MicroRNA in Gastric Cancer Development: Mechanisms and Biomarkers

 ,
,
Abstract
:1. Introduction
2. Features of the microRNA Interaction with Signaling Pathways in GC
3. MicroRNAs Associated with GC Metastasis and Signaling Pathways Regulated by Them
4. MicroRNAs Associated with the Development of GC Chemoresistance and Signaling Pathways Regulated by Them
5. MicroRNA as GC Prognostic and Diagnostic Markers
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bhat, S.A.; Majid, S.; Rehman, M.U. Scenario and future prospects of microRNAs in gastric cancer: A review. Iran. J. Basic Med. Sci. 2019. [Google Scholar] [CrossRef]
- Kim, S.; Bae, W.J.; Ahn, J.M.; Heo, J.H.; Kim, K.M.; Choi, K.W.; Sung, C.O.; Lee, D. MicroRNA signatures associated with lymph node metastasis in intramucosal gastric cancer. Mod. Pathol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Hu, G.F.; Shi, Y.F.; Xu, W. Research progress in microRNA-based therapy for gastric cancer. OncoTargets Ther. 2019, 12, 11393–11411. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Tang, X.; Liu, Y.; Zhu, J.; Liu, J. Induction/reversal of drug resistance in gastric cancer by non-coding RNAs (Review). Int. J. Oncol. 2019, 54, 1511–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.L.; Wang, T.; Zhang, K.H. MicroRNAs as potential biomarkers for diagnosis, therapy and prognosis of gastric cancer. OncoTargets Ther. 2018, 11, 3891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ESMO Guidelines Committee. Gastric Cancer Treatment Recommendations. Available online: https://www.esmo.org/guidelines/gastrointestinal-cancers/gastric-cancer/eupdate-gastric-cancer-treatment-recommendations2 (accessed on 8 October 2020).
- Jackstadt, R.; Hodder, M.C.; Sansom, O.J. WNT and β-Catenin in Cancer: Genes and Therapy. Annu. Rev. Cancer Biol. 2020, 4, 177–196. [Google Scholar] [CrossRef] [Green Version]
- Weng, J.; Li, S.; Lin, H.; Mei, H.; Liu, Y.; Xiao, C.; Zhu, Z.; Cai, W.; Ding, X.; Mi, Y.; et al. PCDHGA9 represses epithelial-mesenchymal transition and metastatic potential in gastric cancer cells by reducing β-catenin transcriptional activity. Cell Death Dis. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Jiang, N.; Dai, Q.; Su, X.; Fu, J.; Feng, X.; Peng, J. Role of PI3K/AKT Pathway in Cancer: The Framework of Malignant Behavior; Springer: Amsterdam, The Netherlands, 2020; Volume 47, ISBN 0123456789. [Google Scholar]
- Matsuoka, T.; Yashiro, M. The role of PI3K/Akt/mTOR signaling in gastric carcinoma. Cancers 2014, 6, 1441–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fresno Vara, J.Á.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. P13K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef]
- Chen, H.T.; Liu, H.; Mao, M.J.; Tan, Y.; Mo, X.Q.; Meng, X.J.; Cao, M.T.; Zhong, C.Y.; Liu, Y.; Shan, H.; et al. Crosstalk between autophagy and epithelial-mesenchymal transition and its application in cancer therapy. Mol. Cancer 2019, 18, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Deng, Z.; Wang, Z.; Wang, D.; Zhang, L.; Su, Q.; Lai, Y.; Li, B.; Luo, Z.; Chen, X.; et al. Zipper-interacting protein kinase promotes epithelial-mesenchymal transition, invasion and metastasis through AKT and NF-κB signaling and is associated with metastasis and poor prognosis in gastric cancer patients. Oncotarget 2015. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, K.; Karin, M. NF-B, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef]
- Cao, Z.; Liao, Q.; Su, M.; Huang, K.; Jin, J.; Cao, D. AKT and ERK dual inhibitors: The way forward? Cancer Lett. 2019, 459, 30–40. [Google Scholar] [CrossRef]
- van de Stolpe, A. Quantitative measurement of functional activity of the pi3k signaling pathway in cancer. Cancers 2019, 11, 293. [Google Scholar] [CrossRef] [Green Version]
- Tseng, P.C.; Chen, C.L.; Shan, Y.S.; Chang, W.T.; Liu, H.S.; Hong, T.M.; Hsieh, C.Y.; Lin, S.H.; Lin, C.F. An increase in integrin-linked kinase noncanonically confers NF-κB-mediated growth advantages to gastric cancer cells by activating ERK1/2. Cell Commun. Signal. 2014. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, O.; Naumann, M. NF-κB signaling in gastric cancer. Toxins 2017, 9, 119. [Google Scholar] [CrossRef]
- Nam, S.; Kim, J.H.; Lee, D.H. RHoA in gastric cancer: Functional roles and therapeutic potential. Front. Genet. 2019, 10, 438. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; Dijke, P. Ten TGF-β-mediated epithelial-mesenchymal transition and cancer metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Liu, H.; Jiang, M.; Zhang, S.; Chen, J.; Fan, X. Targeting MicroRNA-21 Suppresses Gastric Cancer Cell Proliferation and Migration via PTEN/Akt Signaling Axis. Cell Transplant. 2019, 28, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Fei, Z.; Zhu, R. MiR-21 modulates cisplatin resistance of gastric cancer cells by inhibiting autophagy via the PI3K/Akt/mTOR pathway. Anticancer Drugs 2020. [Google Scholar] [CrossRef]
- Fang, Y.; Shen, H.; Li, H.; Cao, Y.; Qin, R.; Long, L.; Zhu, X.; Xie, C.; Xu, W. MiR-106a confers cisplatin resistance by regulating PTEN/Akt pathway in gastric cancer cells. Acta Biochim. Biophys. Sin. 2013. [Google Scholar] [CrossRef] [Green Version]
- Hong, K.J.; Wu, D.C.; Cheng, K.H.; Chen, L.T.; Hung, W.C. RECK Inhibits Stemness Gene Expression and Tumorigenicity of Gastric Cancer Cells by Suppressing ADAM-Mediated Notch1 Activation. J. Cell. Physiol. 2014. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Z.; Gao, C.; Chen, P.; Chen, J.; Liu, W.; Xiao, S.; Lu, H. miR-21 plays a pivotal role in gastric cancer pathogenesis and progression. Lab. Investig. 2008. [Google Scholar] [CrossRef]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther.-Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, N.; He, S.; Lui, Y.; Lu, G.; Zhao, L. MicroRNA-106a targets TIMP2 to regulate invasion and metastasis of gastric cancer. FEBS Lett. 2014. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.Y.; Wang, L.P.; Wang, Q.; Han, P.; Zhuang, W.P.; Li, M.J.; Yuan, H. miR-302b regulates cell cycles by targeting CDK2 via ERK signaling pathway in gastric cancer. Cancer Med. 2016. [Google Scholar] [CrossRef]
- Tang, L.; Hu, H.; He, Y.; Mcleod, H.L.; Xiao, D.; Chen, P.; Shen, L.; Zeng, S.; Yin, X.; Ge, J.; et al. The relationship between miR-302b and EphA2 and their clinical significance in gastric cancer. J. Cancer 2018, 9, 3109–3116. [Google Scholar] [CrossRef]
