
Impact of Treadmill Interval Running on the Appearance of Zinc Finger Protein FHL2 in Bone Marrow Cells in a Rat Model: A Pilot Study

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experiment
2.2. Maximal Aerobic Speed Test
2.3. Training Protocol
2.4. Bone Histology
2.5. Imaging Processing
2.6. Statistical Analysis
3. Results
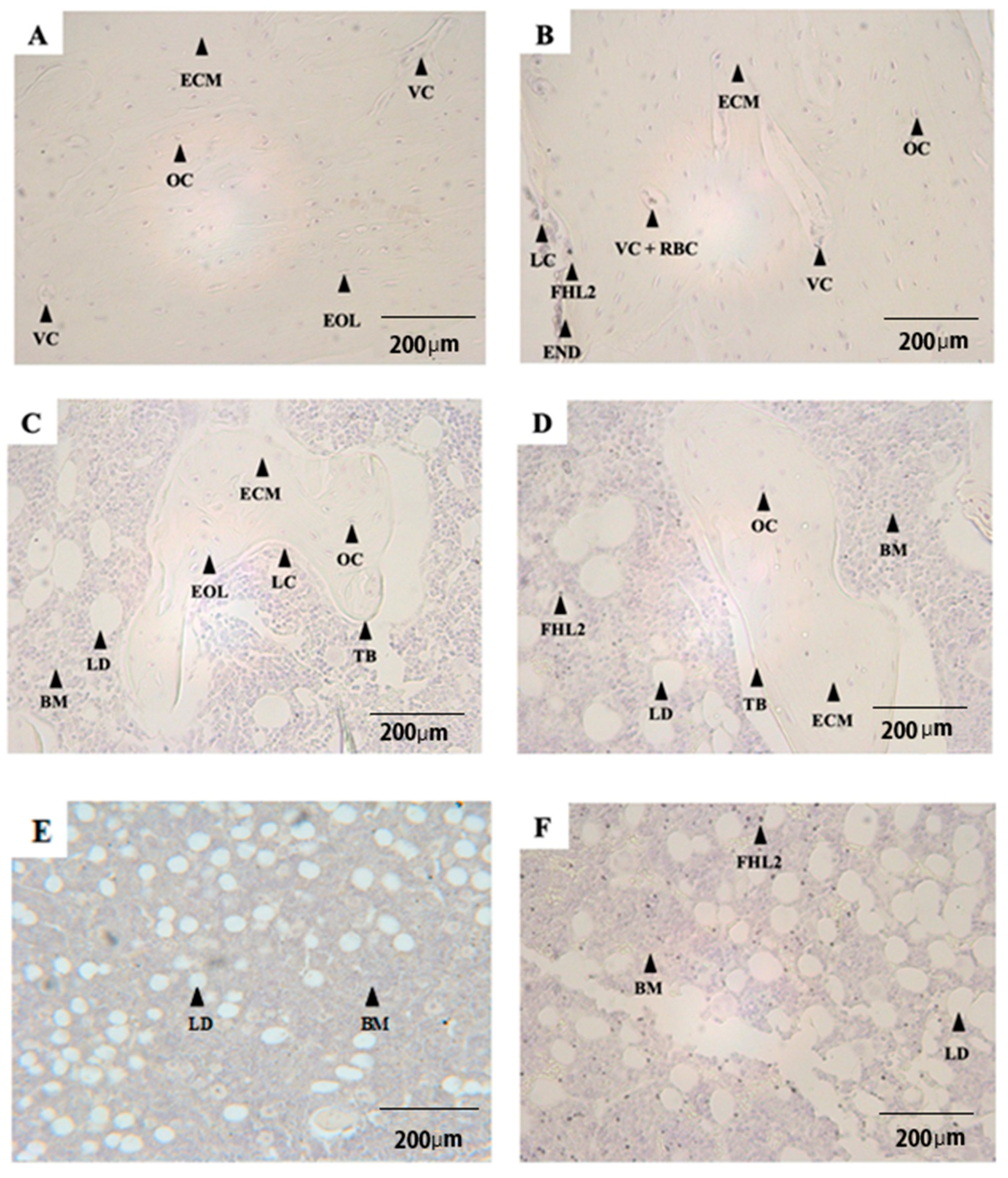
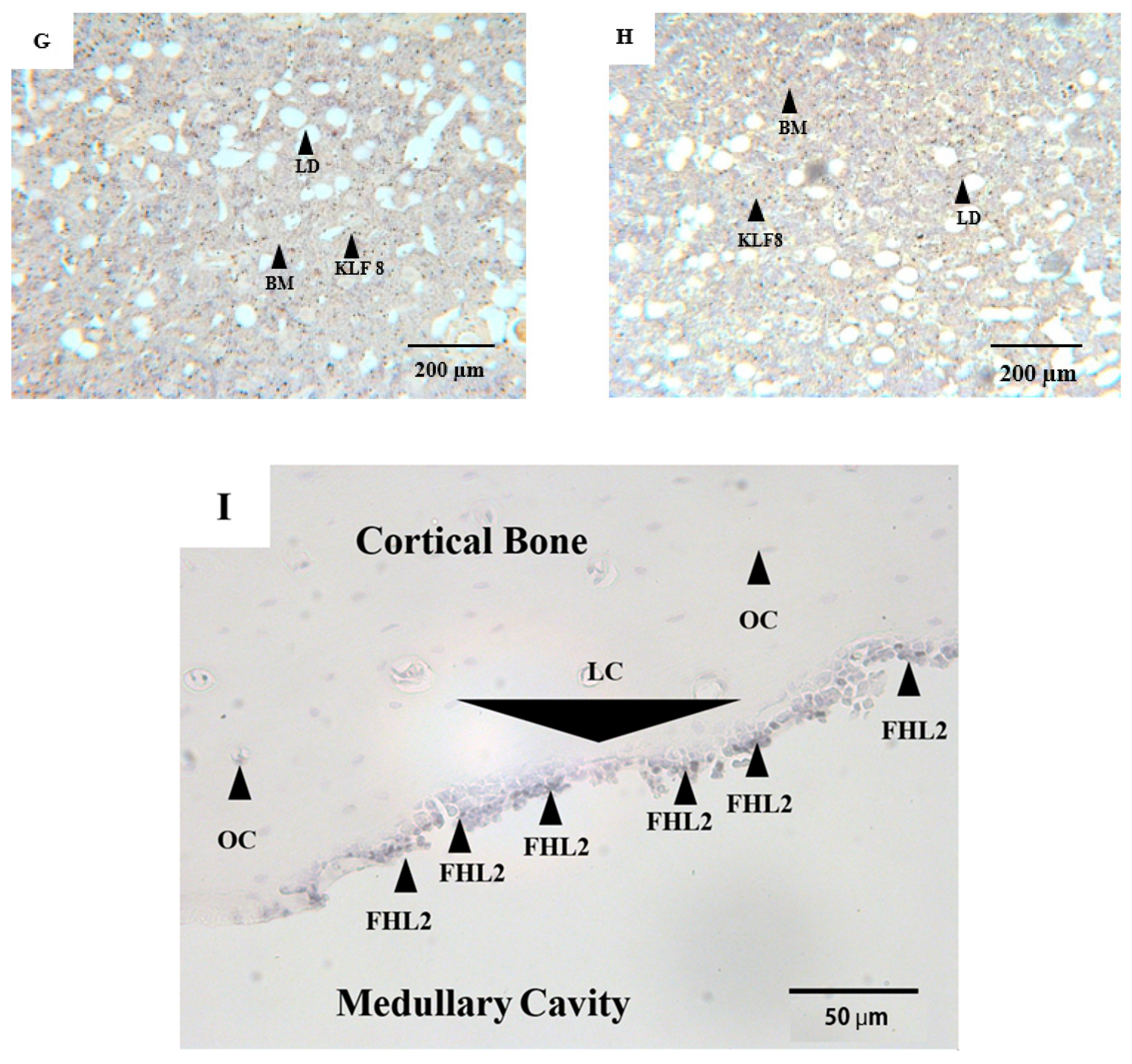
3.1. Qualitative Observations
3.2. Quantitative Analyses
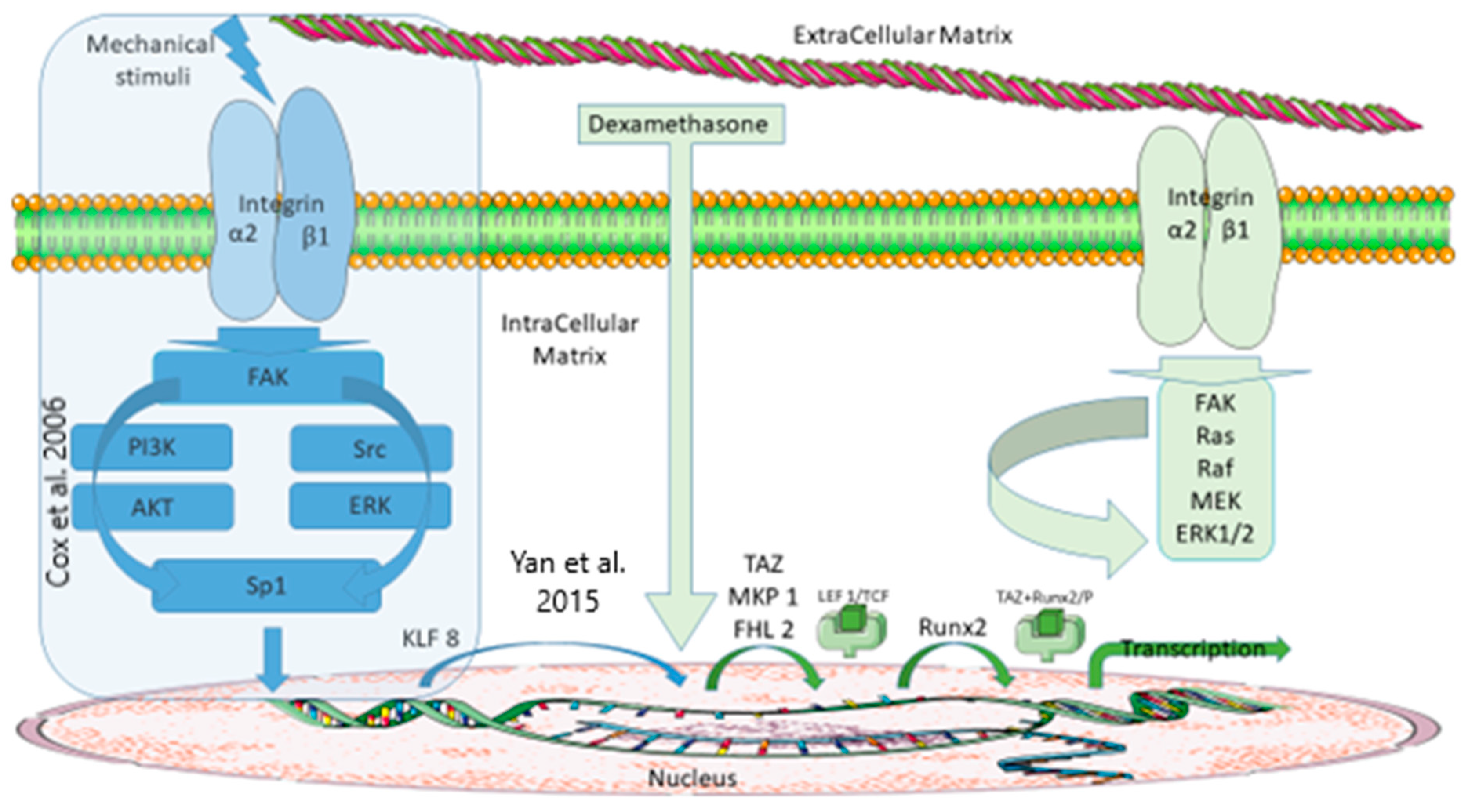
4. Discussion
How Could FHL2 Be Synthetized in Response to a Mechanical Stimulus?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smiles, W.J.; Hawley, J.A.; Camera, D.M. Effects of Skeletal Muscle Energy Availability on Protein Turnover Responses to Exercise. J. Exp. Biol. 2016, 219, 214–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawley, J.A.; Hargreaves, M.; Joyner, M.J.; Zierath, J.R. Integrative Biology of Exercise. Cell 2014, 159, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neufer, P.D.; Bamman, M.M.; Muoio, D.M.; Bouchard, C.; Cooper, D.M.; Goodpaster, B.H.; Booth, F.W.; Kohrt, W.M.; Gerszten, R.E.; Mattson, M.P.; et al. Understanding the Cellular and Molecular Mechanisms of Physical Activity-Induced Health Benefits. Cell Metab. 2015, 22, 4–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rindom, E.; Vissing, K. Mechanosensitive Molecular Networks Involved in Transducing Resistance Exercise-Signals into Muscle Protein Accretion. Front. Physiol. 2016, 7, 547. [Google Scholar] [CrossRef] [Green Version]
- Chicurel, M.E.; Chen, C.S.; Ingber, D.E. Cellular Control Lies in the Balance of Forces. Curr. Opin. Cell Biol. 1998, 10, 232–239. [Google Scholar] [CrossRef]
