Growth Plate Pathology in the Mucopolysaccharidosis Type VI Rat Model—An Experimental and Computational Approach

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of Growth Plate Pathology in the MPS VI Rat Model
2.1.1. Animals
2.1.2. Sample Collection
2.1.3. Histological Analyses
2.1.4. Quantitative Analysis of Growth Plates
2.1.5. Statistical Analysis
2.2. In Silico Analysis
3. Results
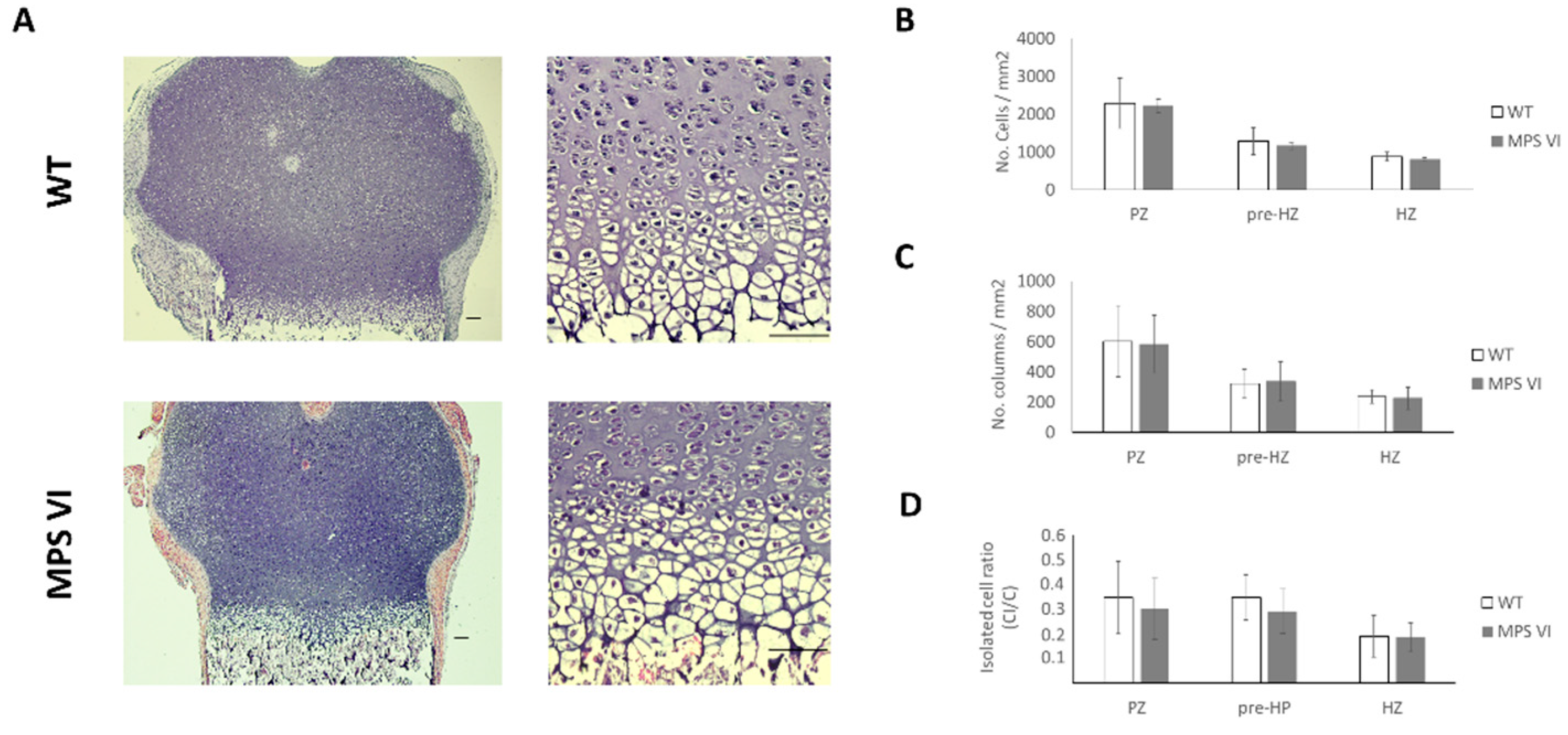
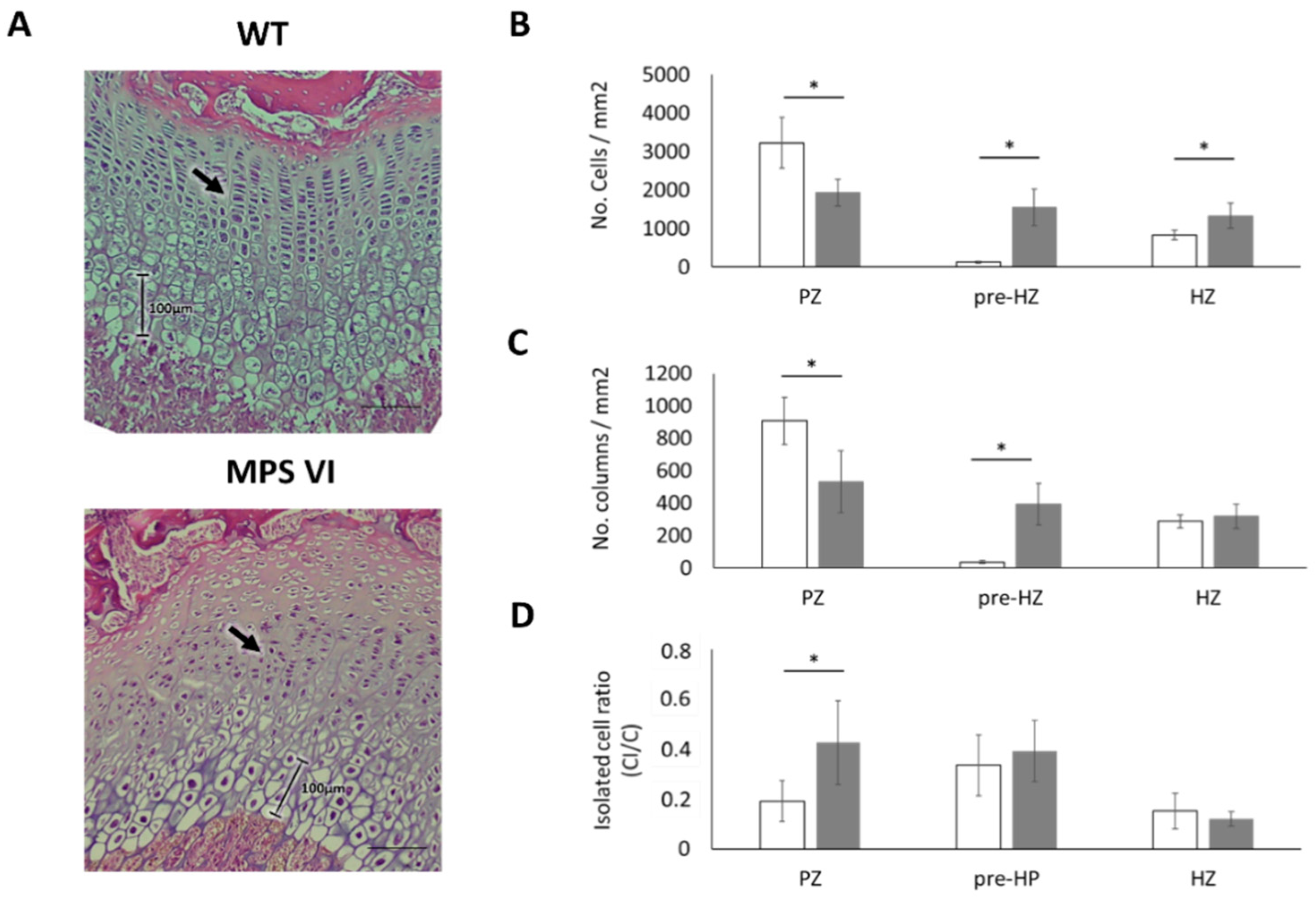
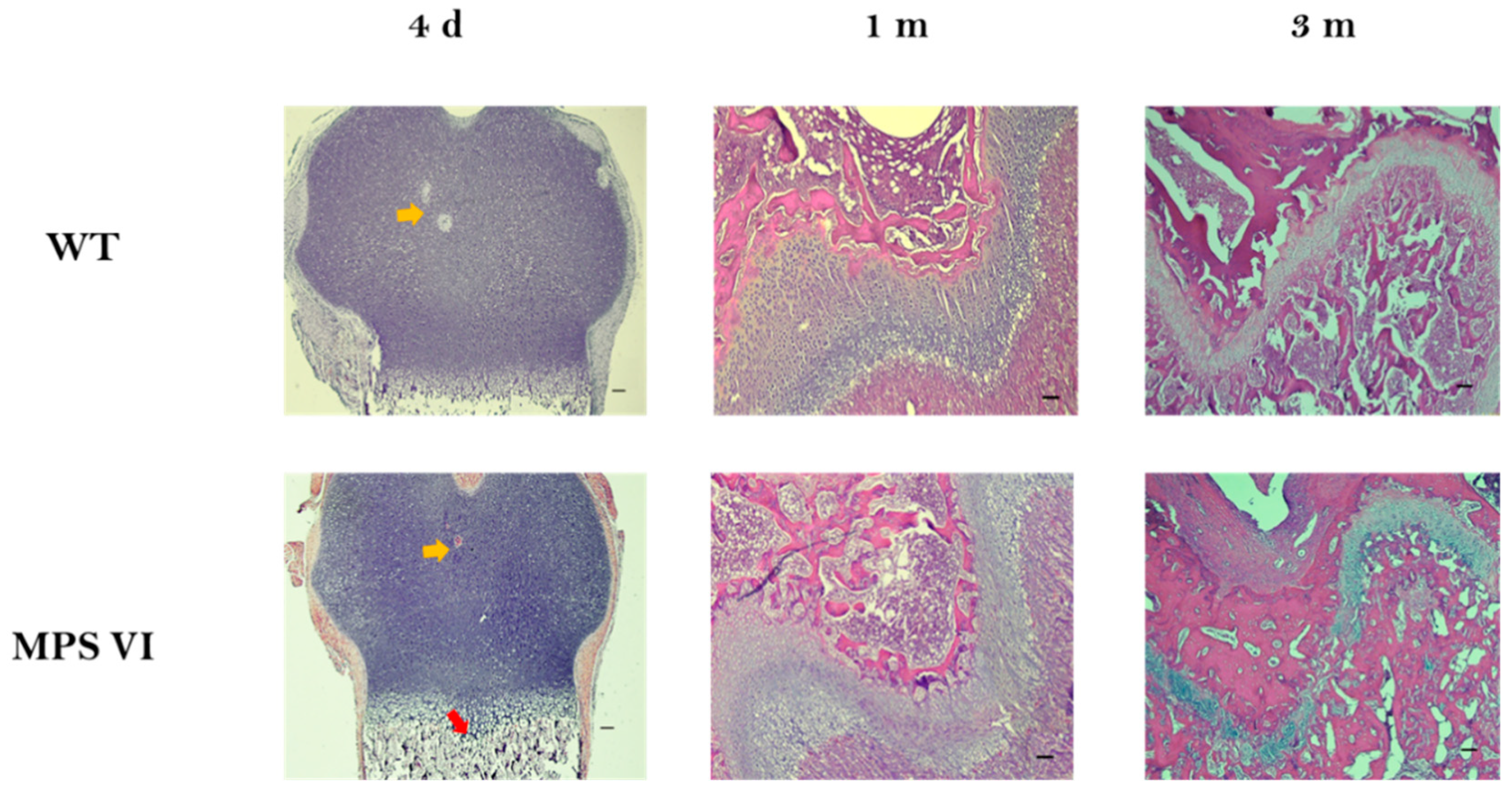
3.1. Histological Description
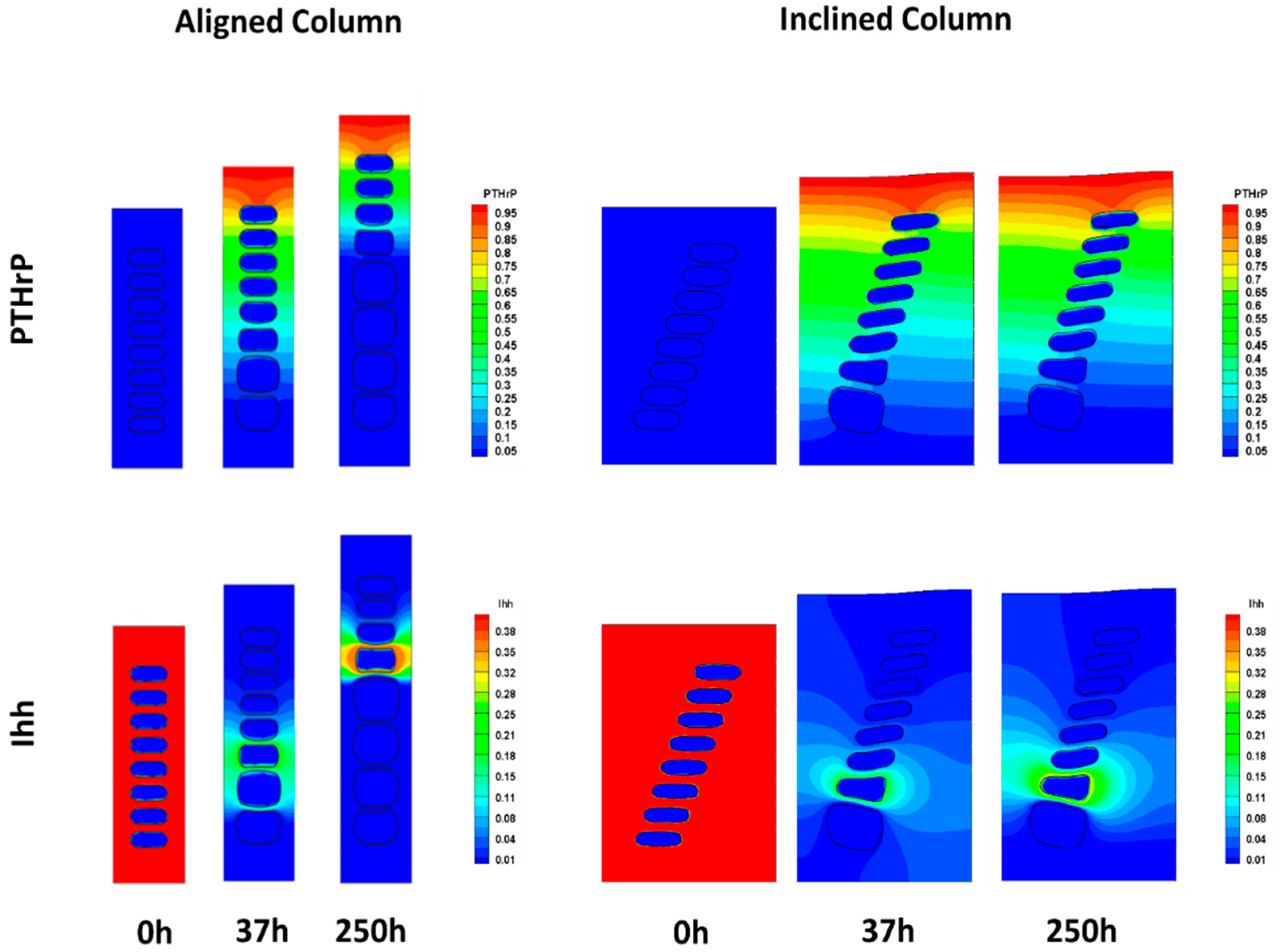
3.2. In Silico Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MPS | Mucopolysaccharidoses |
| WT | Wild type |
| MPS VI | Mucopolysaccharidosis type VI |
| MPS VII | Mucopolysaccharidosis type VII |
| DS | Dermatan sulphate |
| C4S | Chondroitin 4 sulphate |
| PCM | Pericellular matrix |
| ECM | Extracellular matrix |
| GP | Growth plate |
| RPZ | Resting + proliferative zone |
| PZ | Proliferative zone |
| Pre-HZ | Pre-hypertrophic zone |
| HZ | Hypertrophic zone |
| PTHrP | Parathiroid related protein |
