Geochemistry and the Origin of Life: From Extraterrestrial Processes, Chemical Evolution on Earth, Fossilized Life’s Records, to Natures of the Extant Life

 ,
,  , ,
, , 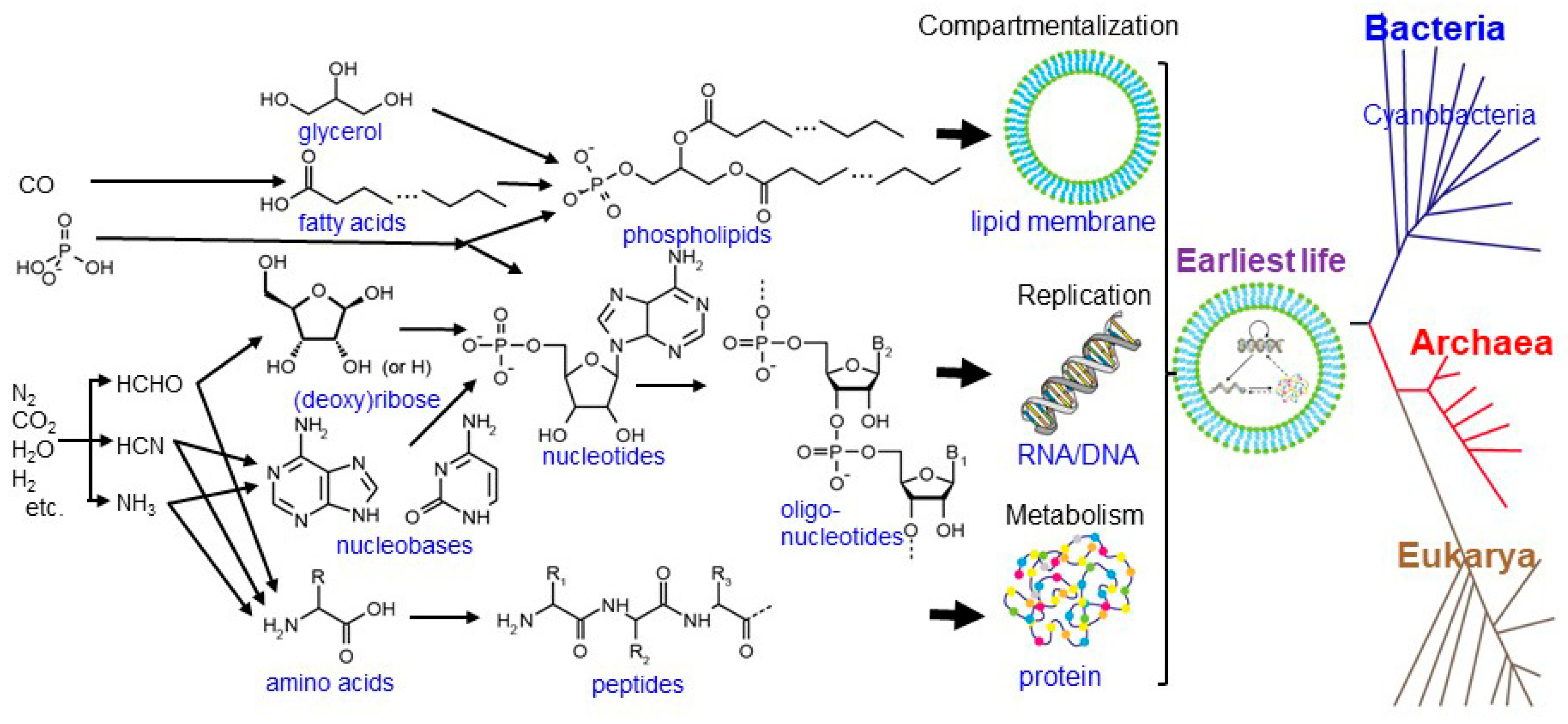{kind=link}
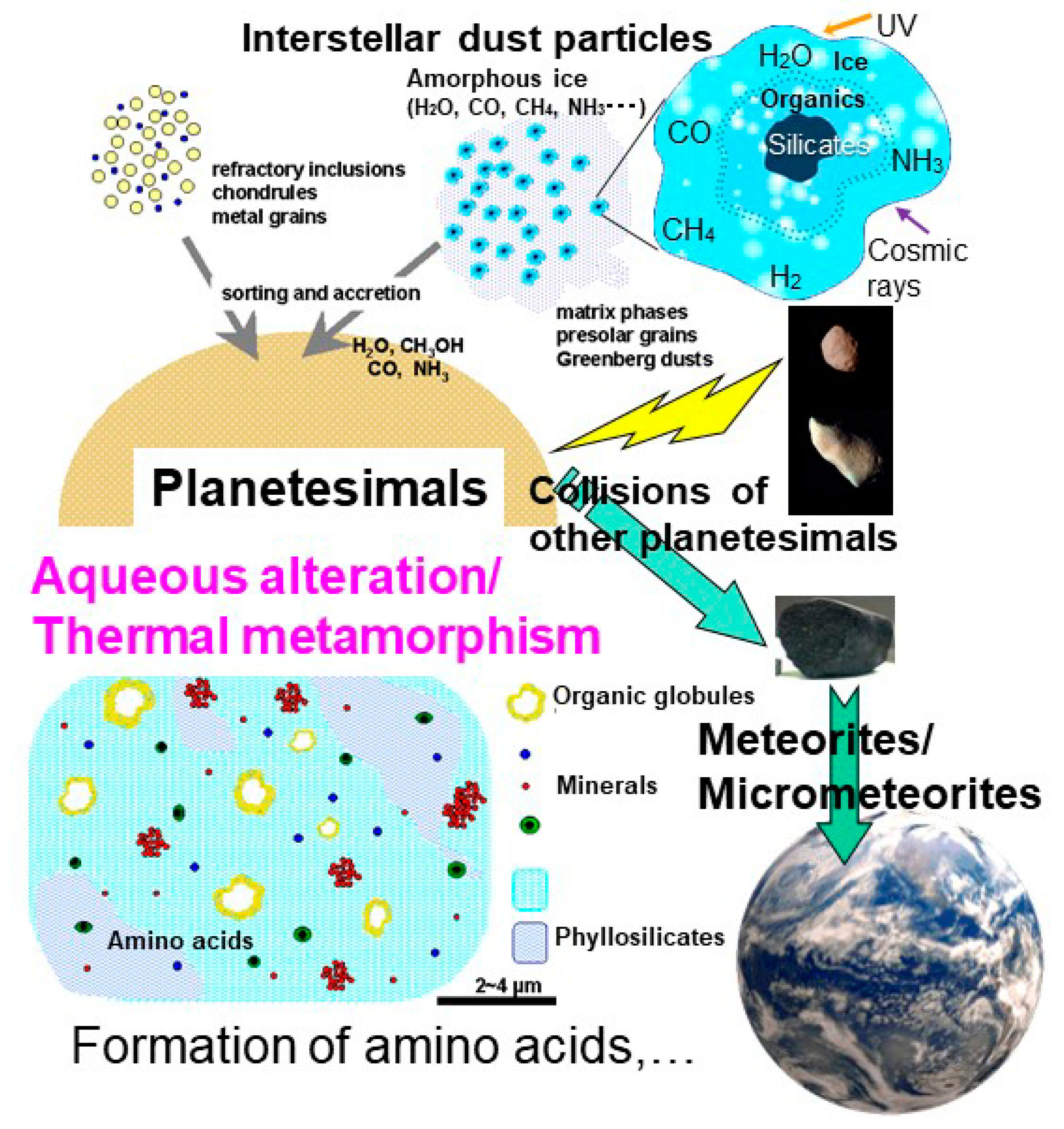{kind=link}
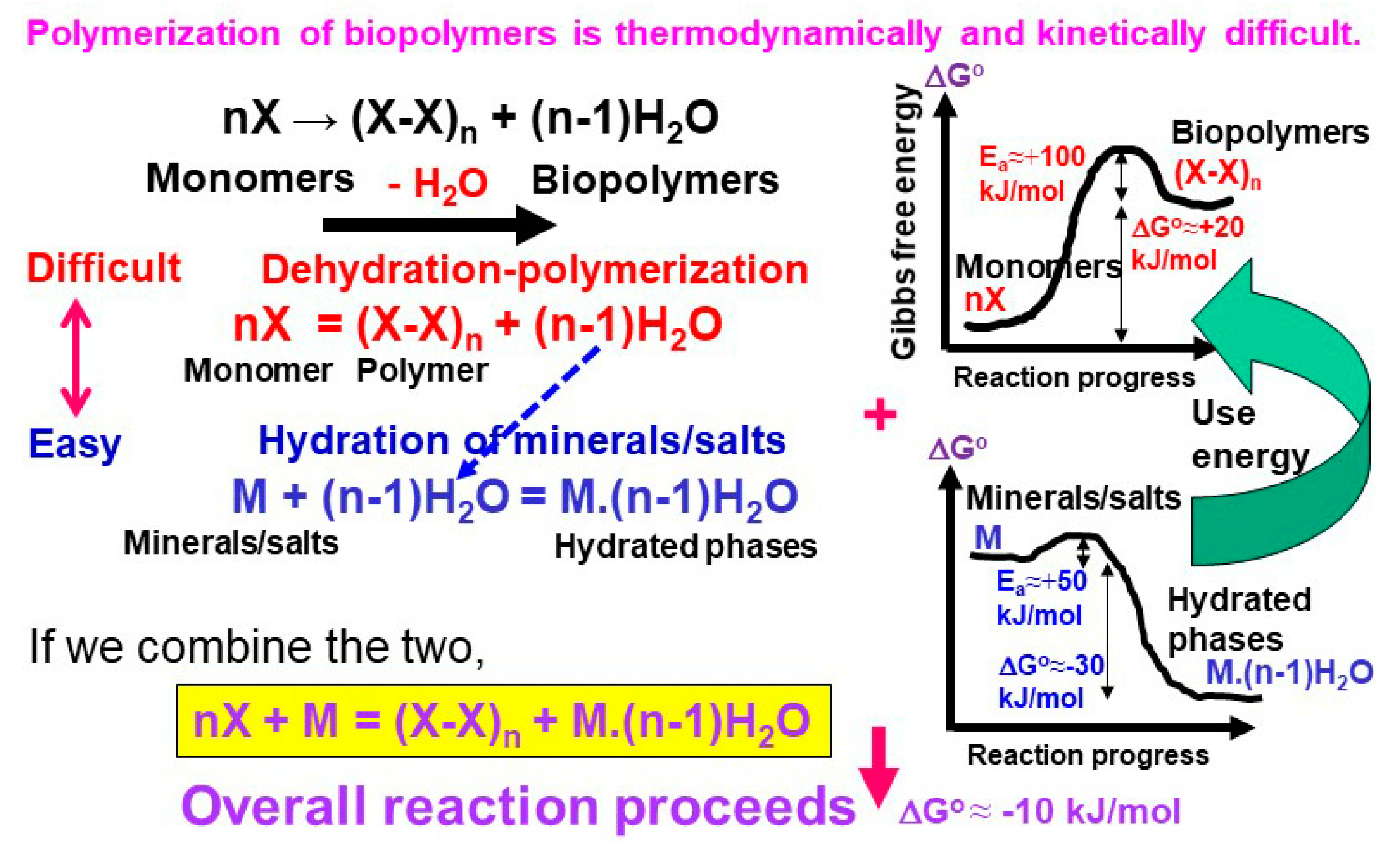{kind=link}
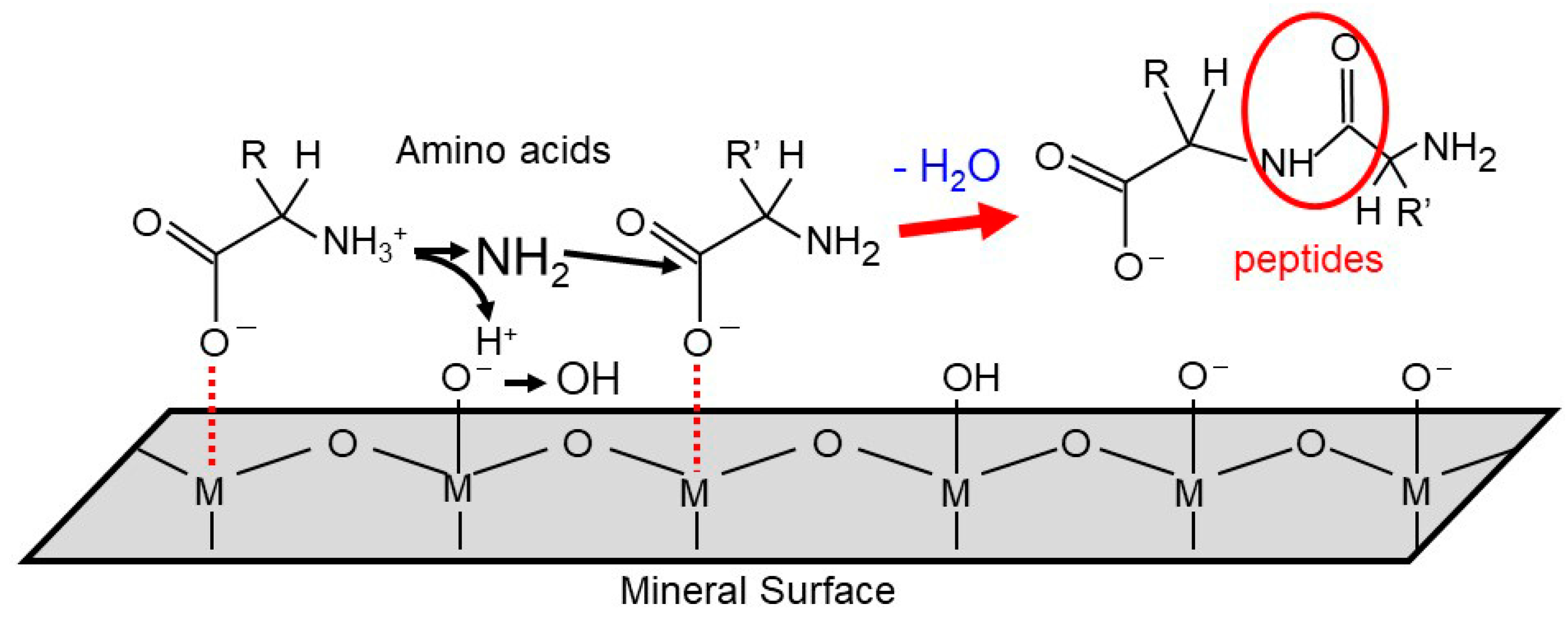{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Origins of the Building Blocks of Life
2.1. Organics in the Cosmic Environments and Their Delivery to Earth
2.2. Required Conditions for Prebiotic Chemistry
2.3. Roles of Mineral Surfaces
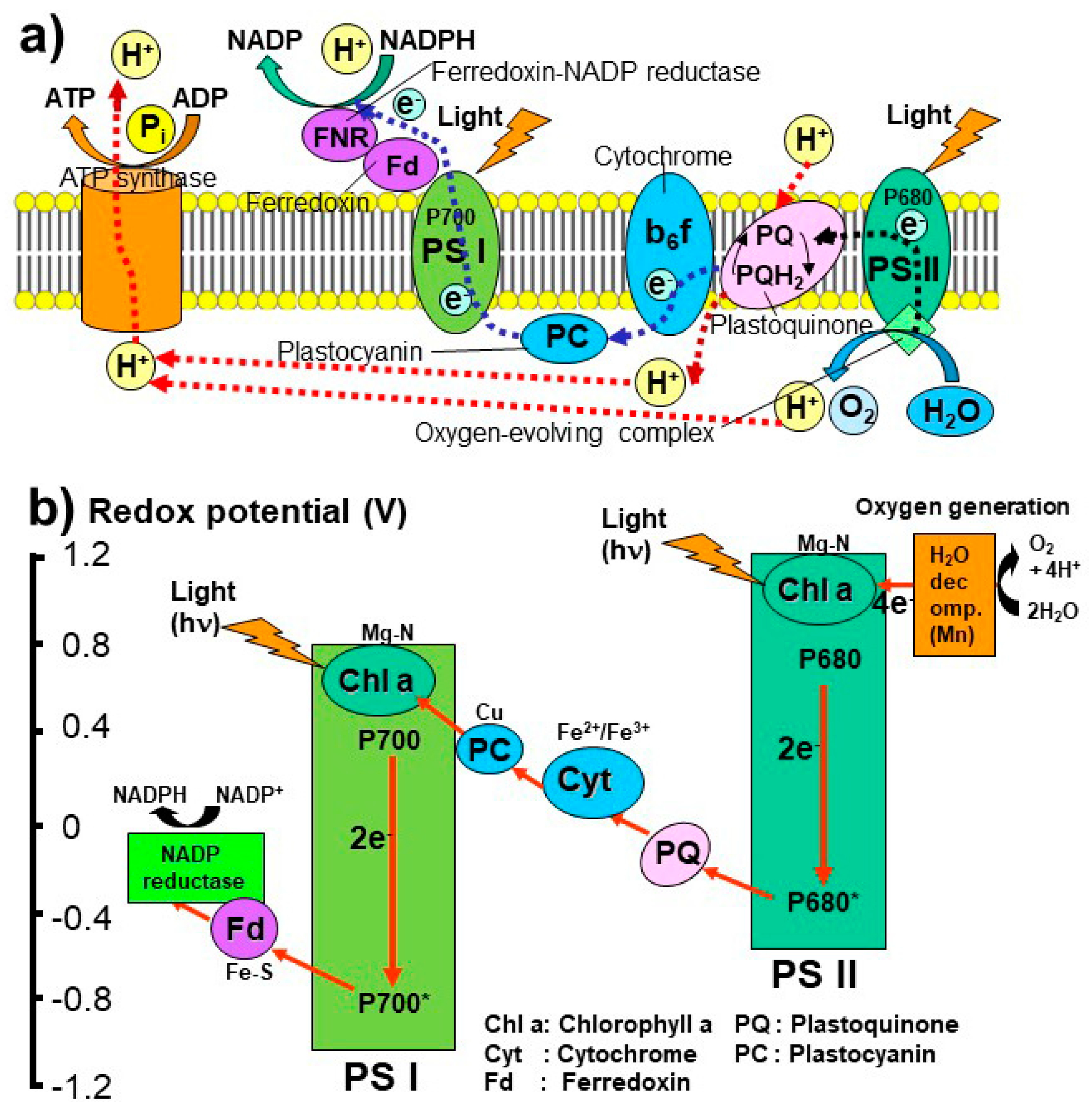
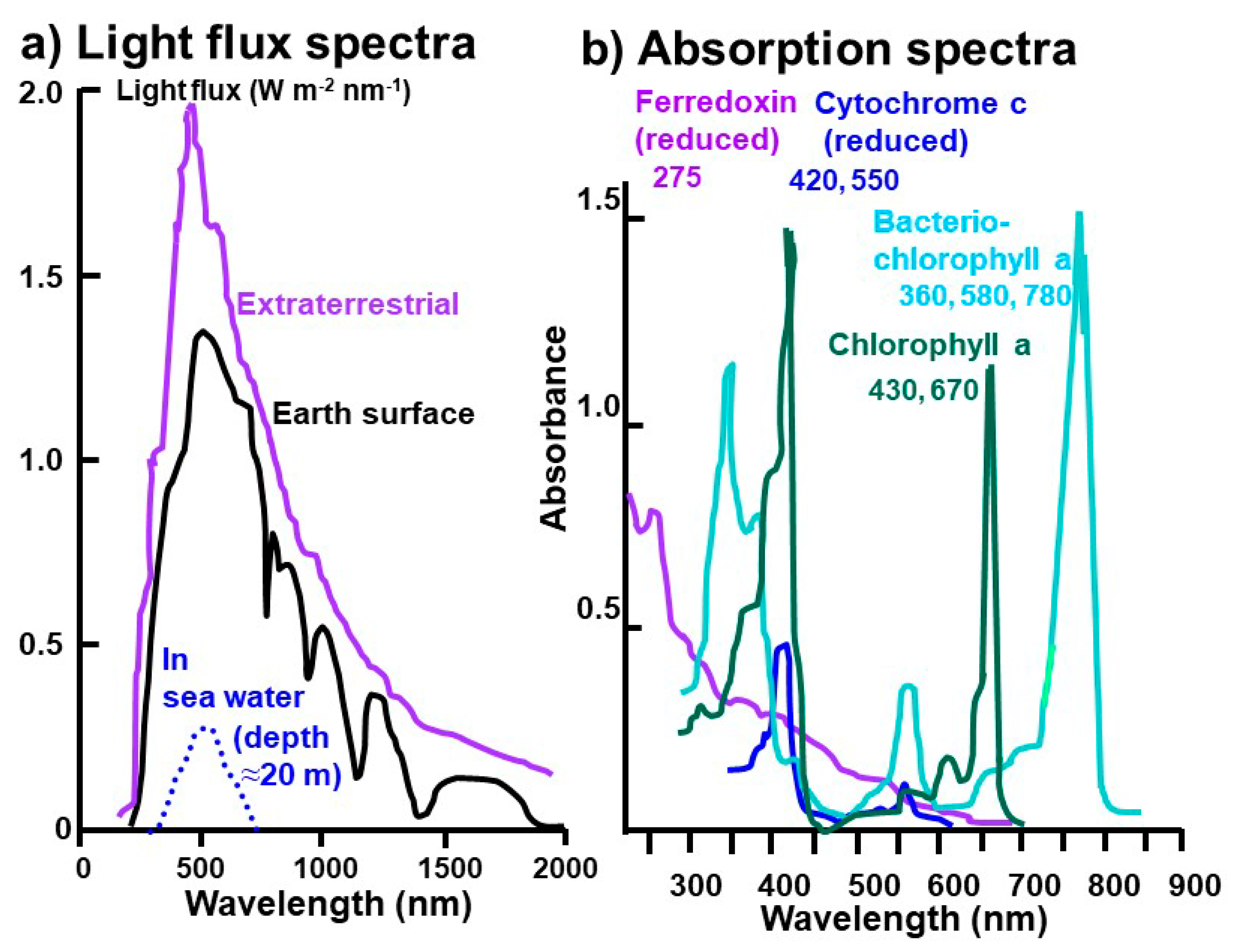
3. Origin and Evolution of Photosynthesis: A New Hypothesis
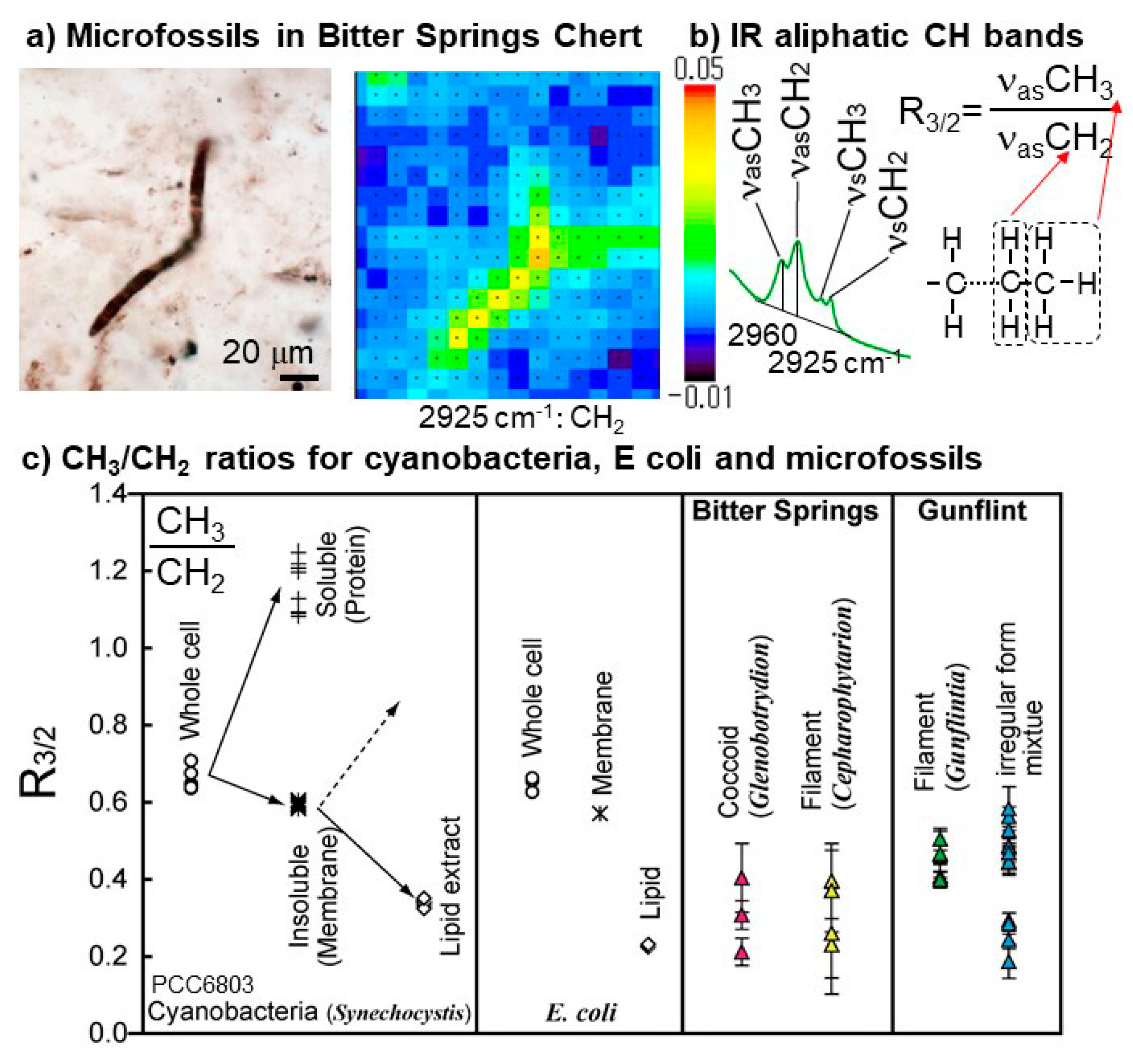
4. Fossilized Evidence of Life
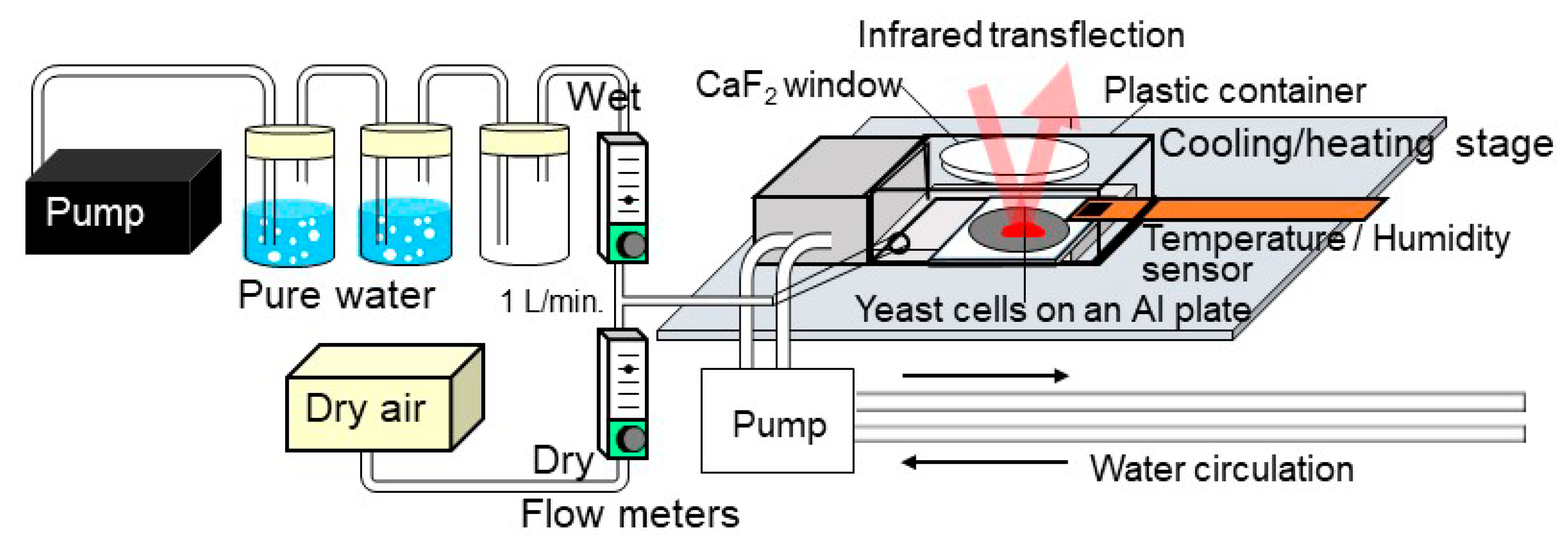
5. Roles of Water for Life through Studies on Anhydrobiosis
6. Summary and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zubay, G. Origins of Life on the Earth and in the Cosmos, 2nd ed.; Academic Press: San Diego, CA, USA, 2000; 564p. [Google Scholar]
- Brack, A. Introduction. In The Molecular Origin of Life. Assembling Pieces of the Puzzle; Brack, A., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 1–10. [Google Scholar]
- Maurel, M.C. La Naissance de la Vie et L’evolution Prebiotique a L’evolution Biologique; Diderot-Multimedia-EDL: Paris, France, 1999; 176p. [Google Scholar]
- Lahav, N. Biogenesis. Theories of Life’s Origin; Oxford University Press: Oxford, UK, 1999; 349p. [Google Scholar]
- Nakashima, S.; Maruyama, S.; Brack, A.; Windley, B.F. Geochemistry and the Origin of Life; Universal Academy Press: Tokyo, Japan, 2001; 355p. [Google Scholar]
- Kitadai, N.; Maruyama, S. Origins of building blocks of life: A review. Geosci. Front. 2018, 9, 1117–1153. [Google Scholar] [CrossRef]
- Maruyama, S.; Isozaki, Y.; Nakashima, S.; Windley, B.F. History of the Earth and life. In Geochemistry and the Origin of Life; Nakashima, S., Maruyama, S., Brack, A., Windley, B.F., Eds.; Universal Academy Press: Tokyo, Japan, 2001; pp. 285–328. [Google Scholar]
- Herbst, E.; van Dishoeck, E.F. Complex organic interstellar molecules. Annu. Rev. Astron. Astrophys. 2009, 47, 427–480. [Google Scholar] [CrossRef]
- Greenberg, J.M. Making a comet nucleus. Astron. Astrophys. 1998, 330, 375–380. [Google Scholar]
- Kasamatsu, T.; Kaneko, T.; Saito, T.; Kobayashi, K. Formation of organic compounds in simulated interstellar media with high energy particles. Bull. Chem. Soc. Jpn. 1997, 70, 1021–1026. [Google Scholar] [CrossRef]
- Bernstein, M.P.; Dworkin, J.P.; Sandford, S.A.; Cooper, G.W.; Allamandola, L.J. Racemic amino acids from the ultraviolet photolysis of interstellar ice analogues. Nature 2002, 416, 401–403. [Google Scholar] [CrossRef] [PubMed]
- Muñoz Caro, G.; Meierhenrich, U.; Schutte, W.; Barbier, B.; Arcones Segovia, A.; Rosenbauer, H.; Thiemann, W.-P.; Brack, A.; Greenberg, J. Amino acids from ultraviolet irradiation of interstellar ice analogues. Nature 2002, 416, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Zolensky, M.E.; Tomita, S.; Nakashima, S.; Tomeoka, K. Hollow organic globules in the Tagish Lake meteorite as possible products of primitive organic reactions. Int. J. Astrobiol. 2002, 1, 179–189. [Google Scholar] [CrossRef]
- Nakamura-Messenger, K.; Messenger, S.; Keller, L.P.; Clemett, S.J.; Zolensky, M.E. Organic globules in the tagish lake meteorite: Remnants of the protosolar disk. Science 2006, 314, 1439–1442. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, G.J.; Davis, A.M.; Zinner, E.K. The distribution of Al-26 in the early solar-system—A reappraisal. Meteoritics 1995, 30, 365–386. [Google Scholar] [CrossRef]
- Zolensky, M.E.; Bourcier, W.L.; Gooding, J.L. Aqueous alteration on the hydrous asteroids—Results of EQ3/6 computer-simulations. Icarus 1989, 78, 411–425. [Google Scholar] [CrossRef]
- Brearley, A.J. The action of water. In Meteorites and the Early Solar System II; Lauretta, D.S., McSween, J.H.Y., Eds.; University of Arizona Press: Tucson, AZ, USA, 2006; pp. 587–624. [Google Scholar]