- Huang, J.; He, Y.; Mcleod, H.L.; Xie, Y.; Xiao, D.; Hu, H.; Chen, P.; Shen, L.; Zeng, S.; Yin, X.; et al. miR-302b inhibits tumorigenesis by targeting EphA2 via Wnt/ β-catenin/EMT signaling cascade in gastric cancer. BMC Cancer 2017. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, X.; Zhou, F.; Jin, Z.; Chen, D.; Wang, P.; Zhang, S.; Zhuge, Y.; Shang, Y.; Zou, X. Downregulation of leucine-rich repeats and immunoglobulin-like domains 1 by microRNA-20a modulates gastric cancer multidrug resistance. Cancer Sci. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Zhou, X.; Du, Y.; Huang, Z.; Zhu, J.; Xu, J.; Cheng, G.; Shu, Y.; Liu, P.; Zhu, W.; et al. MiR-20a induces cisplatin resistance of a human gastric cancer cell line via targeting CYLD. Mol. Med. Rep. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Wang, Q.; Yuan, G.; Quan, J.; Dong, D.; Lun, Y.; Sun, B. Hsa_circ_0001649 restrains gastric carcinoma growth and metastasis by downregulation of miR-20a. J. Clin. Lab. Anal. 2020. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Wang, L.N.; Li, W.; Zuo, Q.F.; Li, M.M.; Zou, Q.M.; Xiao, B. Downregulation of miR-491-5p promotes gastric cancer metastasis by regulating SNAIL and FGFR4. Cancer Sci. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, R.; Liu, Z.; Tong, D.; Yang, Y.; Guo, B.; Wang, X.; Zhao, L.; Huang, C. MiR-491-5p, mediated by Foxi1, functions as a tumor suppressor by targeting Wnt3a/β-catenin signaling in the development of gastric cancer. Cell Death Dis. 2017. [Google Scholar] [CrossRef]
- Xu, N.; Lian, Y.J.; Dai, X.; Wang, Y.J. miR-7 Increases Cisplatin Sensitivity of Gastric Cancer Cells Through Suppressing mTOR. Technol. Cancer Res. Treat. 2017. [Google Scholar] [CrossRef] [Green Version]
- Ye, T.; Yang, M.; Huang, D.; Wang, X.; Xue, B.; Tian, N.; Xu, X.; Bao, L.; Hu, H.; Lv, T.; et al. MicroRNA-7 as a potential therapeutic target for aberrant NF-κB-driven distant metastasis of gastric cancer. J. Exp. Clin. Cancer Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Dou, W.; He, L.; Liang, S.; Tie, J.; Liu, C.; Li, T.; Lu, Y.; Mo, P.; Shi, Y.; et al. MicroRNA-7 functions as an anti-metastatic microRNA in gastric cancer by targeting insulin-like growth factor-1 receptor. Oncogene 2013. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Shan, X.; Zhou, X.; Qiu, T.; Zhu, W.; Ding, Y.; Shu, Y.; Liu, P. miR-1271 Regulates Cisplatin Resistance of Human Gastric Cancer Cell Lines by Targeting IGF1R, IRS1, mTOR, and BCL2. Anticancer. Agents Med. Chem. 2014. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, I.; Letelier, P.; Riffo-Campos, A.L.; Brebi, P.; Roa, J.C. Emerging role of mirnas in the drug resistance of gastric cancer. Int. J. Mol. Sci. 2016, 17, 424. [Google Scholar] [CrossRef] [Green Version]
- Denduluri, S.K.; Idowu, O.; Wang, Z.; Liao, Z.; Yan, Z.; Mohammed, M.K.; Ye, J.; Wei, Q.; Wang, J.; Zhao, L.; et al. Insulin-like growth factor (IGF) signaling intumorigenesis and the development ofcancer drug resistance. Genes Dis. 2015, 2, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Yang, S.; Wu, Y.; Fang, X.; Wang, Y.; Song, Y.; Han, T. MicroRNA-216a inhibits the metastasis of gastric cancer cells by targeting JAK2/STAT3-mediated EMT process. Oncotarget 2017, 8, 88870–88881. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Shi, L.; Xu, Y.; Xu, T.; Fan, R.; Cao, M.; Xu, W.; Song, J. BRD4 promotes the stemness of gastric cancer cells via attenuating miR-216a-3p-mediated inhibition of Wnt/β-catenin signaling. Eur. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, J.; Zheng, Y.; Ma, C.; Liu, X.E.; Sun, X. miR-216a-3p inhibits the proliferation, migration, and invasion of human gastric cancer cells via targeting RUNX1 and activating the NF-κB signaling pathway. Oncol. Res. 2018. [Google Scholar] [CrossRef]
- Wang, S.H.; Li, X.; Zhou, L.S.; Cao, Z.W.; Shi, C.; Zhou, C.Z.; Wen, Y.G.; Shen, Y.; Li, J.K. MicroRNA-148a suppresses human gastric cancer cell metastasis by reversing epithelial-to-mesenchymal transition. Tumor Biol. 2013. [Google Scholar] [CrossRef]
- Zheng, B.; Liang, L.; Wang, C.; Huang, S.; Cao, X.; Zha, R.; Liu, L.; Jia, D.; Tian, Q.; Wu, J.; et al. MicroRNA-148a suppresses tumor cell invasion and metastasis by downregulating ROCK1 in gastric cancer. Clin. Cancer Res. 2011. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Wang, W.; Li, Z.; Chen, Z.; Zhi, X.; Xu, J.; Li, Q.; Wang, L.; Huang, X.; Wang, L.; et al. MicroRNA-148a-3p enhances cisplatin cytotoxicity in gastric cancer through mitochondrial fission induction and cyto-protective autophagy suppression. Cancer Lett. 2017. [Google Scholar] [CrossRef]
- Li, Y.; Yan, X.; Shi, J.; He, Y.; Xu, J.; Lin, L.; Chen, W.; Lin, X.; Lin, X. Aberrantly expressed miR-188-5p promotes gastric cancer metastasis by activating Wnt/β-catenin signaling. BMC Cancer 2019. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Tan, L.; Yin, Z.; Zhu, W.; Tao, K.; Wang, G.; Shi, W.; Gao, J. MIR17HG-miR-18a/19a axis, regulated by interferon regulatory factor-1, promotes gastric cancer metastasis via Wnt/β-catenin signalling. Cell Death Dis. 2019. [Google Scholar] [CrossRef]
- Sun, G.L.; Li, Z.; Wang, W.Z.; Chen, Z.; Zhang, L.; Li, Q.; Wei, S.; Li, B.W.; Xu, J.H.; Chen, L.; et al. miR-324-3p promotes gastric cancer development by activating Smad4-mediated Wnt/beta-catenin signaling pathway. J. Gastroenterol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Zhi, X.; Tao, J.; Xiang, G.; Cao, H.; Liu, Z.; Yang, K.; Lv, C.; Ni, S. APRIL induces cisplatin resistance in gastric cancer cells via activation of the NF-κB pathway. Cell. Physiol. Biochem. 2015. [Google Scholar] [CrossRef]
- Zhang, L.; Lei, J.; Fang, Z.L.; Xiong, J.P. MiR-128b is down-regulated in gastric cancer and negatively regulates tumour cell viability by targeting PDK1/Akt/NF-κB axis. J. Biosci. 2016. [Google Scholar] [CrossRef]
- Tian, L.; Zhao, Z.; Xie, L.; Zhu, J.P. MiR-361-5p suppresses chemoresistance of gastric cancer cells by targeting FOXM1 via the PI3K/Akt/mTOR pathway. Oncotarget 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medici, D.; Hay, E.D.; Olsen, B.R. Snail and slug promote epithelial-mesenchymal transition through β-catenin-T-cell factor-4-dependent expression of transforming growth factor-β3. Mol. Biol. Cell 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Wu, D.; Li, S.; Li, X.; Wang, L.; Lu, Y.; Song, Q.; Sun, X.; Wang, X. Microrna-582 potentiates liver and lung metastasis of gastric carcinoma cells through the foxo3-mediated pi3k/akt/snail pathway. Cancer Manag. Res. 2020. [Google Scholar] [CrossRef]