| Ihh | Indian hedgehog |
| CI | Isolated cell density |
| C | Cellular density |
| CD | Column density |
| RT | Room temperature |
| GAG | Glycosaminoglycan |
| Col II | Collagen type II |
References
- Neufeld, E.F.; Muenzer, J. The Mucopolysaccharidoses. In The Online Metabolic and Molecular Bases of Inherited Disease; Valle, D., Vogelstain, B., Kinzler, K., Antonarakis, S., Ballabio, A., Mitchell, G.A., Brunetti-Pierri, N., Scriver, C.R., Sly, W.S., Beaudet, A., Eds.; McGraw-Hill: New York, NY, USA, 2006; Available online: http://www.ommbid.com (accessed on 20 January 2019).
- Morishita, K.; Petty, R.E. Musculoskeletal manifestations of mucopolysaccharidoses. Rheumatology (Oxf.) 2011, 50 (Suppl. 5), v19–v25. [Google Scholar] [CrossRef] [Green Version]
- Oussoren, E.; Brands, M.M.; Ruijter, G.J.; der Ploeg, A.T.; Reuser, A.J. Bone, joint and tooth development in mucopolysaccharidoses: Relevance to therapeutic options. Biochim. Biophys. Acta 2011, 1812, 1542–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burdan, F.; Szumilo, J.; Korobowicz, A.; Farooquee, R.; Patel, S.; Patel, A.; Dave, A.; Szumilo, M.; Solecki, M.; Klepacz, R.; et