- Fujiya, W.; Sugiura, N.; Hotta, H.; Ichimura, K.; Sano, Y. Evidence for the late formation of hydrous asteroids from young meteoritic carbonates. Nat. Commun. 2012, 3, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huss, G.R.; Rubin, A.E.; Grossman, J.N. Thermal metamorphism in chondrites. In Meteorites and the Early Solar System II; Lauretta, D.S., McSween, J.H.Y., Eds.; University of Arizona Press: Tucson, AZ, USA, 2006; pp. 567–586. [Google Scholar]
- Cooper, G.; Kimmich, N.; Belisle, W.; Sarinana, J.; Brabham, K.; Garrel, L. Carbonaceous meteorites as a source of sugar-related organic compounds for the early Earth. Nature 2001, 414, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Callahan, M.P.; Smith, K.E.; Cleaves, H.J., II; Ruzicka, J.; Stern, J.C.; Glavin, D.P.; House, C.H.; Dworkin, J.P. Carbonaceous meteorites contain a wide range of extraterrestrial nucleobases. Proc. Natl. Acad. Sci. USA 2011, 108, 13995–13998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, Z.; Botta, O.; Fogel, M.L.; Sephton, M.A.; Glavin, D.P.; Watson, J.S.; Dworkin, J.P.; Schwartz, A.W.; Ehrenfreund, P. Extraterrestrial nucleobases in the Murchison meteorite. Earth Planet. Sci. Lett. 2008, 270, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Elsila, J.E.; Aponte, J.C.; Blackmond, D.G.; Burton, A.S.; Dworkin, J.P.; Glavin, D.P. Meteoritic amino acids: Diversity in compositions reflects parent body histories. ACS Cent. Sci. 2016, 2, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Sandford, S.A.; Bera, P.P.; Lee, T.J.; Materese, C.K.; Nuevo, M. Photosynthesis and photo-stability of nucleic acids in prebiotic extraterrestrial environments. In Photoinduced Phenomena in Nucleic Acids II: DNA Fragments and Phenomenological Aspects; Barbatti, M., Borin, A.C., Ullrich, S., Eds.; Springer: Berlin, Germany, 2015; Volume 356, pp. 123–164. [Google Scholar]
- Greenberg, J.M.; Li, A.; Mendozagomez, C.X.; Schutte, W.A.; Gerakines, P.A.; Degroot, M. Approaching the interstellar grain organic refractory component. Astrophys. J. 1995, 455, L177–L180. [Google Scholar] [CrossRef]
- Nuth, J.A., III; Johnson, N.M.; Manning, S. A self-perpetuating catalyst for the production of complex organic molecules in protostellar nebulae. Astrophys. J. Lett. 2008, 673, L225–L228. [Google Scholar] [CrossRef]
- Pizzarello, S. Catalytic syntheses of amino acids and their significance for nebular and planetary chemistry. Meteorit. Planet. Sci. 2012, 47, 1291–1296. [Google Scholar] [CrossRef] [Green Version]
- Pizzarello, S.; Cooper, G.W.; Flynn, G.J. The nature and distribution of the organic material in carbonaceous chondrites and interplanetary dust particles. In Meteorites and the Early Solar System II; Lauretta, D.S., McSween, J.H.Y., Eds.; University of Arizona Press: Tucson, AZ, USA, 2006; pp. 625–651. [Google Scholar]
- Cody, G.D.; Heying, E.; Alexander, C.M.O.; Nittler, L.R.; Kilcoyne, A.L.D.; Sandford, S.A.; Stroud, R.M. Establishing a molecular relationship between chondritic and cometary organic solids. Proc. Natl. Acad. Sci. USA 2011, 108, 19171–19176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butlerow, A. Formation synthétique d’une substance sucrée. C. R. Acad. Sci. 1861, 53, 145–147. [Google Scholar]
- Kebukawa, Y.; Kilcoyne, A.L.D.; Cody, G.D. Exploring the potential formation of organic solids in chondrites and comets through polymerization of interstellar formaldehyde. Astrophys. J. 2013, 771, 19. [Google Scholar] [CrossRef]
- Kebukawa, Y.; Cody, G.D. A kinetic study of the formation of organic solids from formaldehyde: Implications for the origin of extraterrestrial organic solids in primitive solar system objects. Icarus 2015, 248, 412–423. [Google Scholar] [CrossRef]
- Kebukawa, Y.; Chan, Q.H.S.; Tachibana, S.; Kobayashi, K.; Zolensky, M.E. One-pot synthesis of amino acid precursors with insoluble organic matter in planetesimals with aqueous activity. Sci. Adv. 2017, 3, e1602093. [Google Scholar] [CrossRef] [PubMed]
- Kebukawa, Y.; Nakashima, S.; Zolensky, M.E. Kinetics of organic matter degradation in the Murchison meteorite for the evaluation of parentbody temperature history. Meteorit. Planet. Sci. 2010, 45, 99–113. [Google Scholar] [CrossRef]
