
The Pressure Dependence of the Stability of the G-quadruplex Formed by d(TGGGGT)

Graduate Department of Pharmaceutical Sciences, Leslie Dan Faculty of Pharmacy, University of Toronto, Toronto, ON M5G1M2, Canada
*
Author to whom correspondence should be addressed.
Life 2022, 12(5), 765; https://0-doi-org.brum.beds.ac.uk/10.3390/life12050765
Submission received: 1 April 2022
/
Revised: 17 May 2022
/
Accepted: 18 May 2022
/
Published: 21 May 2022
(This article belongs to the Special Issue Biophysics of Nucleic Acids Celebrating the 75th Birthday of Professor Kenneth J. Breslauer)
Abstract
:The G-quadruplex (GQ), a tetrahelix formed by guanine-rich nucleic acid sequences, is a potential drug target for several diseases. Monomolecular GQs are stabilized by guanine tetrads and non-guanine regions that form loops. Hydrostatic pressure destabilizes the folded, monomolecular GQ structures. In this communication, we present data on the effect of pressure on the conformational stability of the tetramolecular GQ, d[5′-TGGGGT-3′]4. This molecule does not have loops linking the tetrads; thus, its physical properties presumably reflect those of the tetrads alone. Understanding the properties of the tetrads will aid in understanding the contribution of the other structural components to the stability of GQ DNA. By measuring UV light absorption, we have studied the effect of hydrostatic pressure on the thermal stability of the tetramolecular d[5′-TGGGGT-3′]4 in the presence of sodium ions. Our data show that, unlike monomolecular GQ, the temperature at which d[5′-TGGGGT-3′]4 dissociates to form the constituent monomers is nearly independent of pressure up to 200 MPa. This implies that there is no net molar volume difference (∆V) between the GQ and the unfolded random-coil states. This finding further suggests that the large negative ∆V values for the unfolding of monomolecular GQ are due to the presence of the loop regions in those structures.
1. Introduction
The guanine-quadruplex (GQ) is a secondary structure of DNA and RNA formed by molecules with sequences rich in guanines in the presence of some cations, such as sodium or potassium. The cation coordinates a tetrad of mutually hydrogen-bonded guanine residues that arrange in a plane, and these tetrads stack on one another to form the quadruplex. Intramolecular GQs form by the folding of a single DNA or RNA molecule, whereas intermolecular GQs contain two or four guanine-rich molecules [1,2,3]. Guanine-rich DNA sequences that form GQs in vitro are implicated in a number of human diseases and are situated in conserved regions in the genome across species [4,5,6,7,8,9,10,11]. The ability of GQs to self-assemble has led to their use in nanotechnology applications based on DNA [12,13,14].
Stability studies relying on changes in temperature, pH, and solvent parameters for GQ-containing solutions provide insight on the energetics of the folding process [15]. By studying the effect of pressure on GQ-containing systems, we can understand the role of water in the folding event [15,16,17,18,19]. The data to date show that the formation of monomolecular GQ structures is accompanied by a net release of water molecules into the bulk from the dehydration of metal cations and the formation of void volume by tetrad stacking [18,20,21]. The water-excluded voids, in turn, lead to a structure that is sensitive to pressure changes [15,22]. By analyzing the effect of pressure on the equilibrium between the folded and unfolded forms of the molecule, we can obtain the volumetric parameters that characterize the process [15,23,24,25]. Volumetric studies of GQs are important for understanding the hydration of these structures and the role the intracellular milieu may play in stabilizing or destabilizing these structures in vivo [26,27,28,29].
The effect of hydrostatic pressure on the conformational stability of monomolecular GQs has received some attention. In all cases, increasing the pressure leads to a destabilization of the folded structure to an oligonucleotide with no distinct secondary structure [15]. In addition, the role of the loops in the observed destabilization caused by pressure has also been investigated [21,30].
In this manuscript, we present data on the effect of hydrostatic pressure on the tetramolecular GQ formed by the hexanucleotide d[5′-TGGGGT-3′] (TG4T). The four-stranded quadruplex, d[5′-TGGGGT-3′]4 (TG4T-GQ), does not have any loop structures and only consists of four G-tetrads with single thymine residues at the 5′- and 3′- ends. Thus, the volumetric properties of this quadruplex should be representative of those of the G-tetrads that stabilize the structure. Our data show, rather surprisingly, that in contrast to the behaviour observed for monomolecular GQs, the thermal stability of the d[5′-TGGGGT-3′]4 structure is independent of pressure up to at least 200 MPa.
