Validation of PfSNP-LAMP-Lateral Flow Dipstick for Detection of Single Nucleotide Polymorphism Associated with Pyrimethamine Resistance in Plasmodium falciparum

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Blood Sample Collection and Genomic DNA Extraction
2.2. DNA Sequencing
2.3. Recombinant Plasmid Construction
2.4. PfSNP-LAMP-LFD Conditions
3. Results
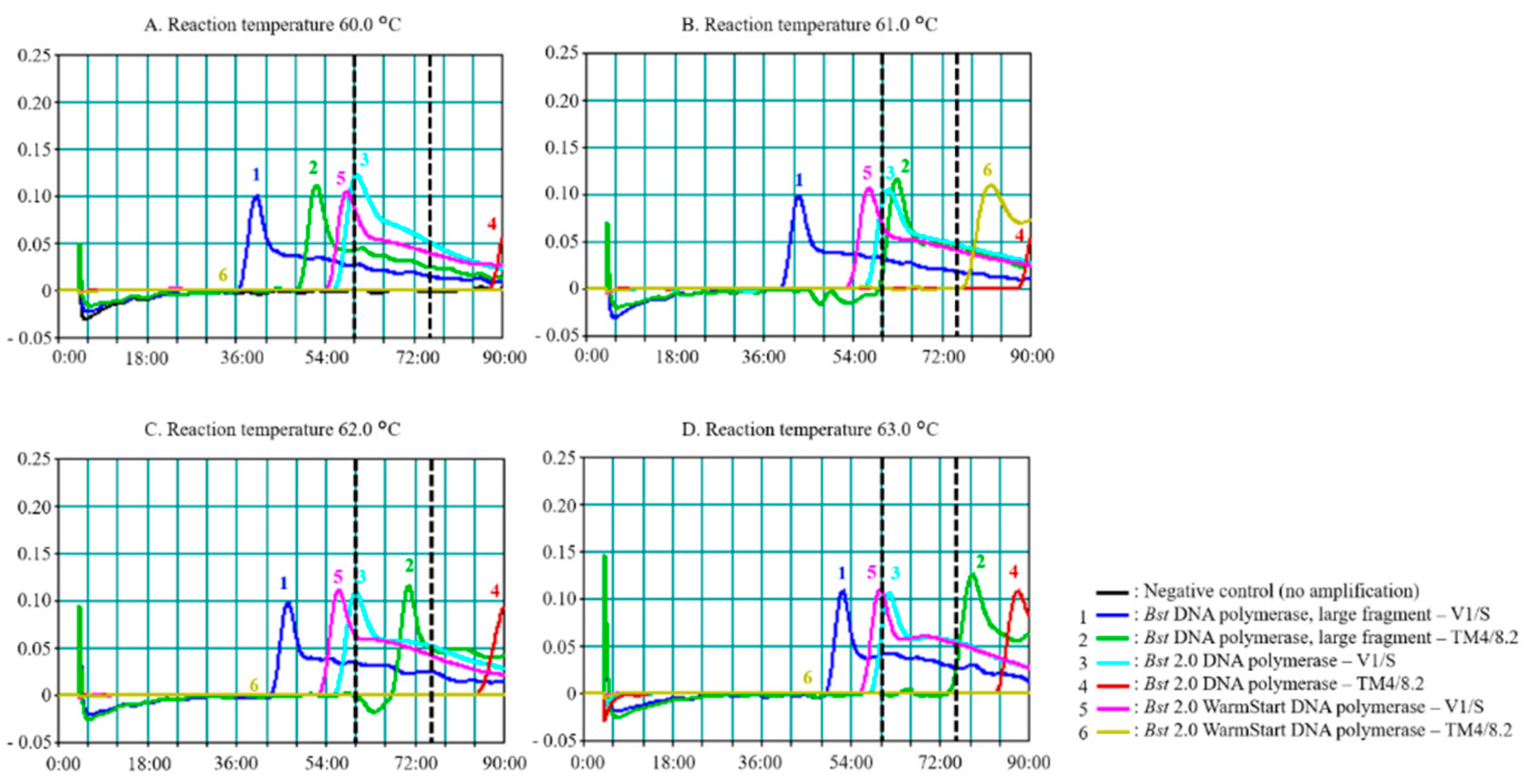
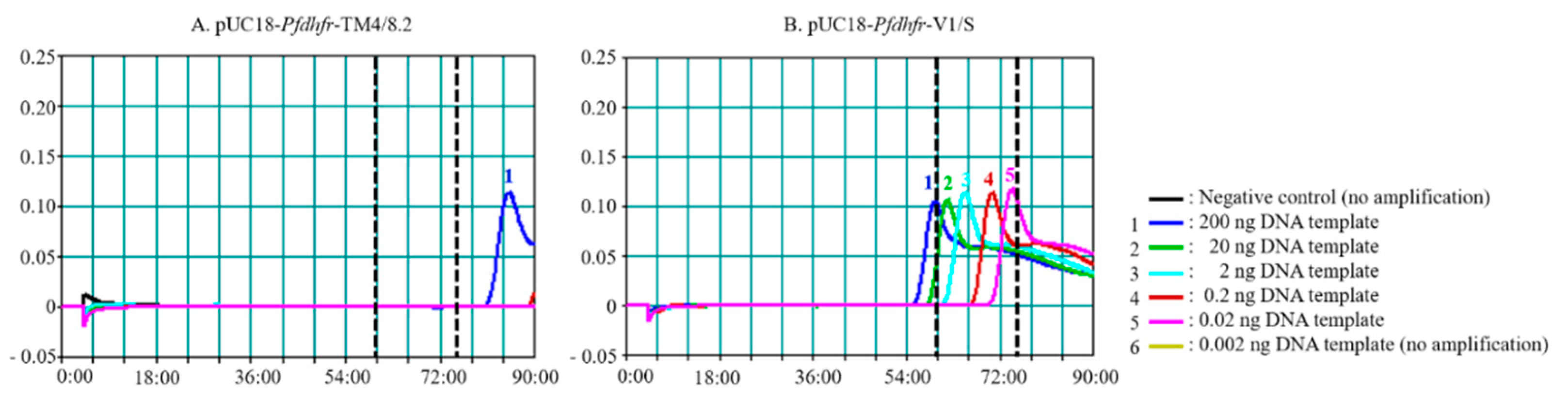
3.1. Effects of Enzyme and Reaction Time on PfSNP-LAMP Sensitivity and Specificity
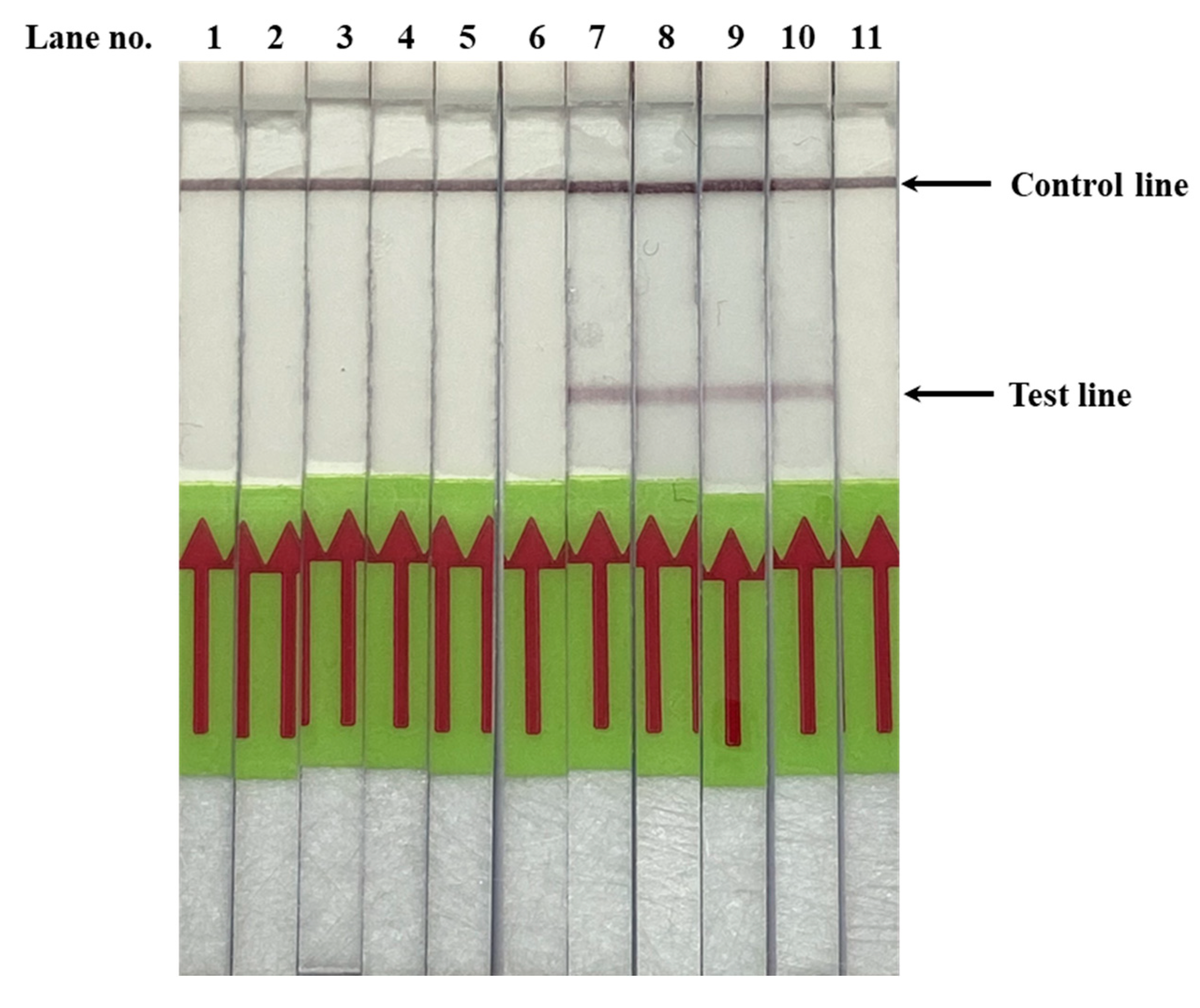
3.2. Validation of PfSNP-LAMP-LFD in Clinical Blood Samples from Malaria Patients
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- WHO. WHO Seasonal Malaria Chemoprevention with Sulfadoxine–Pyrimethamine plus Amodiaquine in Children: A Field Guide. August 2013; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- WHO. WHO Policy Brief for the Implementation of Intermittent Preventive Treatment of Malaria in Pregnancy Using Sulfadoxine-Pyrimethamine (IPTp-SP), April 2013 (rev. January 2014); World health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Bzik, D.J.; Li, W.B.; Horii, T.; Inselburg, J. Molecular cloning and sequence analysis of the Plasmodium falciparum dihydrofolate reductase-thymidylate synthase gene. Proc. Natl. Acad. Sci. USA 1987, 84, 8360–8364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Triglia, T.; Cowman, A.F. Primary structure and expression of the dihydropteroate synthetase gene of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1994, 91, 7149–7153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowman, A.F.; Morry, M.J.; Biggs, B.A.; Cross, G.A.; Foote, S.J. Amino acid changes linked to pyrimethamine resistance in the dihydrofolate reductase-thymidylate synthase gene of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1988, 85, 9109–9113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, D.S.; Walliker, D.; Wellems, T.E. Evidence that a point mutation in dihydrofolate reductase-thymidylate synthase confers resistance to pyrimethamine in falciparum malaria. Proc. Natl. Acad. Sci. USA 1988, 85, 9114–9118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorkman, A.; Phillips-Howard, P.A. The epidemiology of drug-resistant malaria. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Peterson, D.S.; Milhous, W.K.; Wellems, T.E. Molecular basis of differential resistance to cycloguanil and pyrimethamine in Plasmodium falciparum malaria. Proc. Natl. Acad. Sci. USA 1990, 87, 3018–3022. [Google Scholar] [CrossRef] [Green Version]
- Plowe, C.V.; Cortese, J.F.; Djimde, A.; Nwanyanwu, O.C.; Watkins, W.M.; Winstanley, P.A.; Estrada-Franco, J.G.; Mollinedo, R.E.; Avila, J.C.; Cespedes, J.L.; et al. Mutations in Plasmodium falciparum dihydrofolate reductase and dihydropteroate synthase and epidemiologic patterns of pyrimethamine-sulfadoxine use and resistance. J. Infect. Dis. 1997, 176, 1590–1596. [Google Scholar] [CrossRef] [Green Version]
- Yuthavong, Y.; Vilaivan, T.; Chareonsethakul, N.; Kamchonwongpaisan, S.; Sirawaraporn, W.; Quarrell, R.; Lowe, G. Development of a lead inhibitor for the A16V+S108T mutant of dihydrofolate reductase from the cycloguanil-resistant strain (T9/94) of Plasmodium falciparum. J. Med. Chem. 2000, 43, 2738–2744. [Google Scholar] [CrossRef]
- Wongsrichanalai, C.; Pickard, A.L.; Wernsdorfer, W.H.; Meshnick, S.R. Epidemiology of drug-resistant malaria. Lancet Infect. Dis. 2002, 2, 209–218. [Google Scholar] [CrossRef]
- Nair, S.; Williams, J.T.; Brockman, A.; Paiphun, L.; Mayxay, M.; Newton, P.N.; Guthmann, J.P.; Smithuis, F.M.; Hien, T.T.; White, N.J.; et al. A selective sweep driven by pyrimethamine treatment in southeast asian malaria parasites. Mol. Biol. Evol. 2003, 20, 1526–1536. [Google Scholar] [CrossRef]
- Roper, C.; Pearce, R.; Nair, S.; Sharp, B.; Nosten, F.; Anderson, T.J.C. Intercontinental spread of pyrimethamine—Resistant malaria. Science 2004, 305, 1124. [Google Scholar] [CrossRef] [PubMed]