al. Morphology and physiology of the epiphyseal growth plate. Folia Histochem. Cytobiol. 2009, 47, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, A.W.; Cormack, D.H. Ham’s Histology, 9th ed.; Lippincott: Philadelphia, PA, USA, 1987. [Google Scholar]
- Mackie, E.J.; Ahmed, Y.A.; Tatarczuch, L.; Chen, K.S.; Mirams, M. Endochondral ossification: How cartilage is converted into bone in the developing skeleton. Int. J. Biochem. Cell Biol. 2008, 40, 46–62. [Google Scholar] [CrossRef] [PubMed]
- Mackie, E.J.; Tatarczuch, L.; Mirams, M. The skeleton: A multi-functional complex organ: The growth plate chondrocyte and endochondral ossification. J. Endocrinol. 2011, 211, 109–121. [Google Scholar] [CrossRef]
- Ballock, R.T.; O’Keefe, R.J. The biology of the growth plate. J. Bone Joint Surg. Am. 2003, 85-A, 715–726. [Google Scholar] [CrossRef]
- Anderson, C.E.; Crane, J.T.; Harper, H.A.; Hunter, T.W. Morquio’s disease and dysplasia epiphysalis multiplex. A study of epiphyseal cartilage in seven cases. J. Bone Joint Surg. Am. 1962, 44-A, 295–306. [Google Scholar] [CrossRef]
