scholar] [CrossRef]
- Saino, H.; Luther, F.; Carter, D.H.; Natali, A.J.; Turner, D.L.; Shahtaheri, S.M.; Aaron, J.E. Evidence for an extensive collagen Type III proximal domain in the rat fe.mur II. Expansion with exercise. Bone 2003, 32, 660–668. [Google Scholar] [CrossRef]
- Guadalupe-Grau, A.; Fuentes, T.; Guerra, B.; Calbet, A.L. Exercise and Bone Mass in Adults. Sports Med. 2009, 39, 439–468. [Google Scholar] [CrossRef]
- Blakytny, R.; Spraul, M.; Jude, E.B. The Diabetic Bone: A Cellular and Molecular Perspective. Int. J. Lower Ext. Wounds 2001, 10, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Moyer-Mileur, L.J.; Slater, H.; Jordan, K.C.; Murray, M.A. IGF-1 and IGF-binding proteins and bone mass, geometry, and strength: Relation to metabolic control in adolescent girls with type 1 diabetes. J. Bone Miner. Res. 2008, 23, 1884–1891. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Groups | N | Initial BW (g) | Final BW (g) | Initial BG (mg/dL) | BG 7 Days Post-STZ Injection (mg/dL) | Final BG (mg/dL) |
|---|---|---|---|---|---|---|
| CS | 10 | 88.61 ± 1.12 | 347.11 ± 6.74 §a | 71.52 ± 1.89 | - | 94.3 ± 6.41 b |
| CE | 10 | 90.80 ± 2.54 | 281.32 ± 11.14 *a | 72.20 ± 5.66 | - | 80.0 ± 3.17 |
| DS | 10 | 89.52 ± 1.82 | 160.40 ± 3.74 *a | 72.10 ± 5.78 | 409.00 ± 28.72 b | 573.3 ± 12.13 *bc |
| DE | 10 | 88.91 ± 3.81 | 164.11± 12.56 §a | 73.41 ± 4.26 | 367.61 ± 19.46 b | 504.4 ± 30.43 §bc |
| DSI | 10 | 91.80 ± 3.09 | 240.31 ± 6.17 *#a | 62.12 ± 1.08 | 392.12 ± 16.85 b | 312.1 ± 44.20 *#b |
| DEI | 10 | 88.60 ± 1.69 | 254.72 ± 6.27 §+a | 69.13 ± 3.11 | 365.02 ± 16.18 b | 339.6 ± 50.22 §+b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomes, G.d.J.; da Silva, M.F.; da Silva, E.; Del Carlo, R.J.; da Cunha, D.N.Q.; Carneiro-Junior, M.A.; Prímola-Gomes, T.N.; Natali, A.J. Swimming Training Does Not Affect the Recovery of Femoral Midshaft Structural and Mechanical Properties in Growing Diabetic Rats Treated with Insulin. Life 2021, 11, 786. https://0-doi-org.brum.beds.ac.uk/10.3390/life11080786
Gomes GdJ, da Silva MF, da Silva E, Del Carlo RJ, da Cunha DNQ, Carneiro-Junior MA, Prímola-Gomes TN, Natali AJ. Swimming Training Does Not Affect the Recovery of Femoral Midshaft Structural and Mechanical Properties in Growing Diabetic Rats Treated with Insulin. Life. 2021; 11(8):786. https://0-doi-org.brum.beds.ac.uk/10.3390/life11080786
Chicago/Turabian StyleGomes, Gilton de Jesus, Márcia Ferreira da Silva, Edson da Silva, Ricardo Junqueira Del Carlo, Daise Nunes Queiroz da Cunha, Miguel Araújo Carneiro-Junior, Thales Nicolau Prímola-Gomes, and Antônio José Natali. 2021. "Swimming Training Does Not Affect the Recovery of Femoral Midshaft Structural and Mechanical Properties in Growing Diabetic Rats Treated with Insulin" Life 11, no. 8: 786. https://0-doi-org.brum.beds.ac.uk/10.3390/life11080786