- Zhu, H.; Wu, Y.; Kang, M.; Zhang, B. MiR-877 suppresses gastric cancer progression by downregulating AQP3. J. Int. Med. Res. 2020. [Google Scholar] [CrossRef]
- He, Y.; Ge, Y.; Jiang, M.; Zhou, J.; Luo, D.; Fan, H.; Shi, L.; Lin, L.; Yang, L. MiR-592 Promotes Gastric Cancer Proliferation, Migration, and Invasion Through the PI3K/AKT and MAPK/ERK Signaling Pathways by Targeting Spry2. Cell. Physiol. Biochem. 2018. [Google Scholar] [CrossRef]
- Wang, J.; Wen, T.; Li, Z.; Che, X.; Gong, L.; Yang, X.; Zhang, J.; Tang, H.; He, L.; Qu, X.; et al. MicroRNA-1224 inhibits tumor metastasis in intestinal-type gastric cancer by directly targeting FAK. Front. Oncol. 2019. [Google Scholar] [CrossRef]
- Cao, J.M.; Li, G.Z.; Han, M.; Xu, H.L.; Huang, K.M. MiR-30c-5p suppresses migration, invasion and epithelial to mesenchymal transition of gastric cancer via targeting MTA1. Biomed. Pharmacother. 2017. [Google Scholar] [CrossRef]
- Lei, C.; Du, F.; Sun, L.; Li, T.; Li, T.; Min, Y.; Nie, A.; Wang, X.; Geng, L.; Lu, Y.; et al. MIR-143 & MIR-145 inhibit gastric cancer cell migration & metastasis by suppressing MYO6. Cell Death Dis. 2017. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Ju, Y.; Ren, F.; Qiu, Y.; Tomita, Y.; Tomoeda, M.; Kishida, M.; Wang, Y.; Jin, L.; Su, F.; et al. Insulin receptor substrate 1/2 (IRS1/2) regulates Wnt/β-Catenin signaling through blocking autophagic degradation of dishevelled. J. Biol. Chem. 2014, 289, 11230–11241. [Google Scholar] [CrossRef] [Green Version]
- Takei, Y.; Suzuki, A.; Mihara, K.; Yanagihara, K. The microRNA miR-516a-3p regulates the Wnt pathway by targeting extracellular sulfatase 1 in human scirrhous gastric cancers: Anti-metastatic therapy via miRNA-based medicine. Med. Res. Arch. 2017. [Google Scholar] [CrossRef]
- Liu, Z.; Long, J.; Du, R.; Ge, C.; Guo, K.; Xu, Y. miR-204 regulates the EMT by targeting snai1 to suppress the invasion and migration of gastric cancer. Tumor Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Li, W.; Li, Y.; Wang, J.; Li, Y.; Tang, Y.; Lu, S. MicroRNA-93 promotes proliferation and metastasis of gastric cancer via targeting TIMP2. PLoS ONE 2017. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, Y.; Liu, M.; Wang, Y.; Yang, T.; Li, D.; Ding, F.; Bai, G.; Li, Q. TIMP2 is a Poor Prognostic Factor and Predicts Metastatic Biological Behavior in Gastric Cancer. Sci. Rep. 2018. [CrossRef]
- Li, R.; Yuan, W.; Mei, W.; Yang, K.; Chen, Z. MicroRNA 520d-3p inhibits gastric cancer cell proliferation, migration, and invasion by downregulating EphA2 expression. Mol. Cell. Biochem. 2014. [Google Scholar] [CrossRef]
- Song, S.Z.; Lin, S.; Liu, J.N.; Zhang, M.B.; Du, Y.T.; Zhang, D.D.; Xu, W.H.; Wang, H.B. Targeting of SPP1 by microRNA-340 inhibits gastric cancer cell epithelial–mesenchymal transition through inhibition of the PI3K/AKT signaling pathway. J. Cell. Physiol. 2019, 234, 18587–18601. [Google Scholar] [CrossRef]
- Xu, M.; Qin, S.; Cao, F.; Ding, S.; Li, M. MicroRNA-379 inhibits metastasis and epithelial-mesenchymal transition via targeting FAK/AKT signaling in gastric cancer. Int. J. Oncol. 2017, 51, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Xing, X.; Gu, F.; Zhou, G.; Liu, X.; Li, B. Ropivacaine inhibits the growth, migration and invasion of gastric cancer through attenuation of WEE1 and PI3K/AKT signaling via miR-520a-3p. OncoTargets Ther. 2020, 13, 5309–5321. [Google Scholar] [CrossRef]
- Jiang, M.; Shi, L.; Yang, C.; Ge, Y.; Lin, L.; Fan, H.; He, Y.; Zhang, D.; Miao, Y.; Yang, L. miR-1254 inhibits cell proliferation, migration, and invasion by down-regulating Smurf1 in gastric cancer. Cell Death Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhu, X.; Shou, T.; Yang, L.; Cheng, X.; Wang, J.; Deng, L.; Zheng, Y. MicroRNA-28 promotes cell proliferation and invasion in Gastric cancer via the pten/pi3k/akt signalling pathway. Mol. Med. Rep. 2018. [Google Scholar] [CrossRef] [Green Version]
- Liao, Z.; Li, Y.; Zhou, Y.; Huang, Q.; Dong, J. MicroRNA-197 inhibits gastric cancer progression by directly targeting metadherin. Mol. Med. Rep. 2018. [Google Scholar] [CrossRef] [Green Version]
- You, D.; Wang, D.; Liu, P.; Chu, Y.; Zhang, X.; Ding, X.; Li, X.; Mao, T.; Jing, X.; Tian, Z.; et al. MicroRNA-498 inhibits the proliferation, migration and invasion of gastric cancer through targeting BMI-1 and suppressing AKT pathway. Hum. Cell 2020, 33, 366–376. [Google Scholar] [CrossRef]
- Wan, H.Y.; Guo, L.M.; Liu, T.; Liu, M.; Li, X.; Tang, H. Regulation of the transcription factor NF-κB1 by microRNA-9 in human gastric adenocarcinoma. Mol. Cancer 2010. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Kang, W.; Zhang, B.; Wu, F.; Dong, Y.; Tong, J.H.M.; Yang, W.; Zhou, Y.; Zhang, L.; Cheng, A.S.L.; et al. miR-508-3p concordantly silences NFKB1 and RELA to inactivate canonical NF-ΚB signaling in gastric carcinogenesis. Mol. Cancer 2016. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Zhang, X.; Feng, X.; Fan, X.; Jin, Z. The crosstalk between microRNAs and the Wnt/β-catenin signaling pathway in cancer. Oncotarget 2017, 8, 14089–14106. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Zhang, Y.; Zhang, L.; Huang, F.; Li, J.; Wang, S. MicroRNAs, TGF-β signaling, and the inflammatory microenvironment in cancer. Tumor Biol. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Wang, J.K. The functional analysis of MicroRNAs involved in NF-κB signaling. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 1764–1774. [Google Scholar]
- Kipkeeva, F.M.; Muzaffarova, Т.А.; Nikulin, M.P.; Apanovich, P.V.; Narimanov, M.N.; Malikhova, O.A.; Kushlinskii, N.E.; Stilidi, I.S.; Karpukhin, A.V. A Group of miRNA as Candidates for Prognostic Biomarkers of Gastric Cancer Metastasis. Bull. Exp. Biol. Med. 2020, 169, 77–80. [Google Scholar] [CrossRef]
- Liu, W.; Song, N.; Yao, H.; Zhao, L.; Liu, H.; Li, G. miR-221 and miR-222 simultaneously target RECK and regulate growth and invasion of gastric cancer cells. Med. Sci. Monit. 2015. [Google Scholar] [CrossRef] [Green Version]
- Larki, P.; Ahadi, A.; Zare, A.; Tarighi, S.; Zaheri, M.; Souri, M.; Zali, M.R.; Ghaedi, H.; Omrani, M.D. Up-regulation of miR-21, miR-25, miR-93, and miR-106b in gastric cancer. Iran. Biomed. J. 2018, 22, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Tan, Z.H.; Tang, X.; Mo, M.S.; Liu, Y.P.; Gan, R.L.; Li, Y.; Zhang, L.; Li, G.Q. miR-374b-5p suppresses RECK expression and promotes gastric cancer cell invasion and metastasis. World J. Gastroenterol. 2014. [Google Scholar] [CrossRef]