2. Results
2.1. Formation of TG4T-GQ
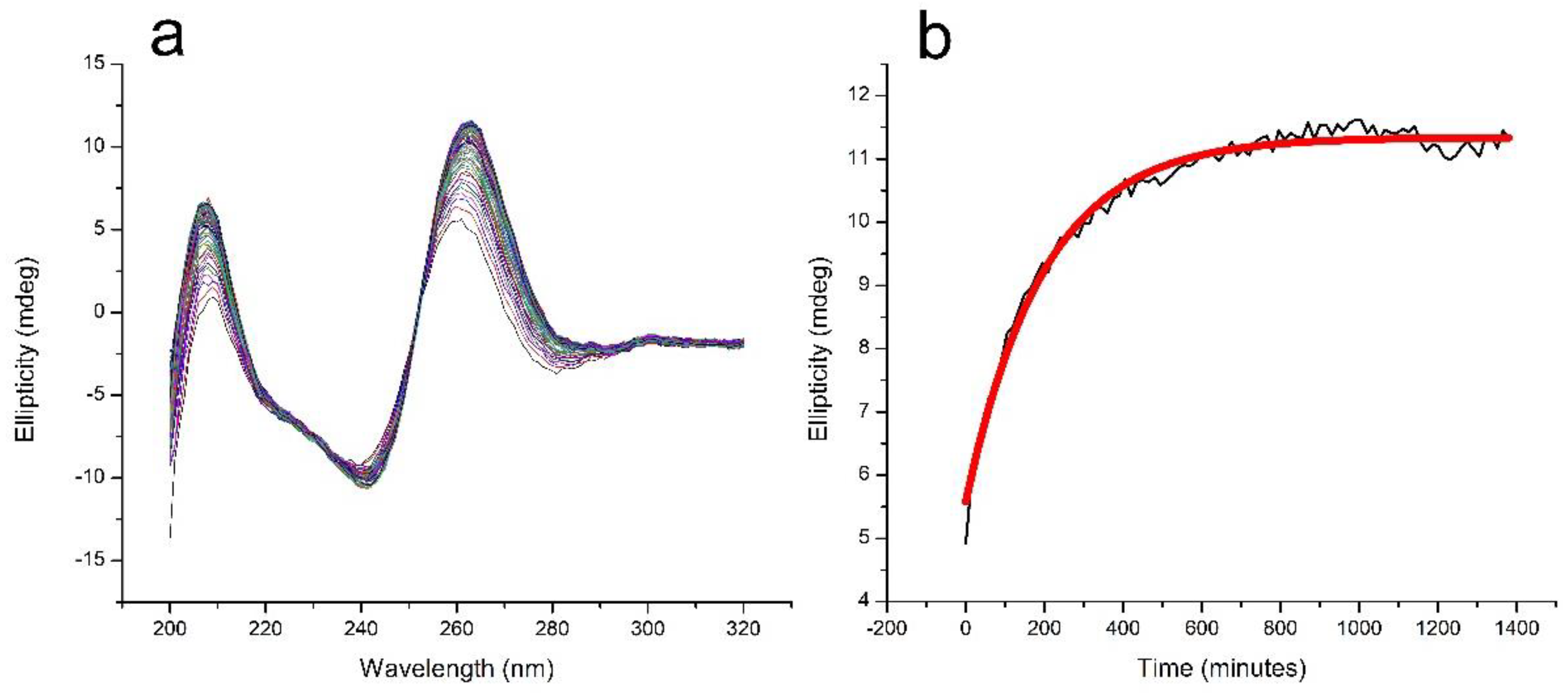
The formation of TG4T-GQ from its constituent oligonucleotides is slow [31,32]. We monitored the formation of TG4T-GQ in a solution containing 100 μM oligo concentration and 100 mM NaCl at room temperature and observed continued formation up until the last day the sample was measured at 22 days (data not shown). To facilitate the formation of TG4T-GQ for the purposes of studying the pressure dependence of its thermal stability, we increased the NaCl concentration to 1000 mM. Figure 1a presents 92 circular dichroism (CD) spectra taken at 15-min intervals over 1380 min (23 h) to demonstrate the formation of the quadruplex. The maximum in the CD spectrum, at 263 nm, is characteristic of the formation of the four-stranded structure, and a plot of the intensity of the CD signal at 263 nm as a function of time is shown in Figure 1b. The signal at 263 nm leveled off by the end of this incubation period of 23 h. It is important to note the results in Figure 1b are qualitative in nature, as we fit them to a simple exponential without consideration of a fourth-order reaction mechanism. Rather than elucidate the kinetics parameters, we were interested in determining a starting point for our experiments.