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.H.; Percivalle, C.; Ritson, D.J.; Duffy, C.D.; Sutherland, J.D. Common origins of RNA, protein and lipids precursors in a cyanosulfidic protometabolism. Nat. Chem. 2015, 7, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. The physiology and habitat of the last universal common ancestor. Nat. Microbiol. 2016, 1, 16116. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, T.; Ishida, A.; Hori, M.; Igisu, M.; Koike, M.; Méjean, P.; Takahata, N.; Sano, Y.; Komiya, T. Early trace of life from 3.95 Ga sedimentary rocks in Labrador, Canada. Nature 2017, 549, 516–518. [Google Scholar] [CrossRef] [PubMed]
- Nunoura, T.; Chikaraishi, Y.; Izaki, R.; Suwa, T.; Sato, T.; Harada, T.; Mori, K.; Kato, Y.; Miyazaki, M.; Shimamura, S.; et al. A primordial and reversible TCA cycle in a facultatively chemolithoautotrophic thermophile. Science 2018, 359, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Hall, A.J.; Martin, W. Serpentinization as a source of energy at the origin of life. Geobiology 2010, 8, 355–371. [Google Scholar] [CrossRef] [PubMed]
- McCollom, T.M. Abiotic methane formation during experimental serpentinization of olivine. Proc. Natl. Acad. Sci. USA 2016, 113, 13965–13970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Nakamura, R.; Kasaya, T.; Kumagai, H.; Suzuki, K.; Takai, K. Spontaneous and widespread electricity generation in natural deep-sea hydrothermal fields. Angew. Chem. Int. Ed. 2017, 56, 5725–5728. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, R.; Takashima, T.; Kato, S.; Takai, K.; Yamamoto, M.; Hashimoto, K. Electrical current generation across a black smoker chimney. Angew. Chem. Int. Ed. 2010, 49, 7692–7694. [Google Scholar] [CrossRef] [PubMed]
- Ang, R.; Khan, A.U.; Tsujii, N.; Takai, K.; Nakamura, R.; Mori, T. Thermoelectricity generation and electron–magnon scattering in a natural chalcopyrite mineral from a deep-sea hydrothermal vent. Angew. Chem. Int. Ed. 2015, 54, 12909–12913. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Nakamura, R.; Oguri, K.; Kawagucci, S.; Suzuki, K.; Hashimoto, K.; Takai, K. Generation of electricity and illumination by an environmental fuel cell in deep-sea hydrothermal vents. Angew. Chem. Int. Ed. 2013, 52, 10758–10761. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.J.; Hall, A.J. The emergence of life from monosulphide bubbles at a submarine hydrothermal redox and pH front. J. Geol. Soc. Lond. 1997, 154, 377–402. [Google Scholar] [CrossRef]
- Maslennikov, V.V.; Maslennikova, S.P.; Large, R.R.; Danyushevsky, L.V.; Herrington, R.J.; Ayupova, N.R.; Zaykov, V.V.; Lein, A.Y.; Tseluyko, A.S.; Melekestseva, I.Y.; et al. Chimneys in Paleozoic massive sulfide mounds of the Urals VMS deposits: Mineral and trace element comparison with modern black, grey, white and clear smokers. Ore Geol. Rev. 2017, 85, 64–106. [Google Scholar] [CrossRef]
- Russell, M.J.; Barge, L.M.; Bhartia, R.; Bocanegra, D.; Bracheer, P.J.; Branscomb, E.; Kidd, R.; McGlynn, S.; Meier, D.H.; Nitschke, W.; et al. The drive to life on wet and icy worlds. Astrobiology 2014, 14, 308–343. [Google Scholar] [CrossRef] [PubMed]
- Kitadai, N.; Nakamura, R.; Yamamoto, M.; Takai, K.; Li, Y.; Yamaguchi, A.; Gilbert, A.; Ueno, Y.; Yoshida, N.; Oono, Y. Geoelectrochemical CO production: Implications of the autotrophic origin of life. Sci. Adv. 2018, 4, eaao7265. [Google Scholar] [CrossRef] [PubMed]
- Huber, C.; Wächtershäuser, G. Activated acetic acid by carbon fixation on (Fe,Ni)S under primordial conditions. Science 1997, 276, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Huber, C.; Wächtershäuser, G. Peptides by activation of amino acids with CO on (Ni,Fe)S surfaces: Implications for the origin of life. Science 1998, 281, 670–672. [Google Scholar] [CrossRef] [PubMed]
- Huber, C.; Wächtershäuser, G. α-hydroxy and α-amino acids under possible Hadean, volcanic origin-of-life conditions. Science 2006, 314, 630–632. [Google Scholar] [CrossRef] [PubMed]
- Yamagushi, A.; Yamamoto, M.; Takai, K.; Ishii, T.; Hashimoto, K.; Nakamura, R. Electrochemical CO2 reduction by Ni-containing iron sulfides; How is CO2 electrochemically reduced at bisulfide-bearing deep-sea hydrothermal precipitates? Electrochim. Acta 2014, 141, 311–318. [Google Scholar] [CrossRef]
- Morrill, P.L.; Kuenen, J.G.; Johnson, O.J.; Suzuki, S.; Rietze, A.; Sessions, A.L.; Fogel, M.L.; Nealson, K.H. Geochemistry and geobiology of a present-day serpentinization site in California: The Cedars. Geochim. Cosmochim. Acta 2013, 109, 222–240. [Google Scholar] [CrossRef] [Green Version]