- Shen, B.; Yu, S.; Zhang, Y.; Yuan, Y.; Li, X.; Zhong, J.; Feng, J. miR-590-5p regulates gastric cancer cell growth and chemosensitivity through RECK and the AKT/ERK pathway. OncoTargets Ther. 2016, 9, 6009–6019. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.D.; Cheng, J.T.; Chandoo, A.; Sun, X.W.; Zhang, L.; Lu, M.D.; Sun, W.J.; Huang, Y.P. MicroRNA-33a prevents epithelial-mesenchymal transition, invasion, and metastasis of gastric cancer cells through the snail/slug pathway. Am. J. Physiol.-Gastrointest. Liver Physiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.; He, Y.; Ouyang, K.; Xiong, H.; Li, J.; Yuan, X. microRNA-140-5p suppresses cell proliferation and invasion in gastric cancer by targeting WNT1 in the WNT/β-catenin signaling pathway. Oncol. Lett. 2018, 16, 6369–6376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Yang, B.; Lin, S.; Xing, R.; Lu, Y. Downregulation of miR-142-5p promotes tumor metastasis through directly regulating CYR61 expression in gastric cancer. Gastric Cancer 2019, 22, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Ma, J.; Yang, Y.; He, X.; Pan, X.; Wang, Z.; Qian, Y. Effects of mIR-330-3P on invasion, migration and EMT of gastric cancer cells by targeting PRRX1-mediated Wnt/β-catenin signaling pathway. OncoTargets Ther. 2020, 13, 3411–3423. [Google Scholar] [CrossRef] [Green Version]
- Tian, L.; Zhao, Z.; Xie, L.; Zhu, J.P. MiR-361-5p inhibits the mobility of gastric cancer cells through suppressing epithelial-mesenchymal transition via the Wnt/β-catenin pathway. Gene 2018, 675, 102–109. [Google Scholar] [CrossRef]
- Chen, J.Q.; Huang, Z.P.; Li, H.F.; Ou, Y.L.; Huo, F.; Hu, L. kai MicroRNA-520f-3p inhibits proliferation of gastric cancer cells via targeting SOX9 and thereby inactivating Wnt signaling. Sci. Rep. 2020. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Zhang, F.; Lin, X.; Huang, C.; Zhang, Y.; Li, Y.; Lin, J.; Chen, W.; Lin, X. MicroRNA-1225-5p inhibits proliferation and metastasis of gastric carcinoma through repressing insulin receptor substrate-1 and activation of β-catenin signaling. Oncotarget 2016. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cai, O.; Tan, S. Loxl1-as1 drives the progression of gastric cancer via regulating mir-142-5p/pik3ca axis. OncoTargets Ther. 2019, 12, 11345–11357. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Qi, T.; Yang, D.; Qi, M.; Li, D.; Xiang, X.; Huang, K.; Tong, Q. microRNA-9 Suppresses the Proliferation, Invasion and Metastasis of Gastric Cancer Cells through Targeting Cyclin D1 and Ets1. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Kogo, R.; Mimori, K.; Tanaka, F.; Komune, S.; Mori, M. Clinical significance of miR-146a in gastric cancer cases. Clin. Cancer Res. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, F.; Wang, C.; Wang, W.; Liu, W.; Wu, B. MicroRNA-31 inhibits tumor invasion and metastasis by targeting RhoA in human gastric cancer. Oncol. Rep. 2017, 38, 1133–1139. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wu, H.; Wan, X.; Liu, L.; He, Y.; Zhu, L.; Liu, S.; Yao, H.; Zhu, Z. MicroRNA-585 suppresses tumor proliferation and migration in gastric cancer by directly targeting MAPK1. Biochem. Biophys. Res. Commun. 2018. [Google Scholar] [CrossRef]
- Xu, Y.C.; Liu, X.; Li, M.; Li, Y.; Li, C.Y.; Lu, Y.; Sanches, J.; Wang, L.; Du, Y.; Mao, L.M.; et al. A Novel Mechanism of Doxorubicin Resistance and Tumorigenesis Mediated by MicroRNA-501-5p-Suppressed BLID. Mol. Ther.-Nucleic Acids 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Huang, B.; Wang, H.Y.; Chang, A.; Zheng, X.F.S. Emerging Role of MicroRNAs in mTOR Signaling. Cell. Mol. Life Sci. 2017, 74, 2613–2625. [Google Scholar] [CrossRef]
- Yan, R.; Li, K.; Yuan, D.W.; Wang, H.N.; Zhang, Y.; Dang, C.X.; Zhu, K. Downregulation of microRNA-4295 enhances cisplatin-induced gastric cancer cell apoptosis through the EGFR/PI3K/Akt signaling pathway by targeting LRIG1. Int. J. Oncol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Niu, W.; Zhang, H.; Jun, M.; Zhang, H. Downregulation of MicroRNA-147 inhibits cell proliferation and increases the chemosensitivity of gastric cancer cells to 5-Fluorouracil by directly targeting PTEN. Oncol. Res. 2018. [Google Scholar] [CrossRef]
- Jia, J.; Zhan, D.; Li, J.; Li, Z.; Li, H.; Qian, J. The contrary functions of lncRNA HOTAIR/miR-17-5p/PTEN axis and Shenqifuzheng injection on chemosensitivity of gastric cancer cells. J. Cell. Mol. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Jian, B.; Li, Z.; Xiao, D.; He, G.; Bai, L.; Yang, Q. Downregulation of microRNA-193-3p inhibits tumor proliferation migration and chemoresistance in human gastric cancer by regulating PTEN gene. Tumor Biol. 2016, 37, 8941–8949. [Google Scholar] [CrossRef]
- Huang, X.; Li, Z.; Zhang, Q.; Wang, W.; Li, B.; Wang, L.; Xu, Z.; Zeng, A.; Zhang, X.; Zhang, X.; et al. Circular RNA AKT3 upregulates PIK3R1 to enhance cisplatin resistance in gastric cancer via miR-198 suppression. Mol. Cancer 2019. [Google Scholar] [CrossRef]
- Zhou, N.; Qu, Y.; Xu, C.; Tang, Y. Upregulation of microRNA-375 increases the cisplatin-sensitivity of human gastric cancer cells by regulating ERBB2. Exp. Ther. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Han, M.; Zhou, N.; Tang, Y.; Tang, X.S. MicroRNA-495 confers increased sensitivity to chemotherapeutic agents in gastric cancer via the mammalian target of rapamycin (mTOR) signaling pathway by interacting with human epidermal growth factor receptor 2 (ERBB2). Med. Sci. Monit. 2018. [Google Scholar] [CrossRef]