- Trepat, X.; Deng, L.; An, S.S.; Navajas, D.; Tschumperlin, D.J.; Gerthoffer, W.T.; Butler, J.P.; Fredberg, J.J. Universal Physical Responses to Stretch in the Living Cell. Nature 2007, 447, 592–595. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhou, J.; Chowdhury, F.; Cheng, J.; Wang, N.; Wang, F. Role of Mechanical Factors in Fate Decisions of Stem Cells. Regen. Med. 2011, 6, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, C. Muscle LIM Protein and Nesprin-1 in Mechanotransduction. Ph.D. Thesis, Freie Universität Berlin, Berlin, Germany, 2016. [Google Scholar]
- Vogel, V.; Sheetz, M. Local Force and Geometry Sensing Regulate Cell Functions. Nat. Rev. Mol. Cell Biol. 2006, 7, 265–275. [Google Scholar] [CrossRef]
- Sharili, A.S.; Connelly, J.T. Nucleocytoplasmic Shuttling: A Common Theme in Mechanotransduction. Biochem. Soc. Trans. 2014, 42, 645–649. [Google Scholar] [CrossRef]
- Portier, H.; Benaitreau, D.; Pallu, S. Does Physical Exercise Always Improve Bone Quality in Rats? Life 2020, 10, 217. [Google Scholar] [CrossRef]
- Courteix, D.; Lespessailles, E.; Peres, S.L.; Obert, P.; Germain, P.; Benhamou, C.L. Effect of Physical Training on Bone Mineral Density in Prepubertal Girls: A Comparative Study between Impact-Loading and Non-Impact-Loading Sports. Osteoporos. Int. 1998, 8, 152–158. [Google Scholar] [CrossRef]
- Taaffe, D.R.; Robinson, T.L.; Snow, C.M.; Marcus, R. High-Impact Exercise Promotes Bone Gain in Well-Trained Female Athletes. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1997, 12, 255–260. [Google Scholar] [CrossRef]