- Mulkidjaniana, A.Y.; Bychkovc, A.Y.; Dibrova, D.V.; Galperine, M.Y.; Koonin, E.V. Origin of first cells at terrestrial, anoxic geothermal fields. Proc. Natl. Acad. Sci. USA 2012, 109, E821–E830. [Google Scholar] [CrossRef] [PubMed]
- Fenus, M.; Michalčíková, R.; Shestivská, V.; Sponer, J.; Sponer, J.E.; Civiš, S. High-energy chemistry of formamide: A simpler way for nucleobase formation. J. Phys. Chem. A 2014, 118, 719–736. [Google Scholar]
- Saladino, R.; Carotaa, E.; Bottaa, G.; Kapralovb, M.; Timoshenkob, G.N.; Rozanovb, A.Y.; Krasavinb, E.; Di Mauro, E. Meteorite-catalyzed syntheses of nucleosides and of other prebiotic compounds from formamide under proton irradiation. Proc. Natl. Acad. Sci. USA 2015, 112, E2746–E2755. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Botta, G.; Mattia Bizzarri, B.; Di Mauro, E.; Garcia Ruiz, J.M. A Global scale scenario for prebiotic chemistry: Silica-based self-assembled mineral structures and formamide. Biochemistry 2016, 55, 2806–2811. [Google Scholar] [CrossRef] [PubMed]
- Sponer, J.E.; Sponer, J.; Novkov, O.; Brabec, V.; Sedo, O.; Zdrhal, Z.; Costanzo, G.; Pino, S.; Saladino, R.; Di Mauro, E. Emergence of the first catalytic oligonucleotides in a formamide-based origin scenario. Chem. Eur. J. 2016, 22, 3572–3586. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A. Paradoxes in the origin of life. Orig. Life Evol. Biosph. 2014, 44, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.; Barbier, B.; Boillot, F.; Chabin, A. Extraterrestrial organic molecules and the origin of life. In Geochemistry and the Origin of Life; Nakashima, S., Maruyama, S., Brack, A., Windley, B.F., Eds.; Universal Academy Press: Tokyo, Japan, 2001; pp. 17–38. [Google Scholar]
- Maurel, M.C. Functional capabilities of biomolecules at the beginnings of life. —RNA selectivity and adaptability—. In Geochemistry and the Origin of Life; Nakashima, S., Maruyama, S., Brack, A., Windley, B.F., Eds.; Universal Academy Press: Tokyo, Japan, 2001; pp. 107–118. [Google Scholar]
- Hazen, R.M.; Filley, T.R.; Goodfriend, G.A. Selective adsorption of L- and D-amino acids on calcite: Implications for biochemical homochirality. Proc. Natl. Acad. Sci. USA 2001, 98, 5487–5490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, J.D. The physical basis of life. Proc. Phys. Soc. 1949, 62, 537–558. [Google Scholar] [CrossRef]
- Bujdak, J.; Faybikova, K.; Eder, A.; Yongyai, Y.; Rode, B. Peptide-chain elongation—A possible role of montmorillonite in prebiotic synthesis of protein precursors. Orig. Life Evol. Biosph. 1995, 25, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P.; Hill, A.J.; Liu, R.; Orgel, L. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 1996, 381, 59–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bujdak, J.; Rode, B.M. The effect of smectite composition on the catalysis of peptide bond formation. J. Mol. Evol. 1996, 43, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Bujdak, J.; Rode, B. Silica, alumina, and clay-catalyzed alanine peptide bond formation. J. Mol. Evol. 1997, 45, 457–466. [Google Scholar] [CrossRef]
- Bujdak, J.; Rode, B. Silica, alumina and clay catalyzed peptide bond formation: Enhanced efficiency of alumina catalyst. Orig. Life Evol. Biosph. 1999, 29, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P. Catalyzed RNA synthesis for the RNA world. In The Molecular Origin of Life. Assembling Pieces of the Puzzle; Brack, A., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 255–268. [Google Scholar]
- Orgel, L.E. Polymerization on the rocks: Theoretical introduction. Orig. Life Evol. Biosph. 1998, 28, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.R.; Böhler, C.; Orgel, L. Polymerization on the rocks: Negatively charged α-amino acids. Orig. Life Evol. Biosph. 1998, 28, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Orgel, L. Polymerization on the rocks: β-amino acids and arginine. Orig. Life Evol. Biosph. 1998, 28, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Rode, B.M.; Son, H.L.; Suwannachot, Y. The combination of self-induced peptide formation reaction and clay catalysis: A way to higher peptides under primitive Earth conditions. Orig. Life Evol. Biosph. 1999, 29, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Cairns-Smith, A.G. Genetic Takeover and the Mineral Origin of Life; Cambridge University Press: Cambridge, UK, 1982; 488p. [Google Scholar]