- Zhang, J.X.; Xu, Y.; Gao, Y.; Chen, C.; Zheng, Z.S.; Yun, M.; Weng, H.W.; Xie, D.; Ye, S. Decreased expression of miR-939 contributes to chemoresistance and metastasis of gastric cancer via dysregulation of SLC34A2 and Raf/MEK/ERK pathway. Mol. Cancer 2017. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Gao, Y.J.; Hou, R.Z.; Ding, D.Y.; Song, D.F.; Wang, D.Y.; Feng, Y. MicroRNA-206 facilitates gastric cancer cell apoptosis and suppresses cisplatin resistance by targeting MAPK2 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, Q. Poor expression of microRNA-135b results in the inhibition of cisplatin resistance and proliferation and induces the apoptosis of gastric cancer cells through MST1-mediated MAPK signaling pathway. FASEB J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cao, T.; Guo, K.; Zhou, Y.; Liu, H.; Pan, Y.; Hou, Q.; Nie, Y.; Fan, D.; Lu, Y.; et al. Regulation of Integrin Subunit Alpha 2 by miR-135b-5p Modulates Chemoresistance in Gastric Cancer. Front. Oncol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, X.; Liu, X.; Lin, F.; Li, P.; Liu, K.; Geng, R.; Dai, C.; Lin, Y.; Tang, W.; Wu, Z.; et al. MicroRNA-421 regulated by HIF-1α promotes metastasis, inhibits apoptosis, and induces cisplatin resistance by targeting E-cadherin and caspase-3 in gastric cancer. Oncotarget 2016. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, R.; Zhang, S.; Xu, R.; Yang, Q. MicroRNA-574-3p regulates epithelial mesenchymal transition and cisplatin resistance via targeting ZEB1 in human gastric carcinoma cells. Gene 2019. [Google Scholar] [CrossRef]
- Jiang, T.; Dong, P.; Li, L.; Ma, X.; Xu, P.; Zhu, H.; Wang, Y.; Yang, B.; Liu, K.; Liu, J.; et al. MicroRNA-200c regulates cisplatin resistance by targeting ZEB2 in human gastric cancer cells. Oncol. Rep. 2017. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Q.; Pan, D.; Chen, Q.; Zhang, S.W.; Xie, D.Y.; Zheng, X.L.; Chen, H. Sensitization of Gastric Cancer Cells to 5-FU by MicroRNA-204 Through Targeting the TGFBR2-Mediated Epithelial to Mesenchymal Transition. Cell. Physiol. Biochem. 2018. [Google Scholar] [CrossRef]
- Liu, X.; Du, P.; Han, L.; Zhang, A.; Jiang, K.; Zhang, Q. Effects of miR-200a and FH535 combined with taxol on proliferation and invasion of gastric cancer. Pathol. Res. Pract. 2018. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, B.; Xu, L.; Fan, H.; Xie, J.; Wang, D. MicroRNA-146a promotes gastric cancer cell apoptosis by targeting transforming growth factor β-activated kinase 1. Mol. Med. Rep. 2017, 16, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Gao, M.; Ahmed, Y.; Gao, M.; Liu, W.; Zhang, Y.; Xie, X.; Zhao, Q.; Wang, H.; Gu, K. Microrna-362-5p enhances the cisplatin sensitivity of gastric cancer cells by targeting suppressor of zeste 12 protein. Oncol. Lett. 2019. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, J.J.; Yang, X.R.; Yu, Y.L. Upregulation of miR-34c after silencing E2F transcription facto inhibits paclitaxel combined with cisplatin resistance in gastric cancer cells. World J. Gastroenterol. 2020. [Google Scholar] [CrossRef]
- Zhu, C.Y.; Meng, F.Q.; Liu, J. MicroRNA-524-5p suppresses cell proliferation and promotes cell apoptosis in gastric cancer by regulating CASP3. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7968–7977. [Google Scholar] [CrossRef]
- Zhang, Z.; Kong, Y.; Yang, W.; Zhang, B.; Ma, F.; Liu, H.; Hua, Y. MicroRNA-218 enhances gastric cancer celcisplatin sensitivity by targeting survivin. Exp. Ther. Med. 2018. [Google Scholar] [CrossRef]
- Zhang, F.; Li, K.; Yao, X.; Wang, H.; Li, W.; Wu, J.; Li, M.; Zhou, R.; Xu, L.; Zhao, L. A miR-567-PIK3AP1-PI3K/AKT-c-Myc feedback loop regulates tumour growth and chemoresistance in gastric cancer. EBioMedicine 2019. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.; Qin, Y.; Zhi, Q.; Wang, J.; Qin, C. Knockdown of long non-coding RNA HOTAIR inhibits cisplatin resistance of gastric cancer cells through inhibiting the PI3K/Akt and Wnt/β-catenin signaling pathways by up-regulating miR-34a. Int. J. Biol. Macromol. 2018. [Google Scholar] [CrossRef]
- Yan, J.; Dang, Y.; Liu, S.; Zhang, Y.; Zhang, G. LncRNA HOTAIR promotes cisplatin resistance in gastric cancer by targeting miR-126 to activate the PI3K/AKT/MRP1 genes. Tumor Biol. 2016. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Li, C.S.; Zhou, Y.; Lu, X.H. Propofol facilitates cisplatin sensitivity via lncRNA MALAT1/miR-30e/ATG5 axis through suppressing autophagy in gastric cancer. Life Sci. 2020. [Google Scholar] [CrossRef]
- Liu, H.S.; Xiao, H.S. MicroRNAs as potential biomarkers for gastric cancer. World J. Gastroenterol. 2014, 20, 12007–12017. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Wang, L.; Yang, Y.; Gong, L.; Xiao, B.; Liu, X. A serum exosomal microRNA panel as a potential biomarker test for gastric cancer. Biochem. Biophys. Res. Commun. 2017. [Google Scholar] [CrossRef] [PubMed]
- Virgilio, E.; Giarnieri, E.; Giovagnoli, M.R.; Montagnini, M.; Proietti, A.; D’Urso, R.; Mercantini, P.; Balducci, G.; Cavallini, M. Gastric juice MicroRNAs as potential biomarkers for screening gastric cancer: A systematic review. Anticancer Res. 2018, 38, 613–616. [Google Scholar]
- Yin, L.; Xu, G.; Zhu, Y.; Wang, Y. Expression of miR-23a and miR-135 and tumor markers in gastric cancer patients and the significance in diagnosis. Oncol. Lett. 2019, 18, 5853–5858. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Liu, Y. Expression and significance of miR-24 and miR-101 in patients with advanced gastric cancer. Oncol. Lett. 2018, 16, 5769–5774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.S.; Hur, K.; Xu, G.; Choi, B.; Okugawa, Y.; Toiyama, Y.; Oshima, H.; Oshima, M.; Lee, H.J.; Kim, V.N.; et al. MicroRNA-29c mediates initiation of gastric carcinogenesis by directly targeting ITGB1. Gut 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Zhang, S.; Yang, L.; Wei, W.; Gao, J.; Guo, N.; Wu, F. Serum miR-101-3p combined with pepsinogen contributes to the early diagnosis of gastric cancer. BMC Med. Genet. 2020. [Google Scholar] [CrossRef] [Green Version]
- Peng, Q.; Shen, Y.; Lin, K.; Zou, L.; Shen, Y.; Zhu, Y. Comprehensive and integrative analysis identifies microRNA-106 as a novel non-invasive biomarker for detection of gastric cancer. J. Transl. Med. 2018. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.; Min, B.H.; Jang, J.; Kang, S.Y.; Bae, H.; Jang, S.S.; Kim, J.I.; Kim, K.M. MicroRNA Expression Profiles in Gastric Carcinogenesis. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef]
- Link, A.; Schirrmeister, W.; Langner, C.; Varbanova, M.; Bornschein, J.; Wex, T.; Malfertheiner, P. Differential expression of microRNAs in preneoplastic gastric mucosa. Sci. Rep. 2015. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Chen, Y.; Sun, D.; Wang, S.; Ding, K.; Liu, M.; Zhang, Y.; Miao, Y.; Liu, H.; Zhou, F. MicroRNA-181a functions as an oncogene in gastric cancer by targeting caprin-1. Front. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Li, Z.; Guo, Q.; Lu, Y.; Tian, T. Increased expression of miR-181d is associated with poor prognosis and tumor progression of gastric cancer. Cancer Biomark. 2019, 26, 353–360. [Google Scholar] [CrossRef]