- Plowe, C.V.; Kublin, J.G.; Doumbo, O.K. P. falciparum dihydrofolate reductase and dihydropteroate synthase mutations: Epidemiology and role in clinical resistance to antifolates. Drug Resist. Updat. 1998, 1, 389–396. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchai, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eiken Chemical Co., Ltd. Available online: https://www.eiken.co.jp/en/products/lamp/ (accessed on 6 November 2020).
- Poon, L.L.; Wong, B.W.Y.; Ma, E.H.T.; Chan, K.H.; Chow, L.M.C.; Abeyewickreme, W.; Tangpukdee, N.; Yuen, K.Y.; Guan, Y.; Looareesuwan, S.; et al. Sensitive and inexpensive molecular test for falciparum malaria: Detecting Plasmodium falciparum DNA directly from heat-treated blood by loop-mediated isothermal amplification. Clin. Chem. 2006, 52, 303–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, E.T.; Watanabe, R.; Sattabongkot, J.; Khuntirat, B.; Sirichaisinthop, J.; Iriko, H.; Jin, L.; Takeo, S.; Tsuboi, T. Detection of four Plasmodium species by genus and species-specific loop-mediated isothermal amplification for clinical diagnosis. J. Clin. Microbiol. 2007, 45, 2521–2528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buates, S.; Bantuchai, S.; Sattabongkot, J.; Han, E.T.; Tsuboi, T.; Udomsangpetch, R.; Sirichaisinthop, J.; Tan-ariya, P. Development of a reverse transcription-loop-mediated isothermal amplification (RT-LAMP) for clinical detection of Plasmodium falciparum gametocytes. Parasitol. Int. 2010, 59, 414–420. [Google Scholar] [CrossRef]
- Yongkiettrakul, S.; Jaroenram, W.; Arunrut, N.; Chareanchim, W.; Pannehpetch, S.; Suebsing, R.; Kiatpathomchai, W.; Pornthanakasem, W.; Yuthavong, Y.; Kongkasuriyachai, D. Application of loop-mediated isothermal amplification assay combined with lateral flow dipstick for detection of Plasmodium falciparum and Plasmodium vivax. Parasitol. Int. 2014, 63, 777–784. [Google Scholar] [CrossRef]
- Jiang, Y.S.; Bhadra, S.; Li, B.; Wu, Y.R.; Milligan, J.N.; Ellington, A.D. Robust strand exchange reactions for the sequence-specific, real-time detection of nucleic acid amplicons. Anal. Chem. 2015, 87, 3314–3320. [Google Scholar] [CrossRef]
- Yamanaka, E.S.; Tortajada-Genaro, L.A.; Pastor, N.; Maquieira, A. Polymorphism genotyping based on loop-mediated isothermal amplification and smartphone detection. Biosens. Bioelectron. 2018, 109, 177–183. [Google Scholar] [CrossRef]