- Wächtershäuser, G. Before enzymes and templates: Theory of surface metabolism. Microbiol. Rev. 1988, 52, 452–484. [Google Scholar] [PubMed]
- Wächtershäuser, G. Origin of life in an iron-sulfur world. In The Molecular Origin of Life. Assembling Pieces of the Puzzle; Brack, A., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 206–218. [Google Scholar]
- Kitadai, N. Thermodynamic prediction of glycine polymerization as a function of temperature and pH consistent with experimentally obtained results. J. Mol. Evol. 2014, 78, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, S.; Shiota, D. Organic-inorganic interactions and the origin and evolution of life. In Geochemistry and the Origin of Life; Nakashima, S., Maruyama, S., Brack, A., Windley, B.F., Eds.; Universal Academy Press: Tokyo, Japan, 2001; pp. 135–177. [Google Scholar]
- Kitadai, N.; Yokoyama, T.; Nakashima, S. Hydration-dehydration interactions between glycine and anhydrous salts: Implications for chemical evolution of life. Geochim. Cosmochim. Acta 2011, 75, 6285–6299. [Google Scholar] [CrossRef]
- Kitadai, N.; Oonishi, H.; Umemoto, K.; Usui, T.; Fukushi, K.; Nakashima, S. Glycine polymerization on oxide minerals. Orig. Life Evol. Biosph. 2017, 47, 123–143. [Google Scholar] [CrossRef] [PubMed]
- Sakata, K.; Kitadai, N.; Yokoyama, T. Effects of pH and temperature on dimerization rate of glycine: Evaluation of favorable environmental conditions for chemical evolution of life. Geochim. Cosmochim. Acta 2010, 74, 6841–6851. [Google Scholar] [CrossRef]
- Kitadai, N.; Nishiuchi, K.; Nishii, A.; Fukushi, K. Amorphous silica-promoted lysine dimerization: A thermodynamic prediction. Orig. Life Evol. Biosph. 2018, 48, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Sverjensky, D.A. Prediction of surface charge on oxides in salt solutions: Revisions for 1:1 (M+L−) electrolytes. Geochim. Cosmochim. Acta 2005, 69, 225–257. [Google Scholar] [CrossRef]
- Kitadai, N.; Nishiuchi, K.; Tanaka, M. A comprehensive predictive model for sulfate adsorption on oxide minerals. Geochim. Cosmochim. Acta 2018, 238, 150–168. [Google Scholar] [CrossRef]
- Kitadai, N.; Yokoyama, T.; Nakashima, S. ATR-IR spectroscopic study of L-lysine adsorption on amorphous silica. J. Colloid Interface Sci. 2009, 329, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Kitadai, N.; Yokoyama, T.; Nakashima, S. In situ ATR-IR investigation of L-lysine adsorption on montmorillonite. J. Colloid Interface Sci. 2009, 338, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Akouche, M.; Jaber, M.; Zins, E.-L.; Maurel, M.-C.; Lambert, J.-F.; Georgelin, T. Thermal behavior of d-ribose adsorbed on silica: Effect of inorganic salt coadsorption and significance for prebiotic chemistry. Chem. Eur. J. 2016, 22, 15834–15846. [Google Scholar] [CrossRef] [PubMed]
- Akouche, M.; Jaber, M.; Maurel, M.-C.; Lambert, J.-F.; Georgelin, T. Phosphoribosyl pyrophosphate: A molecular vestige of the origin of life on minerals. Angew. Chem. Int. Ed. 2017, 56, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Villa, A.; Saitta, M.; Georgelin, T.; Lambert, J.F.; Guyot, F.; Maurel, M.C.; Pietrucci, F. Synthesis of RNA nucleotides in plausible prebiotic conditions from ab initio computer simulations. J. Phys. Chem. Lett. 2018, 9, 4981–4987. [Google Scholar] [CrossRef] [PubMed]
- Wächtershäuser, G. 1990 Evolution of the first metabolic cycles. Proc. Natl. Acad. Sci. USA 1990, 87, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Aono, M.; Kitadai, N.; Oono, Y. A principled approach to the origin problem. Orig. Life Evol. Biosph. 2015, 45, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, R.E. Early evolution of photosynthesis. Plant Physiol. 2010, 154, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Hohmann-Marriott, M.F.; Blankenship, R.E. Evolution of photosynthesis. Annu. Rev. Plant Biol. 2011, 62, 515–548. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis, 2nd ed.; John Wiley & Sons: Oxford, UK, 2014; 296p. [Google Scholar]