- Liu, Z.; Gao, J.; Gong, H. Effects of Treadmill with Different Intensities on Bone Quality and Muscle Properties in Adult Rats. Biomed. Eng. Online 2019, 18, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein-Nulend, J.; Bakker, A.D.; Bacabac, R.G.; Vatsa, A.; Weinbaum, S. Mechanosensation and Transduction in Osteocytes. Bone 2013, 54, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yuan, Y.; Wu, W.; Sun, Z.; Lei, L.; Fan, J.; Gao, B.; Zou, J. Medium-Intensity Treadmill Exercise Exerts Beneficial Effects on Bone Modeling Through Bone Marrow Mesenchymal Stromal Cells. Front. Cell Dev. Biol. 2020, 8, 600639. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise Review: Mesenchymal Stem Cells: Their Phenotype, Differentiation Capacity, Immunological Features, and Potential for Homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [Green Version]
- James, A.W. Review of Signaling Pathways Governing MSC Osteogenic and Adipogenic Differentiation. Scientifica 2013, 2013, 684736. [Google Scholar] [CrossRef] [PubMed]
- Bouzid, T.; Kim, E.; Riehl, B.D.; Esfahani, A.M.; Rosenbohm, J.; Yang, R.; Duan, B.; Lim, J.Y. The LINC Complex, Mechanotransduction, and Mesenchymal Stem Cell Function and Fate. J. Biol. Eng. 2019, 13, 68. [Google Scholar] [CrossRef] [PubMed]
- Rustad, K.C.; Wong, V.W.; Gurtner, G.C. The Role of Focal Adhesion Complexes in Fibroblast Mechanotransduction during Scar Formation. Differ. Res. Biol. Divers. 2013, 86, 87–91. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, Y.; Wang, M.; Zhao, S.; Zhao, Z.; Fang, J. Mechanotransduction Pathways in the Regulation of Cartilage Chondrocyte Homoeostasis. J. Cell. Mol. Med. 2020, 24, 5408–5419. [Google Scholar] [CrossRef] [Green Version]