- Badolo, A.; Okado, K.; Guelbeogo, W.M.; Aonuma, H.; Bando, H.; Fukumoto, S.; Sagnon, N.; Kanuka, H. Development of an allele-specific, loop-mediated, isothermal amplification method (AS-LAMP) to detect the L1014F kdr-w mutation in Anopheles gambiae sl. Malar. J. 2012, 11, 227. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Chen, R.; Chen, G.; Li, M.; Wang, J.; Zou, J.; Du, F.; Dong, J.; Cui, X.; Huang, X.; et al. One-step colorimetric genotyping of single nucleotide polymorphism using probe-enhanced loop-mediated isothermal amplification (PE-LAMP). Theranostics 2019, 9, 3723–3731. [Google Scholar] [CrossRef] [PubMed]
- Yongkiettrakul, S.; Kampeera, J.; Chareanchim, W.; Rattanajak, R.; Pornthanakasem, W.; Kiatpathomchai, W.; Kongkasuriyachai, D. Simple detection of single nucleotide polymorphism in Plasmodium falciparum by SNP-LAMP assay combined with lateral flow dipstick. Parasitol. Int. 2017, 66, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Itonaga, M.; Matsuzaki, I.; Warigaya, K.; Tamura, T.; Shimizu, Y.; Fujimoto, M.; Fumiyoshi, K.; Masao, I.; Murata, S. Novel methodology for rapid detection of KRAS mutation using PNA-LNA mediated loop-mediated isothermal amplification. PLoS ONE 2016, 11, e0151654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, D.M.; Matthews, S.A.; Yan, G.; Zhou, G.; Lee, M.C.; Sirichaisinthop, J.; Kiattibutr, K.; Fan, Q.; Li, P.; Sattabongkot, J.; et al. Microgeography and molecular epidemiology of malaria at the Thailand-Myanmar border in the malaria pre-elimination phase. Malar. J. 2015, 14, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M.; Kanako, O.; Liu, Q.; Zhou, M.; Kawamoto, F.; Wataya, Y.; Otani, S.; Yamaguchi, Y.; Tanabe, K. Identification of the four species of human malaria parasites by nested PCR that targets variant sequences in the small subunit rRNA gene. Parasitol. Int. 1997, 46, 91–95. [Google Scholar] [CrossRef]