- Martin, W.F.; Bryant, D.A.; Beatty, J.T. A physiological perspective on the origin and evolution of photosynthesis. FEMS Microbiol. Rev. 2018, 42, 205–231. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.C.; Prezelin, B.B.; Baker, K.S.; Bidigare, R.R.; Boucher, N.P.; Coley, T.; Karentz, D.; MacIntyre, S.; Matlick, H.A.; Menzies, D.; et al. Ozone depletion: Ultraviolet radiation and phytoplankton biology in Antarctic waters. Science 1992, 255, 952–959. [Google Scholar] [CrossRef] [PubMed]
- Seinfeld, J.H.; Pandis, S.N. Atmospheric Chemistry and Physics—From Air Pollution to Climate Change, 2nd ed.; John Wiley and Sons: Hoboken, NJ, USA, 2006; 1203p. [Google Scholar]
- Wang, S.; Huang, H.; Kahnt, J.; Thauer, R.K. A Reversible Electron-Bifurcating Ferredoxin- and NAD-Dependent [FeFe]-Hydrogenase (HydABC) in Moorella thermoacetica. J. Bacteriol. 2013, 195, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Granick, S. Evolution of heme and chlorophyll. In Evolving Genes and Proteins; Bryson, V., Vogel, H.J., Eds.; Academic Press: New York, NY, USA, 1965; pp. 67–88. [Google Scholar]
- Olson, J.M.; Pierson, B.K. Evolution of reaction centers in photosynthetic prokaryotes. Int. Rev. Cytol. 1987, 108, 209–248. [Google Scholar] [PubMed]
- Mauzerall, D. Light, iron, Sam Granick and the origin of life. Photosynth. Res. 1992, 33, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.M. Early evolution of chlorophyll-based photosystems. Chemtracts Biochem. Mol. Biol. 1999, 12, 468–482. [Google Scholar]
- Olson, J.M. Photosynthesis in the Archean era. Photosynth. Res. 2006, 88, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Kritsky, M.S.; Telegina, T.A.; Vechtomova, Y.L.; Buglak, A.A. Why flavins are not competitors of chlorophyll in the evolution of biological converters of solar energy. Int. J. Mol. Sci. 2013, 14, 575–593. [Google Scholar] [CrossRef] [PubMed]
- Dodd, M.S.; Papineau, D.; Grenne, T.; Slack, J.F.; Rittner, M.; Pirajno, F.; O’Neil, J.; Little, C.T.S. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nature 2017, 543, 60–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopf, J.W. Microfossils of the early Archean Apex chert: New evidence of the antiquity of life. Science 1993, 260, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Brasier, M.D.; Green, O.R.; Jephocoat, A.P.; Kleppe, A.K.; Van Kranendonk, M.J.; Lindsay, J.F.; Steele, A.; Grassineau, N.V. Questioning the evidence for Earth’s oldest fossils. Nature 2002, 416, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.M. Global methanotrophy at the Archean-Proterozoic transition. In Early life on Earth; Schopf, J.W., Ed.; Columbia University Press: New York, NY, USA, 1994. [Google Scholar]
- Brocks, J.J.; Logan, G.A.; Buick, R.; Summons, R.E. Archean molecular fossils and the early rise of eukaryotes. Science 1999, 285, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Rasmmusen, B.; Fletcher, I.R.; Brocks, J.J.; Kilburn, M.R. Reassessing the first appearance of eukaryotes and cyanobacteria. Nature 2008, 455, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- House, C.H.; Schopf, J.W.; McKeegan, K.D.; Coath, C.D.; Harrison, T.M.; Stetter, K.O. Carbon isotopic composition of individual Precambrian microfossils. Geology 2000, 28, 707–710. [Google Scholar] [CrossRef]
- House, C.H.; Oehler, D.Z.; Sugitani, K.; Mimura, K. Carbon isotopic analyses of ca. 3.0 Ga microstructures imply planktonic autotrophs inhabited Earth’s early oceans. Geology 2013, 41, 651–654. [Google Scholar] [CrossRef]
- Kudryavtsev, A.B.; Schopf, J.W.; Agresti, D.G.; Wdowiak, T.J. In situ laser-Raman imagery of Precambrian microscopic fossils. Proc. Natl. Acad. Sci. USA 2001, 98, 823–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, Y.; Isozaki, Y.; Yurimoto, H.; Maruyama, S. Carbon isotopic signatures of individual Archean microfossils (?) from Western Australia. Int. Geol. Rev. 2001, 43, 196–212. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B.; Agresti, D.G.; Wdowiak, T.J.; Czaja, A.D. Laser-Raman imagery of Earth’s earliest fossils. Nature 2002, 416, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Schopf, J.W.; Kudryavtsev, A.B.; Agresti, D.G.; Czaja, A.D.; Wdowiak, T.J. Raman imagery: A new approach to assess the geochemical maturity and biogenicity of permineralized Precambrian fossils. Astrobiology 2005, 5, 333–371. [Google Scholar] [CrossRef] [PubMed]