- Yavropoulou, M.P.; Yovos, J.G. The Molecular Basis of Bone Mechanotransduction. J. Musculoskelet. Neuronal Interact. 2016, 16, 221–236. [Google Scholar] [PubMed]
- Lyon, R.C.; Zanella, F.; Omens, J.H.; Sheikh, F. Mechanotransduction in Cardiac Hypertrophy and Failure. Circ. Res. 2015, 116, 1462–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.-C.; Thampatty, B.P. An Introductory Review of Cell Mechanobiology. Biomech. Model. Mechanobiol. 2006, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Verrier, S.; Pallu, S.; Bareille, R.; Jonczyk, A.; Meyer, J.; Dard, M.; Amédée, J. Function of Linear and Cyclic RGD-Containing Peptides in Osteoprogenitor Cells Adhesion Process. Biomaterials 2002, 23, 585–596. [Google Scholar] [CrossRef]
- Anderson, C.A.; Kovar, D.R.; Gardel, M.L.; Winkelman, J.D. LIM Domain Proteins in Cell Mechanobiology. Cytoskeleton 2021, 78, 303–311. [Google Scholar] [CrossRef]
- Smith, M.A.; Hoffman, L.M.; Beckerle, M.C. LIM Proteins in Actin Cytoskeleton Mechanoresponse. Trends Cell Biol. 2014, 24, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dion, S.; Demattéi, M.-V.; Renault, S. Les Domaines à Doigts de Zinc: Vers La Modification de La Structure et de l’activité Des Génomes. Med. Sci. 2007, 23, 834–839. [Google Scholar] [CrossRef] [Green Version]
- The Human Protein Atlas. Available online: https://www.proteinatlas.org/ (accessed on 26 November 2021).
- Johannessen, M.; Møller, S.; Hansen, T.; Moens, U.; Van Ghelue, M. The Multifunctional Roles of the Four-and-a-Half-LIM Only Protein FHL2. Cell. Mol. Life Sci. CMLS 2006, 63, 268–284. [Google Scholar] [CrossRef]