2.2. Measurements at Elevated Pressures
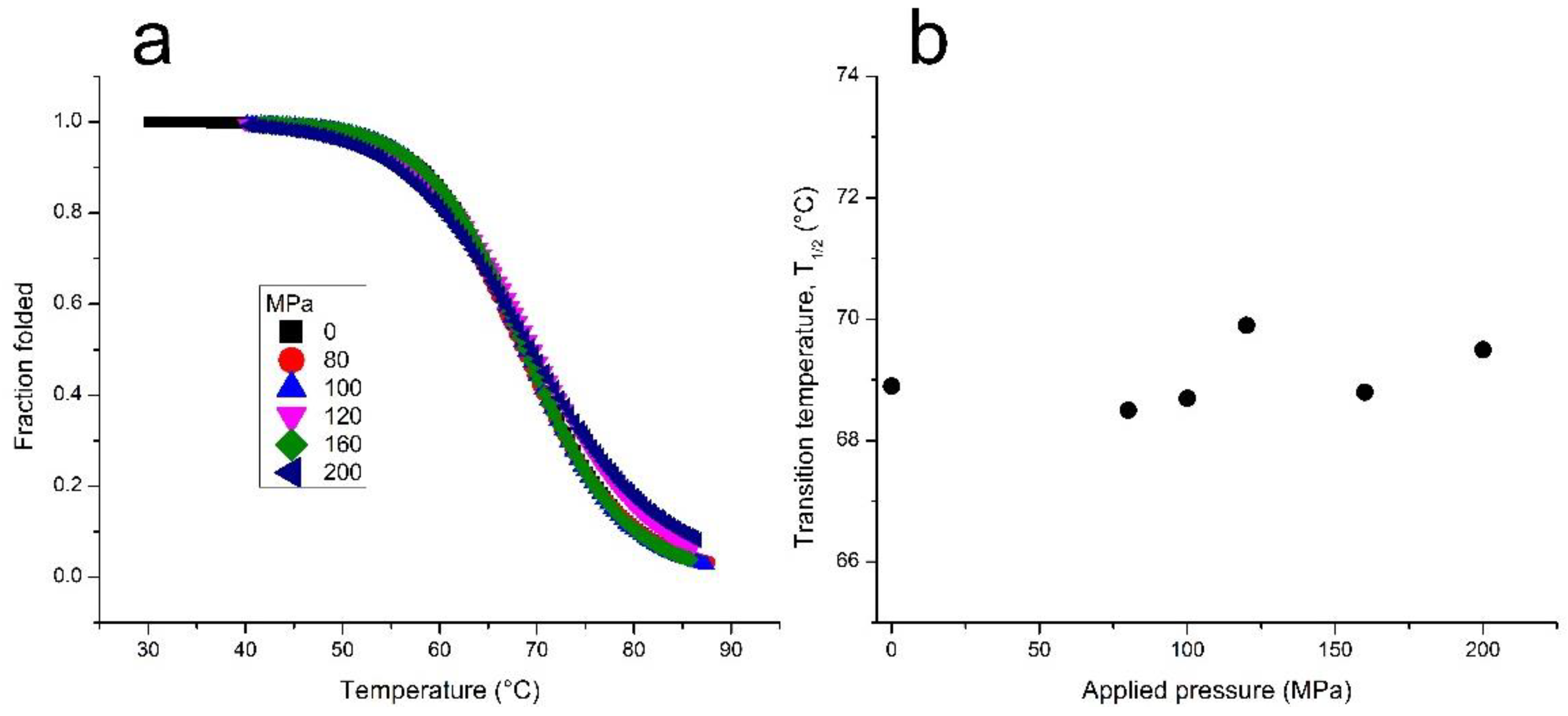
Next, we investigated the effect of pressure on the melting temperature (T1/2) of TG4T-GQ. Using the Clausius–Clapeyron equation, we calculated the volume change associated with the unfolding of the structure. According to the results shown in Figure 2 and Table 1, there was only a very modest change in the T1/2 of TG4T-GQ at pressures as high as 200 MPa. Based on these data, we calculated a molar volume change, ΔV, equal to −1.5 ± 2.3 cm3 mol−1 for the dissociation of TG4T-GQ into its four constituent oligonucleotides, d[5’-TGGGGT-3′]. Analysis of the enthalpy change upon dissociation assuming a single-step reaction is also reported in Table 1.
The four oligonucleotides in TG4T-GQ are oriented parallel to one another. We examined another parallel-stranded GQ formed from the oligo Pu22-T12T13, a variant of a 22-mer sequence proximal to the VEGF transcription initiation site [33,34]. We studied the dependence of the stability of the Pu22-T12T13-GQ on hydrostatic pressure. In comparison to the results for TG4T-GQ, the T1/2 of the parallel-stranded, monomolecular Pu22-T12T13-GQ showed significant destabilization with increasing pressure (Table 1). From our data, we calculated a molar volume change (ΔV) for the unfolding of Pu22-T12T13-GQ equal to −37.9 ± 10 cm3 mol−1.
3. Discussion
The number of studies that explore the effect of hydrostatic pressure on DNA G-quadruplex (GQ) structures is limited. In contrast to double-stranded conformations of nucleic acids, the G-quadruplexes have structural domains that may exhibit differential responses as a function of pressure. G-quadruplexes are stabilized by G-tetrads, which consist of four mutually hydrogen-bonded guanine residues that stack upon each other in the folded GQ structure. In monomolecular G-quadruplexes, successive G-tetrads are connected by nucleobases that are not guanine or cytosine residues. These bases that connect the stacked G-tetrads form the loops, which generally consist of two or three bases although loops with other numbers of bases are also possible. The bases in the loops do not form base-pairing interactions with other bases, and their existence leads to void volumes in folded G-quadruplex structures. The final component of the G-quadruplex structure are the cations that are coordinated by the O6 oxygen atoms of the guanine tetrads. When complexed with the G-tetrads, these ions are dehydrated, but upon denaturation of the G-quadruplex structure, they become rehydrated. The existence of structural voids and the rehydration of the cations released upon denaturation of the G-quadruplex structure are considered to be two of the primary causes of the destabilization of G-quadruplexes by pressure.