- Schopf, J.W.; Kitajima, K.; Spicuzza, M.J.; Kudryavtsev, A.B.; Valley, J.W. SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions. Proc. Natl. Acad. Sci. USA 2018, 115, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Oehler, D.Z.; Robert, F.; Mostefaoui, S.; Meibom, A.; Selo, M.; McKay, D.S. Chemical mapping of Proterozoic organic matter at submicron spatial resolution. Astrobiology 2006, 6, 838–850. [Google Scholar] [CrossRef] [PubMed]
- Igisu, M.; Nakashima, S.; Ueno, Y.; Awramik, S.M.; Maruyama, S. In situ infrared microspectroscopy of ~850 million-year-old prokaryotic fossils. Appl. Spectrosc. 2006, 60, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Igisu, M.; Ueno, Y.; Shimojima, M.; Nakashima, S.; Awramik, S.M.; Ohta, H.; Maruyama, S. Micro-FTIR signature of bacterial lipids in Proterozoic microfossils. Precambrian Res. 2009, 173, 19–26. [Google Scholar] [CrossRef]
- Van Zuilen, M.A.; Chaussidon, M.; Rollion-Bard, C.; Marty, B. Carbonaceous cherts of the Barberton Greenstone Belt, South Africa: Isotopic, chemical and structural characteristics of individual microstructures. Geochim. Cosmochim. Acta 2007, 71, 655–669. [Google Scholar] [CrossRef]
- Wacey, D.; Kilburn, M.R.; Saunders, M.; Cliff, J.; Brasier, M.D. Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia. Nat. Geosci. 2011, 4, 698–702. [Google Scholar] [CrossRef]
- Wacey, D.; Battison, L.; Garwood, R.J.; Hickman-Lewis, K.; Brasier, M.D. Advanced analytical techniques for studying the morphology and chemistry of Proterozoic microfossils. Geol. Soc. Lond. Spec. Publ. 2016, 448, 81–104. [Google Scholar] [CrossRef] [Green Version]
- Lepot, K.; Willford, K.H.; Ushikubo, T.; Sugitani, K.; Mimura, K.; Spicuzza, M.J.; Valley, J.W. Texture-specific isotopic compositions in 3.4 Gyr old organic matter support selective preservation in cell like structures. Geochim. Cosmochim. Acta 2013, 112, 66–86. [Google Scholar] [CrossRef]
- Williford, K.H.; Ushikubo, T.; Schopf, J.W.; Lepot, K.; Kitajima, K.; Valley, J.W. Preservation and detection of microstructural and taxonomic correlations in the carbon isotopic compositions of individual Precambrian microfossils. Geochim. Cosmochim. Acta 2013, 104, 165–182. [Google Scholar] [CrossRef]
- Alleon, J.; Bernard, S.; Le Guillou, C.; Marin-Carbonne, J.; Pont, S.; Beyssac, O.; McKeegan, D.D.; Robert, F. Molecular preservation of 1.88 Ga Gunflint organic microfossils as a function of temperature and mineralogy. Nat. Commun. 2016, 7, 11977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, Y.; Isozaki, Y.; McNamara, K.J. Coccoid like microstructures in a 3.0 Ga chert from Western Australia. Int. Geol. Rev. 2006, 48, 78–88. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B. Confocal laser scanning microscopy and Raman imagery of ancient microscopic fossils. Precambrian Res. 2009, 173, 39–49. [Google Scholar] [CrossRef]
- Chen, J.Y.; Schopf, J.W.; Bottjer, D.J.; Zhang, C.Y.; Kudryavtsev, A.B.; Tripathi, A.B.; Wang, X.Q.; Yang, Y.H.; Gao, X.; Yang, Y. Raman spectra of a Lower Cambrian ctenophore embryo from southwestern Shaanxi, China. Proc. Natl. Acad. Sci. USA 2007, 104, 6289–6292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugitani, K.; Grey, K.; Allwood, A.; Nagaoka, T.; Mimura, K.; Minami, M.; Marshall, C.P.; Van Kranendonk, M.J.; Walter, M.R. Diverse microstructures from Archean chert from the Mount Gldsworthy‒Mount Grant area, Pilbara Craton, Western Australia: Microfossils, dubiofossils, or pseudofossils? Precambrian Res. 2007, 158, 228–262. [Google Scholar] [CrossRef]
- Qu, Y.; Engdahl, A.; Zhu, S.; Vajda, V.; McLoughlin, N. Ultrastructural heterogeneity of carbonaceous material in ancient cherts: Investigating biosignature origin and preservation. Astrobiology 2015, 15, 825–842. [Google Scholar] [CrossRef] [PubMed]
- Pasteris, J.D.; Wopenka, B. Necessary, but not sufficient: Raman identification of disordered carbon as a signature of ancient life. Astrobiology 2003, 3, 727–738. [Google Scholar] [CrossRef] [PubMed]
- Wopenka, B.; Pasteirs, J.D. Structural characterization of kerogens to granulite-facies graphite: Applicability of Raman microprobe spectroscopy. Am. Mineral. 1993, 78, 533–557. [Google Scholar]