- Masemann, D.; Dantas, R.L.; Sitnik, S.; Schied, T.; Nordhoff, C.; Ludwig, S.; Wixler, V. The Four-and-a-Half LIM Domain Protein 2 Supports Influenza A Virus–Induced Lung Inflammation by Restricting the Host Adaptive Immune Response. Am. J. Pathol. 2018, 188, 1236–1245. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Han, S.; Wang, Y.; Cui, C.; Zhu, Q.; Jiang, X.; Yang, C.; Du, H.; Yu, C.; Li, Q.; et al. The LIM-Only Protein FHL2 Is Involved in Autophagy to Regulate the Development of Skeletal Muscle Cell. Int. J. Biol. Sci. 2019, 15, 838–846. [Google Scholar] [CrossRef]
- Chu, P.-H.; Chen, J. The Novel Roles of Four and a Half LIM Proteins 1 and 2 in the Cardiovascular System. Chang Gung Med. J. 2011, 34, 127–134. [Google Scholar]
- Joos, H.; Albrecht, W.; Laufer, S.; Reichel, H.; Brenner, R.E. IL-1β Regulates FHL2 and Other Cytoskeleton-Related Genes in Human Chondrocytes. Mol. Med. 2008, 14, 150–159. [Google Scholar] [CrossRef] [Green Version]
- Brun, J.; Fromigué, O.; Dieudonné, F.-X.; Marty, C.; Chen, J.; Dahan, J.; Wei, Y.; Marie, P.J. The LIM-Only Protein FHL2 Controls Mesenchymal Cell Osteogenic Differentiation and Bone Formation through Wnt5a and Wnt10b. Bone 2013, 53, 6–12. [Google Scholar] [CrossRef]
- Duan, Y.; Qiu, Y.; Huang, X.; Dai, C.; Yang, J.; He, W. Deletion of FHL2 in Fibroblasts Attenuates Fibroblasts Activation and Kidney Fibrosis via Restraining TGF-Β1-Induced Wnt/β-Catenin Signaling. J. Mol. Med. 2020, 98, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Poli, C. Functional Analysis of the Transcriptional Co-Factor Fhl2 during Adipocyte Differentiation. Available online: https://freidok.uni-freiburg.de/data/2812 (accessed on 12 April 2021).