We can write the volume change arising from the denaturation of a monomolecular G-quadruplex as follows:
where ΔV(mmGQ) is the observed volume change for a monomolecular G-quadruplex such as HTel (human telomeric sequence) or VEGF (vascular endothelial growth factor); ΔV(tetrad) is the volume change that arises from the denaturation of the G-tetrads; ΔV(loops) is the contribution of the loops to the observed value of ΔV(mmGQ); and ΔV(ion) is the volume change that arises from the rehydration of the ions coordinated by the O6 oxygen atoms of the G-tetrads from the native structure.
ΔV(mmGQ) = ΔV(tetrad) + ΔV(loops) + ΔV(ion)
The role of the loops in the sensitivity of G-quadruplexes to pressure has been explored [21,30]. These studies show that the loops play an important role in the magnitude of the observed behaviour. Simple modifications to the loop regions result in significant changes in the unfolding volume of the monomolecular GQs formed by HTel and TBA (thrombin binding aptamer) GQs [15,21,30,35]. However, in all cases, regardless of the sequences of the loop, the volume change arising from the unfolding of the G-quadruplex is negative, i.e., the structure is destabilized by pressure. Upon the unfolding of the G-quadruplex structure, the bases that comprise the loops undergo a large change in the solvent-accessible surface area. In addition, the imperfect packing that arises from the structure of the loops in the folded structure leads to void volumes. These void volumes are lost upon the unfolding of the G-quadruplex structure.
To assess the contribution of the G-tetrads and ion release to the pressure dependence of the stability of G-quadruplexes, we have investigated the four-stranded complex formed by TG4T, which associates to form TG4T-GQ. The complex does not have loops; therefore, the observed volume change may be written as:
where ΔV(TG4T-GQ) is the observed volume change in the measurements carried out as a function of hydrostatic pressure. With knowledge of ΔV(TG4T-GQ) and ΔV(ion), we may then calculate the contribution of the G-tetrads to the measured volume change, ΔV(TG4T-GQ).
ΔV(TG4T-GQ) = ΔV(tetrad) + ΔV(ion)
TG4T-GQ has slow folding kinetics. Figure 1a,b show that in 1000 mM NaCl, a solution containing 100 μM TG4T formed over the course of 24 h with a half-time of formation of approximately 140 min. After incubating for 23 h, the CD spectrum was consistent with that of a G-quadruplex structure, and the spectra no longer changed with time. Because of the slow rate of formation of TG4T-GQ, each sample was subjected to only a single thermal denaturation.
When the TG4T-GQ samples were studied at different hydrostatic pressures, we did not observe a change in the stability of TG4T-GQ under the conditions of our measurements (see Figure 1b and Table 1). That is, we did not observe a statistically significant change in the T1/2 as a function of pressure (p) (i.e., ΔT1/2/Δp ~0 °C/MPa). This result suggests that either (1) the volume change arising from rehydration of the ions has nearly the same magnitude but opposite sign from the volume change arising from the disruption of the G-tetrads, or (2) the volume change of both processes is negligible.
The denaturation of the G-tetrads entails the loss of the hydrogen bonds between the guanine residues, the unstacking of the G-tetrads, and the release of bound cations. The guanine–guanine hydrogen bonds lost upon denaturation will be replaced by hydrogen bonds with water. However, the unstacking of the G-tetrads will cause an increase in the solvent-accessible surface area (SASA). This change in SASA will involve exposure of the aromatic bases to water and a negative volume change. The release of the cations bound to the O6 oxygen atoms of the guanine residues will lead to the formation of interactions between the ions and water. This is also a process that proceeds with a negative volume change.
Thus, ΔV(tetrad) and ΔV(ion) are both anticipated to be negative. The observation that the T1/2 of TG4T-GQ is independent of hydrostatic pressure implies that ΔV(tetrad) ≈ −ΔV(ion). In the absence of loops, which give rise to void volumes, most of the effect of hydrostatic pressure on the stability of TG4T-GQ will arise from the changes in the interactions of water with the associated and dissociated states of this system. The properties of water, including its partial molar volume, are sensitive to temperature. For example, the partial molar volume passes from negative values at temperatures below ~50 °C to small positive values above this temperature [36]. From this, we infer that the interactions between the released ions and water are weaker in the higher temperature regime. Similar considerations apply to the interactions of the aromatic bases with water. Taken together, our findings suggest that possibility (2), the volume change of both processes is negligible, is relevant in this system, that is, the volume change arising from the hydration of the released ions and the exposed bases tend toward zero at the T1/2 of TG4T-GQ.
The tetramolecular G-quadruplex structure formed from TG4T has strands in an all-parallel conformation, whereas the monomolecular quadruplex formed from HTel is non-parallel. For this reason, we also present data on the effect of pressure on the conformational stability of the G-quadruplex formed by Pu22-T12T13 (Table 1). Pu22-T12T13 forms a monomolecular structure with parallel strand orientation. The data show that the molar volume change of unfolding this G-quadruplex is large and negative, similar to the other monomolecular structures that have been studied [15]. The results from Pu22-T12T13 and other parallel-stranded GQs from the literature demonstrate that the parallel strand orientation is not a cause for the near-zero ΔV of TG4T-GQ [37]. Thus, we conclude that the behaviour exhibited by TG4T-GQ is not related to the orientation of the strands.