- Tsai, M.M.; Wang, C.S.; Tsai, C.Y.; Huang, C.G.; Lee, K.F.; Huang, H.W.; Lin, Y.H.; Chi, H.C.; Kuo, L.M.; Lu, P.H.; et al. Circulating microRNA-196a/b are novel biomarkers associated with metastatic gastric cancer. Eur. J. Cancer 2016. [Google Scholar] [CrossRef]
- Huang, Z.S.; Guo, X.W.; Zhang, G.; Liang, L.X.; Nong, B. The diagnostic and prognostic value of miR-200c in gastric cancer: A meta-analysis. Dis. Markers 2019. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Liang, Y.; Ma, M.H.; Wu, K.Z.; Zhang, C.D.; Dai, D.Q. Downregulation of microRNA-376a in gastric cancer and association with poor prognosis. Cell. Physiol. Biochem. 2018, 51, 2010–2018. [Google Scholar] [CrossRef]
- Chen, J.; Wu, L.; Sun, Y.; Yin, Q.; Chen, X.; Liang, S.; Meng, Q.; Long, H.; Li, F.; Luo, C.; et al. MIR-421 in plasma as a potential diagnostic biomarker for precancerous gastric lesions and early gastric cancer. PeerJ 2019. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, Y.; Yao, J.; Li, R.; Fan, X. Downregulation of miR-484 is associated with poor prognosis and tumor progression of gastric cancer. Diagn. Pathol. 2020, 15, 1–7. [Google Scholar] [CrossRef]
- Cai, H.; Lin, H.; Cao, W.; Sun, J.; Huang, Y.; Fang, Y. Downregulation of miR-519a Predicts Poor Prognosis and Contributes to Tumor Progression in Gastric Cancer. Oncol. Res. Treat. 2020, 43, 19–26. [Google Scholar] [CrossRef]
- Su, H.; Ren, F.; Jiang, H.; Chen, Y.; Fan, X. Upregulation of microRNA-520a-3p inhibits the proliferation, migration and invasion via spindle and kinetochore associated 2 in gastric cancer. Oncol. Lett. 2019. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhu, M.; Liao, B.; Tian, T.; Li, M.; Wang, Z.; Chen, G. Upregulation of miR-552 Predicts Unfavorable Prognosis of Gastric Cancer and Promotes the Proliferation, Migration, and Invasion of Gastric Cancer Cells. Oncol. Res. Treat. 2020, 43, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Min, C.; Zhang, A.; Qin, J. Increased expression of miR-601 is associated with poor prognosis and tumor progression of gastric cancer. Diagn. Pathol. 2019, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- An, J.X.; Ma, Z.S.; Ma, M.H.; Shao, S.; Cao, F.L.; Dai, D.Q. MiR-1236-3p serves as a new diagnostic and prognostic biomarker for gastric cancer. Cancer Biomark. 2019, 25, 127–132. [Google Scholar] [CrossRef]
- Shang, Y.; Feng, B.; Zhou, L.; Ren, G.; Zhang, Z.; Fan, X.; Sun, Y.; Luo, G.; Liang, J.; Wu, K.; et al. The miR27b-CCNG1-P53-miR-508-5p axis regulates multidrug resistance of gastric cancer. Oncotarget 2016. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Zhao, L.; Xu, X.U.; Xu, H. Analysis of microRNA expression profiles reveals a 5-microRNA prognostic signature for predicting overall survival time in patients with gastric adenocarcinoma. Oncol. Rep. 2019. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, W.; Yang, Y.; Du, L.; Yang, X.; Zheng, G.; Duan, W.; Wang, R.; Zhang, X.; Wang, L. Identification of circulating microRNA signatures as potential noninvasive biomarkers for prediction and prognosis of lymph node metastasis in gastric cancer. Oncotarget 2017, 8, 65132–65142. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, H.; Zhou, X.; Wang, T.; Zhang, J.; Zhu, W.; Zhu, H.; Cheng, W. Five serum-based miRNAs were identified as potential diagnostic biomarkers in gastric cardia adenocarcinoma. Cancer Biomark. 2018. [Google Scholar] [CrossRef]
- Huang, Z.; Zhu, D.; Wu, L.; He, M.; Zhou, X.; Zhang, L.; Zhang, H.; Wang, W.; Zhu, J.; Cheng, W.; et al. Six serum-based miRNAs as potential diagnostic biomarkers for gastric cancer. Cancer Epidemiol. Biomark. Prev. 2017. [Google Scholar] [CrossRef] [Green Version]
- Link, A.; Kupcinskas, J. MicroRNAs as non-invasive diagnostic biomarkers for gastric cancer: Current insights and future perspectives. World J. Gastroenterol. 2018, 24, 3313–3329. [Google Scholar] [CrossRef] [PubMed]
- Rekker, K.; Saare, M.; Roost, A.M.; Kaart, T.; Sõritsa, D.; Karro, H.; Sõritsa, A.; Simón, C.; Salumets, A.; Peters, M. Circulating miR-200-family micro-RNAs have altered plasma levels in patients with endometriosis and vary with blood collection time. Fertil. Steril. 2015. [Google Scholar] [CrossRef] [PubMed]
- Suryawanshi, S.; Vlad, A.M.; Lin, H.M.; Mantia-Smaldone, G.; Laskey, R.; Lee, M.; Lin, Y.; Donnellan, N.; Klein-Patel, M.; Lee, T.; et al. Plasma MicroRNAs as novel biomarkers for endometriosis and endometriosis-associated ovarian cancer. Clin. Cancer Res. 2013. [Google Scholar] [CrossRef] [Green Version]
- Jia, S.Z.; Yang, Y.; Lang, J.; Sun, P.; Leng, J. Plasma miR-17-5p, miR-20a and miR-22 are down-regulated in women with endometriosis. Hum. Reprod. 2013, 28, 322–330. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Yang, S.; Wang, K.; Zhong, Q.; Ma, A.; Pan, X. Plasma miR-126 and miR-143 as Potential Novel Biomarkers for Cerebral Atherosclerosis. J. Stroke Cerebrovasc. Dis. 2019. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, C.; Hu, Z.; Li, G.; Wang, C.; Yang, C.; Huang, D.; Chen, X.; Zhang, H.; Zhuang, R.; et al. A five-microRNA signature identified from genome-wide serum microRNA expression profiling serves as a fingerprint for gastric cancer diagnosis. Eur. J. Cancer 2011. [Google Scholar] [CrossRef]
- Zhu, C.; Ren, C.; Han, J.; Ding, Y.; Du, J.; Dai, N.; Dai, J.; Ma, H.; Hu, Z.; Shen, H.; et al. A five-microRNA panel in plasma was identified as potential biomarker for early detection of gastric cancer. Br. J. Cancer 2014, 2291–2299. [Google Scholar] [CrossRef]
- Kim, D.N.; Chae, H.-S.; Oh, S.T.; Kang, J.-H.; Park, C.H.; Park, W.S.; Takada, K.; Lee, J.M.; Lee, W.-K.; Lee, S.K. Expression of Viral MicroRNAs in Epstein-Barr Virus-Associated Gastric Carcinoma. J. Virol. 2007. [Google Scholar] [CrossRef] [Green Version]
- Treece, A.L.; Duncan, D.L.; Tang, W.; Elmore, S.; Morgan, D.R.; Dominguez, R.L.; Speck, O.; Meyers, M.O.; Gulley, M.L. Gastric adenocarcinoma microRNA profiles in fixed tissue and in plasma reveal cancer-associated and Epstein-Barr virus-related expression patterns. Lab. Investig. 2016. [Google Scholar] [CrossRef] [Green Version]
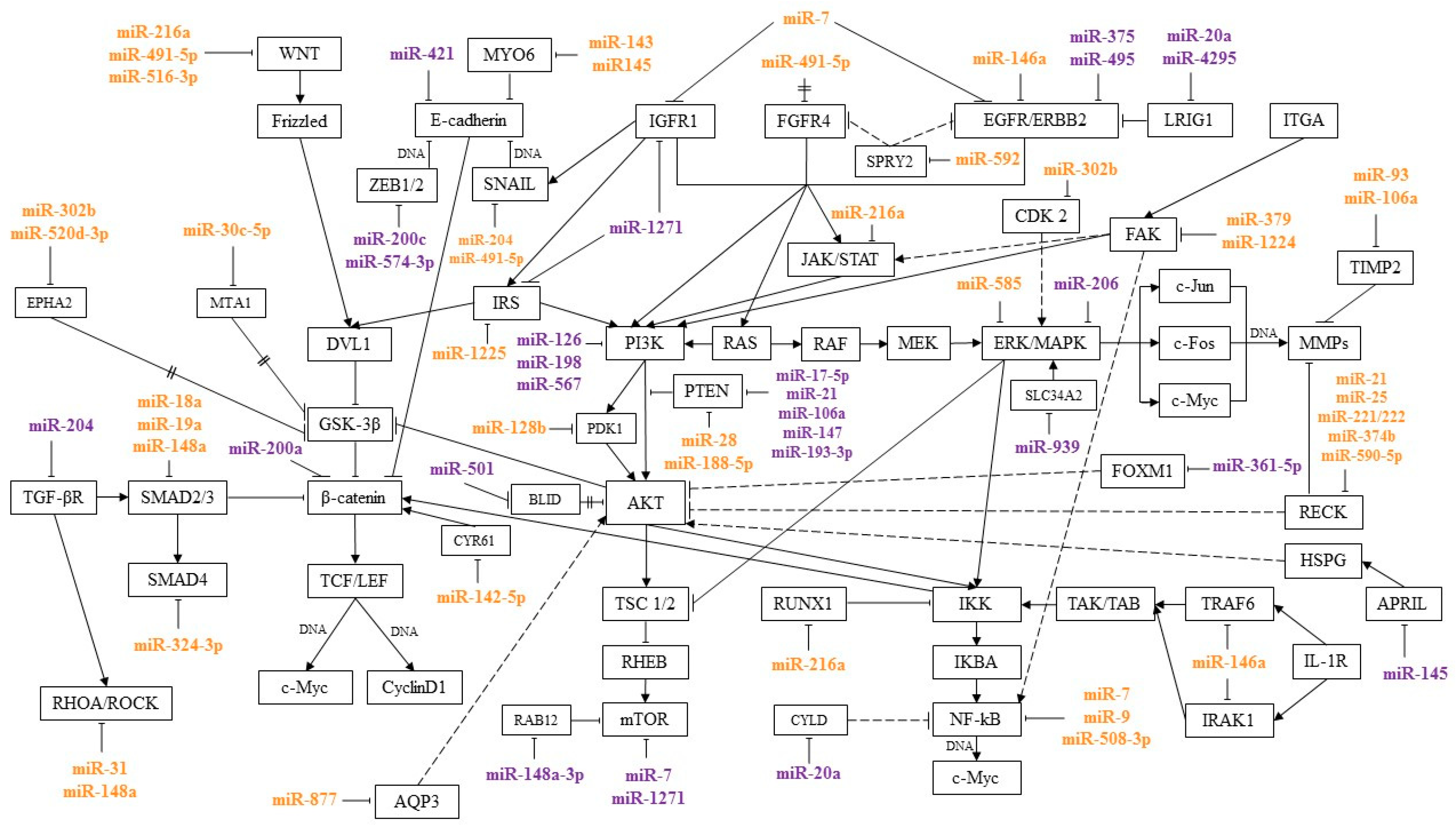
 activation;
activation;  inhibition;
inhibition;  interaction is assumed;
interaction is assumed;  mediated interaction.
activation; inhibition; interaction is assumed; mediated interaction.
mediated interaction.
activation; inhibition; interaction is assumed; mediated interaction.