- Hamidouche, Z.; Haÿ, E.; Vaudin, P.; Charbord, P.; Schüle, R.; Marie, P.J.; Fromigué, O. FHL2 Mediates Dexamethasone-Induced Mesenchymal Cell Differentiation into Osteoblasts by Activating Wnt/Beta-Catenin Signaling-Dependent Runx2 Expression. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2008, 22, 3813–3822. [Google Scholar] [CrossRef]
- Günther, T.; Poli, C.; Müller, J.M.; Catala-Lehnen, P.; Schinke, T.; Yin, N.; Vomstein, S.; Amling, M.; Schüle, R. Fhl2 Deficiency Results in Osteopenia Due to Decreased Activity of Osteoblasts. EMBO J. 2005, 24, 3049–3056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakazawa, N.; Sathe, A.R.; Shivashankar, G.V.; Sheetz, M.P. Matrix Mechanics Controls FHL2 Movement to the Nucleus to Activate P21 Expression. Proc. Natl. Acad. Sci. USA 2016, 113, E6813–E6822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Wang, X.; Li, L.; Fan, R.; Chen, J.; Zhu, T.; Li, W.; Jiang, Y.; Mittal, N.; Wu, W.; et al. FHL2 Regulates Hematopoietic Stem Cell Functions under Stress Conditions. Leukemia 2015, 29, 615–624. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Bian, Z.C.; Yee, K.; Chen, B.P.C.; Chien, S.; Guan, J.-L. Identification of Transcription Factor KLF8 as a Downstream Target of Focal Adhesion Kinase in Its Regulation of Cyclin D1 and Cell Cycle Progression. Mol. Cell 2003, 11, 1503–1515. [Google Scholar] [CrossRef]
- Klf8 in UniProtKB. Available online: https://www.uniprot.org/uniprot/?query=KLF8&sort=score (accessed on 25 January 2022).
- Wang, X.; Urvalek, A.M.; Liu, J.; Zhao, J. Activation of KLF8 Transcription by Focal Adhesion Kinase in Human Ovarian Epithelial and Cancer Cells. J. Biol. Chem. 2008, 283, 13934–13942. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Kim, H.J.; Lee, Y.J.; Lee, M.-Y.; Choi, H.; Lee, H.; Kim, J. Krüppel-Like Factor KLF8 Plays a Critical Role in Adipocyte Differentiation. PLoS ONE 2012, 7, e52474. [Google Scholar] [CrossRef]
- Cox, B.D.; Natarajan, M.; Stettner, M.R.; Gladson, C.L. New Concepts Regarding Focal Adhesion Kinase Promotion of Cell Migration and Proliferation. J. Cell. Biochem. 2006, 99, 35–52. [Google Scholar] [CrossRef]
- Langenbach, F.; Handschel, J. Effects of Dexamethasone, Ascorbic Acid and β-Glycerophosphate on the Osteogenic Differentiation of Stem Cells in Vitro. Stem Cell Res. Ther. 2013, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Bourzac, C.; Bensidhoum, M.; Manassero, M.; Chappard, C.; Michoux, N.; Pallu, S.; Portier, H. Preventive Moderate Continuous Running-Exercise Conditioning Improves the Healing of Non-Critical Size Bone Defects in Male Wistar Rats: A Pilot Study Using ΜCT. Life 2020, 10, 308. [Google Scholar] [CrossRef]
- Lapidot, T.; Dar, A.; Kollet, O. How Do Stem Cells Find Their Way Home? Blood 2005, 106, 1901–1910. [Google Scholar] [CrossRef] [Green Version]
- Gonzalo-Encabo, P.; McNeil, J.; Boyne, D.J.; Courneya, K.S.; Friedenreich, C.M. Dose-Response Effects of Exercise on Bone Mineral Density and Content in Post-Menopausal Women. Scand. J. Med. Sci. Sports 2019, 29, 1121–1129. [Google Scholar] [CrossRef]
- Amaar, Y.G.; Thompson, G.R.; Linkhart, T.A.; Chen, S.-T.; Baylink, D.J.; Mohan, S. Insulin-like Growth Factor-Binding Protein 5 (IGFBP-5) Interacts with a Four and a Half LIM Protein 2 (FHL2). J. Biol. Chem. 2002, 277, 12053–12060. [Google Scholar] [CrossRef] [Green Version]