- Biswas, S.; Escalante, A.; Chaiyaroj, S.; Angkasekwinai, P.; Lal, A.A. Prevalence of point mutations in the dihydrofolate reductase and dihydropteroate synthetase genes of Plasmodium falciparum isolates from India and Thailand: A molecular epidemiologic study. Trop. Med. Int. Health 2001, 5, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Sugaram, R.; Suwannasin, K.; Kunasol, C.; Mathema, V.B.; Day, N.J.; Sudathip, P.; Prempree, P.; Dondorp, A.M.; Imwong, M. Molecular characterization of Plasmodium falciparum antifolate resistance markers in Thailand between 2008 and 2016. Malar. J. 2020, 19, 107. [Google Scholar] [CrossRef] [Green Version]
- Wongsrichanalai, C.; Baracus, M.J.; Muth, S.; Sutamihardja, A.; Wernsdorfer, W.H. A review of malaria diagnostic tools: Microscopy and rapid diagnostic test (RDT). In Defining and Defeating the Intolerable Burden of Malaria III: Progress and Perspectives; Supplement to Volume 77(6) of AJTMH; Breman, J.G., Alilio, M.S., White, N.J., Eds.; American Society of Tropical Medicine and Hygiene: Northbrook, IL, USA, 2007. [Google Scholar]
- WHO. Policy Brief on Malaria Diagnostics in Low-Transmission Settings; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Wu, L.; Van den Hoogen, L.L.; Slater, H.; Walker, P.G.T.; Ghani, A.C.; Drakeley, C.J.; Okell, L.C. Comparison of diagnostics for the detection of asymptomatic Plasmodium falciparum infections to inform control and elimination strategies. Nature 2015, 528, S86–S93. [Google Scholar] [CrossRef] [Green Version]
- Sattabongkot, J.; Suansomjit, C.; Nguitragool, W.; Sirichaisinthop, J.; Warit, S.; Tiensuwan, M.; Buates, S. Prevalence of asymptomatic Plasmodium infections with sub-microscopic parasite densities in the northwestern border of Thailand: A potential threat to malaria elimination. Malar. J. 2018, 17, 329. [Google Scholar] [CrossRef] [Green Version]
- Chahar, M.; Anvikar, A.; Valecha, N. Development and evaluation of a novel HNB based isothermal amplification assay for fast detection of Pyrimethamine resistance (S108N) in Plasmodium falciparum. Int. J. Environ. Res. Public Health 2019, 16, 1635. [Google Scholar] [CrossRef] [Green Version]