{kind=link}
{kind=link}
| Signal Pathway | MicroRNA | Target | Role in GC Metastasis | Materials | Reference |
|---|---|---|---|---|---|
| Wnt/β-catenin | miR-18a miR-19a | SMAD2 | Overexpression of miR-18a was associated with lymph node metastasis and the distant metastases development. Overexpression of miR-19a was associated with lymph node metastasis. | GC tissues | [49] |
| miR-30c-5p | MTA1 | Decreased expression level of miR-30c-5p was associated with lymph nodes metastasis. | GC tissues | [59] | |
| miR-33a | SNAI2 | Overexpression of miR-33a inhibited metastasis and tumor growth in vivo. | GC tissues, cell lines and nude mice | [84] | |
| miR-93 miR-106a | TIMP2 | Overexpression of miR-106a and miR-93 was associated with lymph node metastasis and vascular invasion. | FFPE samples, cell lines and GC tissues | [27,64] | |
| miR-140-5p | WNT1 | Decreased expression level of miR-140-5p was associated with lymph nodes metastasis. | GC tissues | [85] | |
| miR-142-5p | CYR61 | Downregulation of miR-142-5p was associated with the metastases development, relapse and poor prognosis. | GC tissues | [86] | |
| miR-143 miR-145 | MYO6 | miR-143, miR-145 inhibit EMT and metastasis. | GC tissues, cell lines and nude mice | [60] | |
| miR-188-5p | PTEN | Overexpression of miR-188-5p was associated with lymph nodes metastasis and the distant metastases development. | GC tissues | [48] | |
| miR-204 | SNAIL | Decreased expression level of miR-204 was associated with the metastases development. | GC tissues, cell lines and nude mice | [63] | |
| miR-302b miR-520d-3p | EPHA2 | Downregulation of miR-302b correlated with the depth of tumor invasion, lymph nodes metastasis, and later stage of GC. Downregulation of miR-520d-3p correlated with increased tumor invasion, lymph node metastasis, and clinical stage. | GC tissues, cell lines and nude mice | [29,30,66] | |
| miR-330-3p | PRRX1 | Negative correlation between expression level of miR-330-3p and tumor size, TNM stage, and the degree of lymph nodes metastasis | GC tissues | [87] | |
| miR-361-5p | Indirectly affects SNAIL, E-cadherin, β-catenin | Decreased expression level of miR-361-5p correlated with lymph node metastasis and the distant metastases development. | GC tissues, cell lines and nude mice | [88] | |
| miR-516a-3p | SULF1 | Decreased expression level of miR-516a-3p was associated with the spread of metastases in the peritoneum. | Nude mice | [62] | |
| miR-520f-3p | SOX9 | Decreased expression of miR-520f-3p correlated with the depth of tumor invasion, metastases development, and poor prognosis in GC patients. | GC tissues | [89] | |
| miR-1225-5p | IRS1 | Decreased expression level of miR-1225-5p correlated with the depth of tumor invasion, lymph node metastasis, and distant metastases. | GC tissues, cell lines and nude mice | [90] | |
| PI3K/AKT/mTOR | miR-28 | PTEN | MiR-28 knockdown inhibited the proliferative and invasive properties of GC cells. | Cell lines | [71] |
| miR-142-5p | PIK3CA | Overexpression of miR-142-5p inhibited cell proliferation and tumor growth and metastasis. | GC tissues, cell lines and nude mice | [91] | |
| miR-197 | MTDH | Revealed a negative correlation between the expression level of miR-197 and the size of the tumor, the depth of invasion and lymph nodes metastasis. | GC tissues | [72] | |
| miR-340 | SPP1 | Overexpression of miR-340 induced apoptosis, inhibited proliferation, invasion, in vitro migration, and in vivo tumor growth. | Cell lines and nude mice | [67] | |
| miR-379 | FAK | miR-379 was associated with lymph nodes metastasis. | GC tissues | [68] | |
| miR-498 | BMI-1 | Decreased expression level of miR-498 was associated with lymph node metastasis and poor prognosis in GC patients. | GC tissues | [73] | |
| miR-520a-3p | WEE1 | Induced expression of miR-520a-3p decreased tumor size. | Cell lines and nude mice | [69] | |
| miR-1254 | SMURF1 | Decreased expression level of miR-1254 was associated with an increase in tumor size and the degree of lymph nodes metastasis. | GC tissues, cell lines and nude mice | [70] | |
| NF-κB | miR-9 | D1, ETS1, NF-κB1 | Decreased expression level of miR-9 was associated with lymph nodes metastasis and the distant metastases development. | GC tissues, cell lines and nude mice | [74,92] |
| miR-216a-3p | RUNX1 | Increased miR-216 expression correlated with lymph node metastasis. | GC tissues | [44] | |
| miR-508-3p | NFKB1 | Overexpression of miR-508-3p inhibited the invasive properties of GC cells in vitro. In addition, miR-508-3p suppressed the level of MMP9 expression. | Cell lines and nude mice | [75] | |
| NF-kB; Wnt/β-catenin | miR-7 | RELA, IGF1R | MiR-7 transfection inhibits distant metastases in vivo. | GC tissues, cell lines and nude mice | [37,38] |
| miR-146a | IRAK1, EGFR | Decreased expression level of miR-146a was associated with lymph node metastases. | GC tissues | [93] | |
| JAK/STAT | miR-216a | JAK2 | Decreased expression level of miR-216 correlated with lymph node metastasis. | GC tissues and cell lines | [42] |
| NF-κB; JAK/STAT | miR-1224 | FAK | Decreased expression level of miR-1224 was associated with metastasis to lymph nodes in intestinal type of GC. | GC tissues and FFPE tissues | [58] |
| RHOA-ROCK | miR-31 | RHOA | Decreased expression level of miR-31 was associated with lymph nodes metastasis. It was shown that miR-31 inhibited the development of distant metastases. | GC tissues, cell lines and nude mice | [94] |
| RAS/RAF/EKK/MAPK | miR-302b | CDK2 | Decreased expression level of miR-302b was associated with lymph node metastasis and advanced GC. | GC tissue | [28] |
| miR-585 | MAPK1 | Decreased expression level of miR-585 was associated with tumor invasion, lymph node metastasis, and advanced GC. | GC tissues, cell lines and nude mice | [95] | |
| PI3K/AKT/mTOR; RAS/RAF/EKK/MAPK; JAK/STAT and Notch | miR-21 miR-25 miR-221/222 miR-374b-5p miR-590-5p | RECK | Overexpression of these microRNAs was associated with lymph nodes metastasis. | GC tissues and cell lines | [80,81,82,83] |
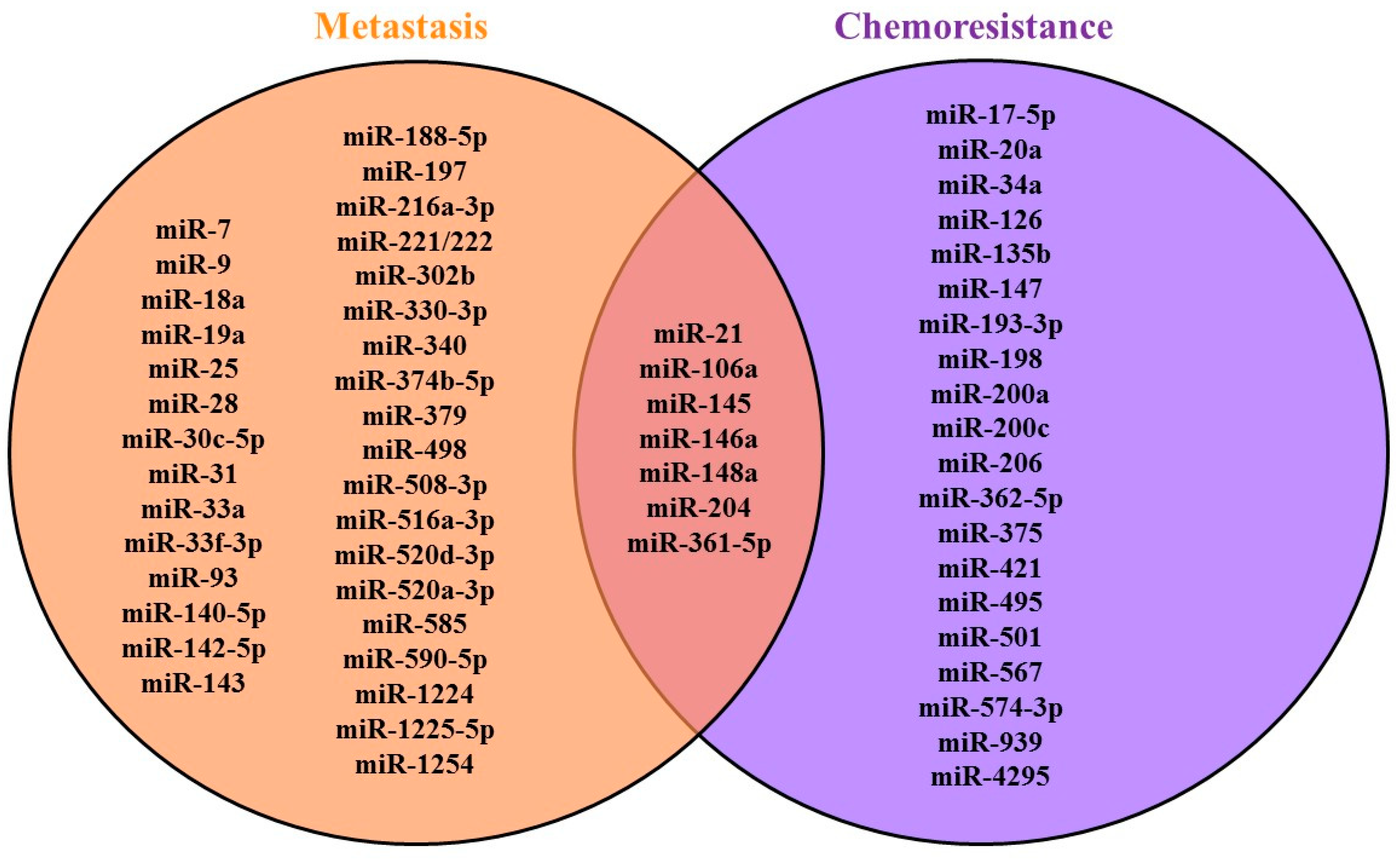
| Signaling Pathway | Process | ||
|---|---|---|---|
| Apoptosis | Apoptosis; Outflow of Drugs from the Cell | Autophagy | |
| PI3K/AKT/mTOR | miR-17-5p miR-21 miR-106a miR-147 miR-193-3p miR-198 miR-375 miR-495 miR-501 miR-4295 | miR-20a miR-126 miR-567 | miR-148a miR-361-5p |
| RAS/RAF/ERK/MAPK | miR-135b miR-206 miR-939 | miR-20a | |
| EMT (Wnt/β-catenin) | miR-200a miR-200c miR-204 miR-421 miR-574-3p | miR-34a | |
| NF-κB | miR-145 miR-146a miR-362-5p | miR-20a | |
| MicroRNA | Characteristics of microRNAs as Candidates for GC Markers | Material/Method | Reference |
|---|---|---|---|