Extending the present findings to the behaviour of single-stranded oligonucleotides that can form tetrahelical GQ conformations suggests that the pressure dependence of these monomolecular structures arises mostly from the differential hydration of the loops in these cases. It would be useful to explore the behaviour of TG4T-GQ stabilized by other ions or other tetramolecular G-quadruplexes, such as d([5′-TGGGT-3′])4 or d([5′-TGGGGGT-3′])4.
In conclusion, we present data that show that the thermal denaturation of the four-stranded G-quadruplex, TG4T-GQ, does not depend on pressure in solutions containing 1000 mM sodium chloride. We attribute the finding that, within experimental error, ΔT1/2/Δp = 0 cm3 mol−1 to volume changes of both the disruption of the G-tetrads and the rehydration of the released cation tending toward zero at the temperature of the denaturation (~68 °C). This result suggests that the large, negative value of ΔV for the thermal denaturation of monomolecular G-quadruplex arises predominately from the presence of the loops in these structures and the void volumes to which they give rise.
4. Materials and Methods
We purchased the oligodeoxyribonucleotides TG4T (d[5′-TGGGGT-3′]) and Pu22-T12T13 (d[5′-CGGGGCGGGCCTTGGGCGGGGT-3′]) from ACGT (Toronto, ON, Canada). DNA was suspended in and dialyzed against Milli-Q® ultra-purified water (Millipore Milli-Q Biocel Water Purification System, Sigma-Aldrich, Oakville, ON, Canada), vacuum-dried, and stored at −20 °C. DNA concentration was determined by measuring the absorbance at 260 nm using a molar extinction coefficient of 57,800 M−1 cm−1 for TG4T and 200,400 M−1 cm−1 for Pu22-T12T13 (Cary model 300 Bio spectrophotometer, Varian Canada, Inc., Mississauga, ON, Canada) [31,38,39,40]. Stock 100 mM tris(hydroxymethyl)aminomethane (tris, min. 99.5%, Bioshop, Burlington, ON, Canada) buffer adjusted to pH 7.5 with HCl was prepared, filter-sterilized (0.22 μM pore size), and diluted to 10 mM as the solvent for the TG4T and NaCl (1000 mM sodium chloride; Bioshop, Burlington, ON, Canada) solutions. TG4T-GQ was formed by heating the DNA samples dissolved in buffer to 95 °C for five minutes and then allowing the sample to cool to room temperature overnight in a 1-L Dewar flask fitted with a loose lid. Pu22-T12T13 was also heated and cooled overnight. Pu22-T12T13 was prepared in a solution of 2 mM KCl (potassium chloride, min 99.5%), 10 mM phosphoric acid, and 0.1 mM EDTA (ethylenediaminetetraacetic acid), titrated to pH 7.0 with TBAOH (tetrabutylammonium hydroxide), all purchased from Sigma-Aldrich, Oakville, ON, Canada. Note that 2 mM KCl was chosen because Pu220T12T13 formation is complete at this concentration [34].
Circular dichroism (CD) spectra were collected on a JASCO model J-1100 CD spectropolarimeter (Jasco, Easton, MD, USA) at 25 °C. The average of at least two scans are reported for each spectrum. The kinetics results in Figure 1 were prepared by first adding salt to the GQ at room temperature in a 1 mm quartz cuvette, placing the sample in the CD instrument, heating to 95 °C and holding for 5 min, then cooling to 25 °C at a rate of 1 °C/minute. The spectra were then collected at 15-min intervals for 23 h.
For the pressure experiments, a 350-mL sample cuvette was positioned in an optical high-pressure cell. Silicon oil was used as the pressure-transmitting liquid [41]. The light absorption of the sample in the pressure cell was monitored on a Uvikon model 860 spectrophotometer (Kontron, Inc., Everett, MA, USA). The temperature of the pressure cell was maintained by a brass thermal jacket with a programmable circulating water bath. The temperature was increased at 0.5 °C/min or 0.9 °C/minute, and the sample temperature was measured with a thermocouple inserted into the pressure cell. Instrument control and data acquisition were achieved using a Windows PC running a Python program. The stability of the G-quadruplex structure was observed at several static pressures between atmospheric pressure and 200 MPa. As described in previous work, we calculated the melting temperature (Tm) as the mid-point of the normalized temperature-induced transition, and the enthalpy from the slope of the normalized transition using the van’t Hoff equation; this assumes that the unfolding is a single-step mechanism [42,43]. In the case of tetramolecular GQs, it is reported as the apparent melting temperature, T1/2 [31,44].
The molar volume change (ΔV) of the unfolding of GQs is calculated using the Clausius–Clapeyron equation,
where ΔH is the change in enthalpy at the transition, and p is the hydrostatic pressure [15,37,45].
Throughout this manuscript, we refer to the single-stranded oligonucleotide, d[5′-TGGGGT-3′], as TG4T. The quadruplex formed from the association of four strands of TG4T, (d[5′-TGGGGT-3′])4, is referred to as TG4T-GQ.