- Buyandelger, B.; Ng, K.-E.; Miocic, S.; Piotrowska, I.; Gunkel, S.; Ku, C.-H.; Knöll, R. MLP (Muscle LIM Protein) as a Stress Sensor in the Heart. Pflüg. Arch. Eur. J. Physiol. 2011, 462, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Jaalouk, D.E.; Lammerding, J. Mechanotransduction Gone Awry. Nat. Rev. Mol. Cell Biol. 2009, 10, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Ingber, D.E.; Wang, N.; Stamenovic, D. Tensegrity, Cellular Biophysics, and the Mechanics of Living Systems. Rep. Prog. Phys. 2014, 77, 046603. [Google Scholar] [CrossRef] [Green Version]
- Biggs, M.J.P.; Dalby, M.J. Focal Adhesions in Osteoneogenesis. Proc. Inst. Mech. Eng. 2010, 224, 1441–1453. [Google Scholar] [CrossRef] [Green Version]
- Tsimbouri, P.M. Adult Stem Cell Responses to Nanostimuli. J. Funct. Biomater. 2015, 6, 598–622. [Google Scholar] [CrossRef] [Green Version]
- Klug, A. The Discovery of Zinc Fingers and Their Development for Practical Applications in Gene Regulation and Genome Manipulation. Q. Rev. Biophys. 2010, 43, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Boateng, S.Y.; Senyo, S.E.; Qi, L.; Goldspink, P.H.; Russell, B. Myocyte Remodeling in Response to Hypertrophic Stimuli Requires Nucleocytoplasmic Shuttling of Muscle LIM Protein. J. Mol. Cell. Cardiol. 2009, 47, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Mehta, T.S.; Monzur, F.; Zhao, J. Determination of Nuclear Localization Signal Sequences for Krüppel-Like Factor 8. Transcr. Factors 2010, 647, 171–186. [Google Scholar] [CrossRef] [Green Version]
- Yi, X.; Zai, H.; Long, X.; Wang, X.; Li, W.; Li, Y. Krüppel-like Factor 8 Induces Epithelial-to-Mesenchymal Transition and Promotes Invasion of Pancreatic Cancer Cells through Transcriptional Activation of Four and a Half LIM-Only Protein 2. Oncol. Lett. 2017, 14, 4883–4889. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Zhang, W.; Wu, Y.; Wu, M.; Zhang, M.; Shi, X.; Zhao, J.; Nan, Q.; Chen, Y.; Wang, L.; et al. KLF8 Promotes Tumorigenesis, Invasion and Metastasis of Colorectal Cancer Cells by Transcriptional Activation of FHL2. Oncotarget 2015, 6, 25402–25417. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Shen, Z.; Tang, L.-N.; Zheng, S.-E.; Sun, Y.-J.; Min, D.-L.; Yao, Y. KLF8 Knockdown Suppresses Proliferation and Invasion in Human Osteosarcoma Cells. Mol. Med. Rep. 2014, 9, 1613–1617. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germain, A.; Bourzac, C.; Pichon, C.; Portier, H.; Pallu, S.; Germain, P. Impact of Treadmill Interval Running on the Appearance of Zinc Finger Protein FHL2 in Bone Marrow Cells in a Rat Model: A Pilot Study. Life 2022, 12, 528. https://0-doi-org.brum.beds.ac.uk/10.3390/life12040528
Germain A, Bourzac C, Pichon C, Portier H, Pallu S, Germain P. Impact of Treadmill Interval Running on the Appearance of Zinc Finger Protein FHL2 in Bone Marrow Cells in a Rat Model: A Pilot Study. Life. 2022; 12(4):528. https://0-doi-org.brum.beds.ac.uk/10.3390/life12040528
Chicago/Turabian StyleGermain, Alexandre, Celine Bourzac, Chantal Pichon, Hugues Portier, Stéphane Pallu, and Philippe Germain. 2022. "Impact of Treadmill Interval Running on the Appearance of Zinc Finger Protein FHL2 in Bone Marrow Cells in a Rat Model: A Pilot Study" Life 12, no. 4: 528. https://0-doi-org.brum.beds.ac.uk/10.3390/life12040528