| miR-9 | Lymph nodes metastasis (p < 0.001) and distant metastases (p = 0.022). | GC tissue samples | [92] |
| miR-19b miR-106a | Diagnostics: AUC = 0.814, sensitivity = 95%, and specificity = 90%. | Circulating exosomal microRNAs | [124] |
| miR-21 miR-106a | Difference in expression in gastric and non-cancerous cancers (p < 0.001). | Gastric juice | [125] |
| miR-23a miR-135 | Diagnostics: miR-23a specificity, sensitivity, and AUC are 67.95, 87.50, and 0.805%, respectively, for miR-135 at 73.08, 82.50, and 0.824%, respectively. | Serum | [126] |
| miR-24 miR-101 | OS (p < 0.01). | GC tissue samples | [127] |
| miR-29c | Decreased expression level in tumor tissue compared to unaffected mucosa (p < 0.0001). | GC tissue samples | [128] |
| miR-30c-5p | Lymph node metastasis (p = 0.014). | GC tissue samples | [59] |
| miR-93 | Lymph node metastasis (p < 0.01). | GC tissue samples | [64] |
| miR-101-3p | When distinguishing atrophic gastritis and gastric cancer: AUC = 0.8749, sensitivity = 72.09%, and specificity = 86.49%. | Serum | [129] |
| miR-106 | Sensitivity = 0.71, specificity = 0.82, and AUC = 0.80. | Serum, tissue, plasma and gastric juice | [130] |
| miR-106a | Lymph node metastasis (p = 0.002), vascular invasion (p = 0.017), and the depth of tumor invasion (p = 0.009). | GC tissue samples | [27] |
| miR-107 | The expression level was increased in adenoma with high-grade dysplasia p = 0.006 and in the early stages of GC p = 0.03. | Microarrays, validation by RT-PCR | [131] |
| miR-129 | Significantly lower levels of miR-129-1-3p (p = 0.007), miR-129-2-3p (p = 0.003), and a combination of two microRNAs (p = 0.003) in GC patients compared to patients with benign stomach diseases. | In gastric juice | [125] |
| miR-140-5p | Lymph node metastasis (p = 0.018); survival—PFS, p < 0.05, OS, p < 0.05). | GC tissue samples | [85] |
| miR-155 miR-223 | Atrophic gastritis (p < 0.0001); gastric cancer (p < 0.05). | GC and AG tissue samples | [132] |
| miR-181a | Lymph node metastasis (p = 0.0124); distant metastases (p = 0.0376). | GC tissue samples | [133] |
| miR-181d | Lymph node metastasis (p < 0.05) and overall survival (p = 0.001). | GC tissue samples | [134] |
| miR-196a | Survival: (p = 0.032, HR = 3.057, 95% CI = 1.1–8.495); Diagnostics: AUC = 0.864, sensitivity 69.5%, specificity 97.6%; metastasis p < 0.001. | Plasma | [135] |
| miR-196b | Survival: p = 0.042, HR = 2.914, 95% CI = 1.036–8.174; diagnostics: AUC = 0.811, sensitivity 62.2%, and specificity 96.1%. | Plasma | [135] |
| miR-197 | Depth of invasion (p = 0.005), lymph node metastases (p = 0.004). | GC tissue samples | [72] |
| miR-200c | Had a prognostic value and moderately diagnostic. Expression data were inconsistent. | Meta-analysis | [136] |
| miR-302b | Lymph node metastases (p = 0.003), OS (HR = 1.86; 95% CI, 1.11–3.14; p = 0.021). | GC tissue samples | [29] |
| miR-330-3p | Lymph node metastasis (p < 0.001). | Serum, GC tissue samples | [87] |
| miR-361-5p | Lymph node metastasis, distant metastases development (p < 0.001). | GC tissue samples | [88] |
| miR-376a | Metastasis to regional lymph nodes (p = 0.02) and poor prognosis (p = 0.02). | Tissues and cell lines of GC | [137] |
| miR-379 | Lymph node metastasis (p < 0.001), OS (p = 0.0007), and PHS (p = 0.0002). | GC tissue samples | [68] |
| miR-421 | Difference in expression among patients with benign and malignant gastric diseases (p < 0.001). | Gastric juice | [125] |
| OS (p = 0.016, HR 2.586, 95% CI 1.194–5.599) and RFS (p = 0.014, HR 2.465, 95% CI 1.201–5.060). | GC tissue samples | [109] | |
| Early stages. Sensitivity 96.67; specificity 95.56; AUC 0.981 (0.942–0.997); p <0.0001. | Serum | [138] | |
| miR-484 | Lymph node metastasis (p = 0.015), distant metastases development (p = 0.005), stage of the disease (p = 0.002), and degree of differentiation (p = 0.006). | GC tissue samples | [139] |
| miR-519a | Lymph node metastasis, degree of differentiation, and stage of the disease (p < 0.05); OS (p = 0.002). | GC tissue samples | [140] |
| miR-520a-3p | Depth of tumor invasion (p < 0.001) and stage of the disease (p < 0.05). | GC tissue samples | [141] |
| miR-552 | Lymph node metastasis (p = 0.018) and OS (p = 0.011), HR = 5.657, 95% CI 1.619–19.761. | GC tissue samples | [142] |
| miR-585 | Depth of tumor invasion (p < 0.010), lymph node metastasis (p < 0.002). | GC tissue samples | [95] |
| miR-601 | Invasion, lymph node metastasis, and the distant metastases development (p < 0.05); OS (p = 0.001). | GC tissue samples | [143] |
| miR-1225-5p | Depth of tumor invasion (p = 0.016), spread of metastases to lymph nodes (p = 0.002), and development of distant metastases (p = 0.01). | GC tissue samples | [90] |
| miR-1236-3p | Lymph node metastasis (p = 0.005), disease stage (p = 0.001), and degree of differentiation (p = 0.001). | GC tissue samples | [144] |
| Potential markers of response to chemotherapy | |||
| miR-27b miR-508-5p | Response to chemotherapy (p = 0.02 and p = 0.04, respectively). | GC tissue samples | [145] |
| miR-939 | Potential marker of sensitivity to chemotherapy; AUC = 0.777, p < 0.001. | GC tissue samples | [105] |
| MicroRNA panels | |||
| miR-7-2 miR-9-3 miR-548o miR-1255a miR-3687 | Patient survival AUC = 0.9 (HR, 2.840; 95% CI, 1.937–4.162; p < 0.01). | TCGA database | [146] |
| miR-143-3p miR-146a miR-451a miR-501-3p | Predicting the development of lymph node metastases in GC; AUC = 0.822 (95% CI, 0.758 to 0.875), specificity = 87.78%, and sensitivity = 63.33%. | Serum | [147] |
| miR-22-5p miR-132-3p miR-200a-3p miR-485-3p miR-2965p | Suggested for diagnostics of GC AUC= 0.724. | Serum | [148] |
| miR-10b-5p miR-20a-3p miR-132-3p miR-185-5p miR-195-5p miR-296-5p | Suggested for diagnostics of GC AUC = 0.702. | Serum | [149] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kipkeeva, F.; Muzaffarova, T.; Korotaeva, A.; Nikulin, M.; Grishina, K.; Mansorunov, D.; Apanovich, P.; Karpukhin, A. MicroRNA in Gastric Cancer Development: Mechanisms and Biomarkers. Diagnostics 2020, 10, 891. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10110891
Kipkeeva F, Muzaffarova T, Korotaeva A, Nikulin M, Grishina K, Mansorunov D, Apanovich P, Karpukhin A. MicroRNA in Gastric Cancer Development: Mechanisms and Biomarkers. Diagnostics. 2020; 10(11):891. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10110891
Chicago/Turabian StyleKipkeeva, Fatimat, Tatyana Muzaffarova, Alexandra Korotaeva, Maxim Nikulin, Kristina Grishina, Danzan Mansorunov, Pavel Apanovich, and Alexander Karpukhin. 2020. "MicroRNA in Gastric Cancer Development: Mechanisms and Biomarkers" Diagnostics 10, no. 11: 891. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10110891