- Mohon, A.N.; Menard, D.; Alam, M.S.; Perera, K.; Pillai, D.R. A novel single-nucleotide polymorphism Loop Mediated Isothermal Amplification assay for detection of Artemisinin-resistant Plasmodium falciparum malaria. Open Forum Infect. Dis. 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Samples | (P %) | Parasite Density (P/µL) 1 | PfSNP-LAMP-LFD (N51I) | |
|---|---|---|---|---|
| P. falciparum2 | 2 | ND | ND | 2 |
| 5 | >0.2 | >10,000 | 5 | |
| 25 | >0.02–0.2 | >1000–10,000 | 25 | |
| 10 | >0.004–0.02 | >200–1000 | 10 | |
| 6 | >0.0002–0.004 | >100–200 | 6 | |
| 3 | 0.0001–0.0002 | >50–100 | 3 | |
| 4 | <0.0001 | <50 | 4 | |
| P. vivax3 | 1 | ND | ND | 0 |
| 72 | <0.0002–0.24 | <10–12,000 | 0 | |
| TOTAL | 128 | 55 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yongkiettrakul, S.; Kolié, F.R.; Kongkasuriyachai, D.; Sattabongkot, J.; Nguitragool, W.; Nawattanapaibool, N.; Suansomjit, C.; Warit, S.; Kangwanrangsan, N.; Buates, S. Validation of PfSNP-LAMP-Lateral Flow Dipstick for Detection of Single Nucleotide Polymorphism Associated with Pyrimethamine Resistance in Plasmodium falciparum. Diagnostics 2020, 10, 948. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10110948
Yongkiettrakul S, Kolié FR, Kongkasuriyachai D, Sattabongkot J, Nguitragool W, Nawattanapaibool N, Suansomjit C, Warit S, Kangwanrangsan N, Buates S. Validation of PfSNP-LAMP-Lateral Flow Dipstick for Detection of Single Nucleotide Polymorphism Associated with Pyrimethamine Resistance in Plasmodium falciparum. Diagnostics. 2020; 10(11):948. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10110948
Chicago/Turabian StyleYongkiettrakul, Suganya, Fassou René Kolié, Darin Kongkasuriyachai, Jetsumon Sattabongkot, Wang Nguitragool, Namfon Nawattanapaibool, Chayanut Suansomjit, Saradee Warit, Niwat Kangwanrangsan, and Sureemas Buates. 2020. "Validation of PfSNP-LAMP-Lateral Flow Dipstick for Detection of Single Nucleotide Polymorphism Associated with Pyrimethamine Resistance in Plasmodium falciparum" Diagnostics 10, no. 11: 948. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10110948