- McClure, J.; Smith, P.S.; Sorby-Adams, G.; Hopwood, J. The histological and ultrastructural features of the epiphyseal plate in Morquio type A syndrome (mucopolysaccharidosis type IVA). Pathology 1986, 18, 217–221. [Google Scholar] [CrossRef]
- Metcalf, J.A.; Zhang, Y.; Hilton, M.J.; Long, F.; Ponder, K.P. Mechanism of shortened bones in mucopolysaccharidosis VII. Mol. Genet. Metab. 2009, 97, 202–211. [Google Scholar] [CrossRef]
- Alliston, T. Chondroitin sulfate and growth factor signaling in the skeleton: Possible links to MPS VI. J. Pediatr. Rehabil. Med. 2010, 3, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Abreu, S.; Hayden, J.; Berthold, P.; Shapiro, I.M.; Decker, S.; Patterson, D.; Haskins, M. Growth plate pathology in feline mucopolysaccharidosis VI. Calcif. Tissue Int. 1995, 57, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Nuttall, J.D.; Brumfield, L.K.; Fazzalari, N.L.; Hopwood, J.J.; Byers, S. Histomorphometric analysis of the tibial growth plate in a feline model of mucopolysaccharidosis type VI. Calcif. Tissue Int. 1999, 65, 47–52. [Google Scholar] [CrossRef]