Author Contributions
Conceptualization, N.T. and R.B.M.J.; formal analysis, N.T. and T.K.; investigation, N.T., T.K. and U.N.F.; methodology, N.T. and R.B.M.J.; validation, N.T. and T.K.; visualization, N.T.; supervision, R.B.M.J.; writing, N.T. and R.B.M.J. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the Natural Science and Engineering Council of Canada (to R.B.M.J.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Tigran Chalikian for the use of the circular dichroism instrument.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Víglaský, V.; Bauer, L.; Tlučková, K. Structural Features of Intra- and Intermolecular G-Quadruplexes Derived from Telomeric Repeats. Biochemistry 2010, 49, 2110–2120. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, J.; Adhikari, S.; Balasubramanian, S. The Structure and Function of DNA G-Quadruplexes. Trends Chem. 2020, 2, 123–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, Topology and Structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Brosh, R.M. G-Quadruplex Nucleic Acids and Human Disease: G-Quadruplex Nucleic Acids and Human Disease. FEBS J. 2010, 277, 3470–3488. [Google Scholar] [CrossRef] [PubMed]
- Maizels, N. G4-associated Human Diseases. EMBO Rep. 2015, 16, 910–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharel, P.; Balaratnam, S.; Beals, N.; Basu, S. The Role of RNA G-quadruplexes in Human Diseases and Therapeutic Strategies. Wiley Interdiscip. Rev. RNA 2020, 11, e1568. [Google Scholar] [CrossRef]
- Capra, J.A.; Paeschke, K.; Singh, M.; Zakian, V.A. G-Quadruplex DNA Sequences Are Evolutionarily Conserved and Associated with Distinct Genomic Features in Saccharomyces Cerevisiae. PLoS Comput. Biol. 2010, 6, e1000861. [Google Scholar] [CrossRef]
- Huppert, J.L.; Balasubramanian, S. G-Quadruplexes in Promoters throughout the Human Genome. Nucleic Acids Res. 2007, 35, 406–413. [Google Scholar] [CrossRef]
- Nakken, S.; Rognes, T.; Hovig, E. The Disruptive Positions in Human G-Quadruplex Motifs Are Less Polymorphic and More Conserved than Their Neutral Counterparts. Nucleic Acids Res. 2009, 37, 5749–5756. [Google Scholar] [CrossRef] [Green Version]
- Eddy, J.; Maizels, N. Gene Function Correlates with Potential for G4 DNA Formation in the Human Genome. Nucleic Acids Res. 2006, 34, 3887–3896. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.; Halder, K.; Halder, R.; Yadav, V.K.; Rawal, P.; Thakur, R.K.; Mohd, F.; Sharma, A.; Chowdhury, S. Genome-Wide Computational and Expression Analyses Reveal G-Quadruplex DNA Motifs as Conserved Cis- Regulatory Elements in Human and Related Species. J. Med. Chem. 2008, 51, 5641–5649. [Google Scholar] [CrossRef] [PubMed]
- Mergny, J.-L.; Sen, D. DNA Quadruple Helices in Nanotechnology. Chem. Rev. 2019, 119, 6290–6325. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.T. G-Quartets 40 Years Later: From 5′-GMP to Molecular Biology and Supramolecular Chemistry. Angew. Chem. Int. Ed. 2004, 43, 668–698. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues Pontinha, A.D.; Chiorcea-Paquim, A.-M.; Eritja, R.; Oliveira-Brett, A.M. Quadruplex Nanostructures of d(TGGGGT): Influence of Sodium and Potassium Ions. Anal. Chem. 2014, 86, 5851–5857. [Google Scholar] [CrossRef]
- Chalikian, T.V.; Macgregor, R.B. Volumetric Properties of Four-Stranded DNA Structures. Biology 2021, 10, 813. [Google Scholar] [CrossRef]
- Miller, M.C.; Buscaglia, R.; Chaires, J.B.; Lane, A.N.; Trent, J.O. Hydration Is a Major Determinant of the G-Quadruplex Stability and Conformation of the Human Telomere 3′ Sequence of d(AG3(TTAG3)3). J. Am. Chem. Soc. 2010, 132, 17105–17107. [Google Scholar] [CrossRef]
- Aslanyan, L.; Ko, J.; Kim, B.G.; Vardanyan, I.; Dalyan, Y.B.; Chalikian, T.V. Effect of Urea on G-Quadruplex Stability. J. Phys. Chem. B 2017, 121, 6511–6519. [Google Scholar] [CrossRef]
- Miyoshi, D.; Karimata, H.; Sugimoto, N. Hydration Regulates Thermodynamics of G-Quadruplex Formation under Molecular Crowding Conditions. J. Am. Chem. Soc. 2006, 128, 7957–7963. [Google Scholar] [CrossRef]
- Li, K.; Yatsunyk, L.; Neidle, S. Water Spines and Networks in G-Quadruplex Structures. Nucleic Acids Res. 2021, 49, 519–528. [Google Scholar] [CrossRef]
- Kankia, B.I.; Marky, L.A. Folding of the Thrombin Aptamer into a G-Quadruplex with Sr2+: Stability, Heat, and Hydration. J. Am. Chem. Soc. 2001, 123, 10799–10804. [Google Scholar] [CrossRef]
- Li, Y.Y.; Dubins, D.N.; Le, D.M.N.T.; Leung, K.; Macgregor, R.B. The Role of Loops and Cation on the Volume of Unfolding of G-Quadruplexes Related to HTel. Biophys. Chem. 2017, 231, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Y.; Shek, Y.L.; Amiri, A.; Dubins, D.N.; Heerklotz, H.; Macgregor, R.B.; Chalikian, T.V. Volumetric Characterization of Sodium-Induced G-Quadruplex Formation. J. Am. Chem. Soc. 2011, 133, 4518–4526. [Google Scholar] [CrossRef] [PubMed]
- Akasaka, K. Probing Conformational Fluctuation of Proteins by Pressure Perturbation. Chem. Rev. 2006, 106, 1814–1835. [Google Scholar] [CrossRef] [PubMed]