- Lahfid, A.; Beyssac, O.; Diville, E.; Negro, F.; Chopin, C.; Goffe, B. Evolution of the Raman spectrum of carbonaceous material in low-grade metasediments of the Glarus Alps (Switzerland). Terra Nova 2010, 22, 354–360. [Google Scholar] [CrossRef]
- Lin, R.; Ritz, G.P. Reflectance FT-IR microspectroscopy of fossil algae contained in organic-rich shales. Appl. Spectrosc. 1993, 47, 265–271. [Google Scholar] [CrossRef]
- Qu, Y.; Wang, J.; Xiao, S.; Whitehouse, M.; Engdahl, A.; Wang, G.; McLoughlin, N. Carbonaceous biosignatures of diverse chemotrophic microbial communities from chert nodules of the Ediacaran Doushantuo Formation. Precambrian Res. 2017, 290, 184–196. [Google Scholar] [CrossRef] [Green Version]
- Igisu, M.; Komiya, T.; Kawashima, M.; Nakashima, S.; Ueno, Y.; Han, J.; Shu, D.; Li, Y.; Maruyama, S.; Takai, K. FTIR microspectroscopy of Ediacaran phosphatized microfossils from the Doushantuo Formation, Weng’an, South China. Gondwana Res. 2014, 25, 1120–1138. [Google Scholar] [CrossRef]
- De Gregorio, B.T.; Sharp, T.G. The structure and distribution of carbon in 3.5 Ga Apex chert: Implications for the biogenicity of earth’s oldest putative microfossils. Am. Mineral. 2006, 91, 784–789. [Google Scholar] [CrossRef]
- De Gregorio, B.T.; Sharp, T.G.; Flynn, G.J.; Wirick, S.; Hervig, R.L. Biogenic origin for earth’s oldest putative microfossils. Geology 2009, 37, 631–634. [Google Scholar] [CrossRef]
- Alleon, J.; Bernard, S.; Le Guillou, C.; Daval, D.; Skouri-Panet, F.; Pont, S.; Delbes, L.; Robert, F. Early entombment within silica minimizes the molecular degradation of microorganisms during advanced diagenesis. Chem. Geol. 2016, 437, 98–108. [Google Scholar] [CrossRef]
- Igisu, M.; Yokoyama, T.; Nakashima, S.; Ueno, Y.; Shimojima, M.; Ohta, H.; Maruyama, S. Changes of aliphatic C–H bonds in cyanobacteria during experimental thermal maturation in the presence or absence of silica as evaluated by FTIR microspectroscopy. Geobiology 2018, 16, 412–428. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Crowe, J.H.; Crowe, L.M.; Carpenter, J.E.; Petrelski, S.; Hoekstra, F.A.; de Araujo, P.; Panek, A.D. Anhydrobiosis: Cellular adaptation to extreme dehydration. In Handbook of Physiology, Section 13: Comparative Physiology; Oxford University Press: Oxford, UK, 1977; pp. 1445–1477. [Google Scholar]
- Cordone, L.; Cottone, G.; Cupane, A.; Emanuele, A.; Giuffrida, S.; Levantino, M. Proteins in saccharides matrices and the trehalose peculiarity: Biochemical and biophysical properties. Curr. Org. Chem. 2015, 19, 1684–1706. [Google Scholar] [CrossRef]
- Wolkers, W.F.; Bochicchio, A.; Selvaggi, G.; Hoekstra, F.A. Fourier transform infrared microspectroscopy detects changes in protein secondary structure associated with desiccation tolerance in developing maize embryos. Plant Physiol. 1998, 116, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Kudo, S.; Ogawa, H.; Yamakita, E.; Watanabe, S.; Suzuki, T.; Nakashima, S. Adsorption of water to collagen as studied by infrared (IR) micro-Spectroscopy combined with relative humidity control system and quartz crystal microbalance. Appl. Spectrosc. 2017, 71, 1621–1632. [Google Scholar] [CrossRef] [PubMed]
- Vladilo, G.; Hassanali, A. Hydrogen bonds and life in the universe. Life 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakashima, S.; Kebukawa, Y.; Kitadai, N.; Igisu, M.; Matsuoka, N. Geochemistry and the Origin of Life: From Extraterrestrial Processes, Chemical Evolution on Earth, Fossilized Life’s Records, to Natures of the Extant Life. Life 2018, 8, 39. https://0-doi-org.brum.beds.ac.uk/10.3390/life8040039
Nakashima S, Kebukawa Y, Kitadai N, Igisu M, Matsuoka N. Geochemistry and the Origin of Life: From Extraterrestrial Processes, Chemical Evolution on Earth, Fossilized Life’s Records, to Natures of the Extant Life. Life. 2018; 8(4):39. https://0-doi-org.brum.beds.ac.uk/10.3390/life8040039
Chicago/Turabian StyleNakashima, Satoru, Yoko Kebukawa, Norio Kitadai, Motoko Igisu, and Natsuki Matsuoka. 2018. "Geochemistry and the Origin of Life: From Extraterrestrial Processes, Chemical Evolution on Earth, Fossilized Life’s Records, to Natures of the Extant Life" Life 8, no. 4: 39. https://0-doi-org.brum.beds.ac.uk/10.3390/life8040039