- Yoshida, M.; Noguchi, J.; Ikadai, H.; Takahashi, M.; Nagase, S. Arylsulfatase B-deficient mucopolysaccharidosis in rats. J. Clin. Investig. 1993, 91, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, L.; Roseti, L.; Desando, G.; Facchini, A.; Grigolo, B. A molecular and histological characterization of cartilage from patients with Morquio syndrome. Osteoarthr. Cartil. 2007, 15, 1311–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valayannopoulos, V.; Nicely, H.; Harmatz, P.; Turbeville, S. Mucopolysaccharidosis VI. Orphanet J. Rare Dis. 2010, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sannaa, N.A.; Al-Abdulwahed, H.Y.; Al-Majed, S.I.; Bouholaigah, I.H. The clinical and genetic Spectrum of Maroteaux-Lamy syndrome (Mucopolysaccharidosis VI) in the Eastern Province of Saudi Arabia. J. Community Genet. 2018, 9, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskins, M.E. Animal models for mucopolysaccharidosis disorders and their clinical relevance. Acta Paediatr. 2007, 96, 56–62. [Google Scholar] [CrossRef]
- Frohbergh, M.; Ge, Y.; Meng, F.; Karabul, N.; Solyom, A.; Lai, A.; Iatridis, J.; Schuchman, E.H.; Simonaro, C.M. Dose responsive effects of subcutaneous pentosan polysulfate injection in mucopolysaccharidosis type VI rats and comparison to oral treatment. PLoS ONE 2014, 9, e100882. [Google Scholar] [CrossRef] [Green Version]
- Simonaro, C.M.; Haskins, M.E.; Schuchman, E.H. Articular chondrocytes from animals with a dermatan sulfate storage disease undergo a high rate of apoptosis and release nitric oxide and inflammatory cytokines: A possible mechanism underlying degenerative joint disease in the mucopolysaccharidoses. Lab. Investig. 2001, 81, 1319–1328. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, M.; Ikadai, H.; Maekawa, A.; Takahashi, M.; Nagase, S. Pathological characteristics of mucopolysaccharidosis VI in the rat. J. Comp. Pathol. 1993, 109, 141–153. [Google Scholar] [CrossRef]
- Quantifying Stained Liver Tissue. Available online: https://imagej.nih.gov/ij/docs/examples/stained-sections/index.html (accessed on 12 January 2020).