- Smeller, L. Pressure–Temperature Phase Diagrams of Biomolecules. Biochim. Biophys. Acta BBA Protein Struct. Mol. Enzymol. 2002, 1595, 11–29. [Google Scholar] [CrossRef]
- Meersman, F.; McMillan, P.F. High Hydrostatic Pressure: A Probing Tool and a Necessary Parameter in Biophysical Chemistry. Chem. Commun. 2014, 50, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Tateishi-Karimata, H.; Takahashi, S.; Ohyama, T.; Sugimoto, N. Effect of Molecular Crowding on the Stability of RNA G-Quadruplexes with Various Numbers of Quartets and Lengths of Loops. Biochemistry 2020, 59, 2640–2649. [Google Scholar] [CrossRef] [PubMed]
- Petraccone, L.; Pagano, B.; Giancola, C. Studying the Effect of Crowding and Dehydration on DNA G-Quadruplexes. Methods 2012, 57, 76–83. [Google Scholar] [CrossRef]
- Strychalski, W.; Guy, R.D. Intracellular Pressure Dynamics in Blebbing Cells. Biophys. J. 2016, 110, 1168–1179. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Sun, S.X. Cellular Pressure and Volume Regulation and Implications for Cell Mechanics. Biophys. J. 2013, 105, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S.; Sugimoto, N. Volumetric Contributions of Loop Regions of G-Quadruplex DNA to the Formation of the Tertiary Structure. Biophys. Chem. 2017, 231, 146–154. [Google Scholar] [CrossRef]
- Mergny, J.-L. Kinetics of Tetramolecular Quadruplexes. Nucleic Acids Res. 2005, 33, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Bardin, C.; Leroy, J.L. The Formation Pathway of Tetramolecular G-Quadruplexes. Nucleic Acids Res. 2008, 36, 477–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, P.; Hatzakis, E.; Guo, K.; Carver, M.; Yang, D. Solution Structure of the Major G-Quadruplex Formed in the Human VEGF Promoter in K+: Insights into Loop Interactions of the Parallel G-Quadruplexes. Nucleic Acids Res. 2013, 41, 10584–10592. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Long, J.; Dubins, D.N.; Chalikian, T.V. Ionic Effects on VEGF G-Quadruplex Stability. J. Phys. Chem. B 2016, 120, 4963–4971. [Google Scholar] [CrossRef] [PubMed]
- Olsen, C.M.; Lee, H.-T.; Marky, L.A. Unfolding Thermodynamics of Intramolecular G-Quadruplexes: Base Sequence Contributions of the Loops. J. Phys. Chem. B 2009, 113, 2587–2595. [Google Scholar] [CrossRef] [PubMed]
- Marcus, Y. The Standard Partial Molar Volumes of Ions in Solution. Part 4. Ionic Volumes in Water at 0–100 °C. J. Phys. Chem. B 2009, 113, 10285–10291. [Google Scholar] [CrossRef]
- Molnár, O.R.; Somkuti, J.; Smeller, L. Negative Volume Changes of Human G-Quadruplexes at Unfolding. Heliyon 2020, 6, e05702. [Google Scholar] [CrossRef]
- Warshaw, M.M.; Tinoco, I. Optical Properties of Sixteen Dinucleoside Phosphates. J. Mol. Biol. 1966, 20, 29–38. [Google Scholar] [CrossRef]
- Cantor, C.R.; Warshaw, M.M.; Shapiro, H. Oligonucleotide Interactions. III. Circular Dichroism Studies of the Conformation of Deoxyoligonucleolides. Biopolymers 1970, 9, 1059–1077. [Google Scholar] [CrossRef]
- Cavaluzzi, M.J.; Borer, P.N. Revised UV Extinction Coefficients for Nucleoside-5′-Monophosphates and Unpaired DNA and RNA. Nucleic Acids Res. 2004, 32, e13. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Q.; Macgregor, R.B. A Temperature-Regulated Iso-Hyperbaric Spectrophotometer: Construction and Performance Characteristics. Anal. Biochem. 1993, 211, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Mergny, J.; Lacroix, L. UV Melting of G-Quadruplexes. Curr. Protoc. Nucleic Acid Chem. 2009, 37. [Google Scholar] [CrossRef] [PubMed]
- Marky, L.A.; Breslauer, K.J. Calculating Thermodynamic Data for Transitions of Any Molecularity from Equilibrium Melting Curves. Biopolymers 1987, 26, 1601–1620. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, J.R.; Davis, P.W.; Freier, S.M. Kinetics of G-Quartet-Mediated Tetramer Formation. Biochemistry 1996, 35, 8002–8008. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Sugimoto, N. Effect of Pressure on Thermal Stability of G-Quadruplex DNA and Double-Stranded DNA Structures. Molecules 2013, 18, 13297–13319. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Kinetics of the formation of TG4T-GQ in aqueous solution containing 100 μM oligo and 1000 mM NaCl, 10 mM tris, pH 7.5. (a) The evolution of the CD spectrum of TG4T-GQ at 25 °C over time; a spectrum was acquired every 15 min for 23 h. (b) Plot of the ellipticity at 263 nm from (a) as a function of time, showing the approach to equilibrium in the formation of TG4T-GQ. The half-time of the formation process is approximately 140 min. It must be noted that these kinetic results are merely a means of estimating the time at which most of the sample has folded into a GQ to establish a starting point for our pressure experiments. The data points were fit to a simple exponential without consideration of the dead time or fourth-order reaction mechanism. Refer to the paper by Mergny and colleagues on tetramolecular kinetics for a more comprehensive kinetic analysis of four-stranded GQ [31].
Figure 1.
Kinetics of the formation of TG4T-GQ in aqueous solution containing 100 μM oligo and 1000 mM NaCl, 10 mM tris, pH 7.5. (a) The evolution of the CD spectrum of TG4T-GQ at 25 °C over time; a spectrum was acquired every 15 min for 23 h. (b) Plot of the ellipticity at 263 nm from (a) as a function of time, showing the approach to equilibrium in the formation of TG4T-GQ. The half-time of the formation process is approximately 140 min. It must be noted that these kinetic results are merely a means of estimating the time at which most of the sample has folded into a GQ to establish a starting point for our pressure experiments. The data points were fit to a simple exponential without consideration of the dead time or fourth-order reaction mechanism. Refer to the paper by Mergny and colleagues on tetramolecular kinetics for a more comprehensive kinetic analysis of four-stranded GQ [31].

Figure 2.
(a) Thermal denaturation of TG4T-GQ at different pressures. Fresh samples were used for each measurement. The samples contained 100 μM TG4T, 1000 mM NaCl, 10 mM tris, pH 7.5. (b) The pressure dependence of the melting temperature for the TG4T-GQ. The error in the T1/2 measurements is ±0.5 °C.
Figure 2.
(a) Thermal denaturation of TG4T-GQ at different pressures. Fresh samples were used for each measurement. The samples contained 100 μM TG4T, 1000 mM NaCl, 10 mM tris, pH 7.5. (b) The pressure dependence of the melting temperature for the TG4T-GQ. The error in the T1/2 measurements is ±0.5 °C.


{kind=link}
{kind=link}
Table 1.
The effect of hydrostatic pressure on the thermal denaturation of TG4T-GQ a and the G-quadruplex formed by Pu22-T12T13 b.
Table 1.
The effect of hydrostatic pressure on the thermal denaturation of TG4T-GQ a and the G-quadruplex formed by Pu22-T12T13 b.
| GQ | Applied Pressure | T1/2 (°C) * | ΔH (kJ) | ΔV (cm3 mol−1) |
|---|---|---|---|---|
| TG4T | 0 | 68.9 | 240 ± 14 | −1.5 ± 2.3 |
| 80 | 68.5 | 270 ± 16 | ||
| 100 | 68.7 | 300 ± 19 | ||
| 120 | 69.9 | 250 ± 19 | ||
| 160 | 68.8 | 280 ± 25 | ||
| 200 | 69.5 | 270 ± 18 | ||
| Pu22-T12T13 | 0 | 71.8 | 190 ± 23 | −37.9 ± 10 ** |
| 80 | 69.9 | 120 ± 19 | ||
| 180 | 60.7 | 130 ± 26 |
a TG4T samples: 100 μM TG4T, 1000 mM NaCl, 10 mM tris, pH 7.5. b Pu22-T12T13 samples: 100 µM Pu22-T12T13 in 2 mM KCl, 10 mM phosphoric acid, 0.1 mM EDTA, pH 7.0. * TG4T samples were heated at 0.9 °C/min, while Pu22-T12T13 samples were heated at 0.5 °C/min. Heating rates of 0.1, 0.5, and 0.9 °C/min did not influence the results for TG4T-GQ (unpublished). We present the data obtained with a 0.9 °C/min heating rate simply because it was the most complete set of data for TG4T. ** This error is an estimate based on prior publications with our pressure instrument [21].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tariq, N.; Kume, T.; Feroze, U.N.; Macgregor, R.B., Jr. The Pressure Dependence of the Stability of the G-quadruplex Formed by d(TGGGGT). Life 2022, 12, 765. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050765
AMA Style
Tariq N, Kume T, Feroze UN, Macgregor RB Jr. The Pressure Dependence of the Stability of the G-quadruplex Formed by d(TGGGGT). Life. 2022; 12(5):765. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050765
Chicago/Turabian StyleTariq, Nabeel, Takuma Kume, Ujala N. Feroze, and Robert B. Macgregor, Jr. 2022. "The Pressure Dependence of the Stability of the G-quadruplex Formed by d(TGGGGT)" Life 12, no. 5: 765. https://0-doi-org.brum.beds.ac.uk/10.3390/life12050765
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.