- Valteau, B.; Grimard, G.; Londono, I.; Moldovan, F.; Villemure, I. In vivo dynamic bone growth modulation is less detrimental but as effective as static growth modulation. Bone 2011, 49, 996–1004. [Google Scholar] [CrossRef]
- Guevara, J.M.; Castro-Abril, H.A.; Barrera, L.A.; Garzón-Alvarado, D.A. A quantitative and qualitative growth plate description: A simple framework for chondrocytes columnar arrangement evaluation. J. Mech. Med. Biol. 2016, 16, 1650054. [Google Scholar] [CrossRef]
- Castro-Abril, H.A.; Guevara, J.M.; Moncayo, M.A.; Shefelbine, S.J.; Barrera, L.A.; Garzón-Alvarado, D.A. Cellular scale model of growth plate: An in silico model of chondrocyte hypertrophy. J. Theor. Biol. 2017, 428, 87–97. [Google Scholar] [CrossRef]
- Karsenty, G.; Kronenberg, H.M.; Settembre, C. Genetic control of bone formation. Annu. Rev. Cell Dev. Biol. 2009, 25, 629–648. [Google Scholar] [CrossRef]
- Kronenberg, H.M. Developmental regulation of the growth plate. Nature 2003, 423, 332–336. [Google Scholar] [CrossRef]
- Provot, S.; Schipani, E. Molecular mechanisms of endochondral bone development. Biochem. Biophys. Res. Commun. 2005, 328, 658–665. [Google Scholar] [CrossRef]
- Chau, M.; Forcinito, P.; Andrade, A.C.; Hegde, A.; Ahn, S.; Lui, J.C.; Baron, J.; Nilsson, O. Organization of the Indian hedgehog-parathyroid hormone-related protein system in the postnatal growth plate. J. Mol. Endocrinol. 2011, 47, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Tryfonidou, M.A.; Hazewinkel, H.A.; Riemers, F.M.; Brinkhof, B.; Penning, L.C.; Karperien, M. Intraspecies disparity in growth rate is associated with differences in expression of local growth plate regulators. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E1044–E1052. [Google Scholar] [CrossRef] [Green Version]
- Giugliani, R.; Federhen, A.; Rojas, M.V.; Vieira, T.; Artigalas, O.; Pinto, L.L.; Azevedo, A.C.; Acosta, A.; Bonfim, C.; Lourenco, C.M.; et al. Mucopolysaccharidosis I, II, and VI: Brief review and guidelines for treatment. Genet. Mol. Biol. 2010, 33, 589–604. [Google Scholar] [CrossRef]
- Jiang, Z.; Derrick-Roberts, A.L.K.; Jackson, M.R.; Rossouw, C.; Pyragius, C.E.; Xian, C.; Fletcher, J.; Byers, S. Delayed development of ossification centers in the tibia of prenatal and early postnatal MPS VII mice. Mol. Genet. Metab. 2018, 124, 135–142. [Google Scholar] [CrossRef]
- Peck, S.H.; O’Donnell, P.J.; Kang, J.L.; Malhotra, N.R.; Dodge, G.R.; Pacifici, M.; Shore, E.M.; Haskins, M.E.; Smith, L.J. Delayed hypertrophic differentiation of epiphyseal chondrocytes contributes to failed secondary ossification in mucopolysaccharidosis VII dogs. Mol. Genet. Metab. 2015, 116, 195–203. [Google Scholar] [CrossRef] [Green Version]
- Eliyahu, E.; Wolfson, T.; Ge, Y.; Jepsen, K.J.; Schuchman, E.H.; Simonaro, C.M. Anti-TNF-alpha therapy enhances the effects of enzyme replacement therapy in rats with mucopolysaccharidosis type VI. PLoS ONE 2011, 6, e22447. [Google Scholar] [CrossRef] [Green Version]
- Kamisan, N.; Naveen, S.V.; Ahmad, R.E.; Kamarul, T. Chondrocyte density, proteoglycan content and gene expressions from native cartilage are species specific and not dependent on cartilage thickness: A comparative analysis between rat, rabbit and goat. BMC Vet. Res. 2013, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Butler, W.F.; Wels, C.M. Glycosaminoglycans of cat intervertebral disc. Biochem. J. 1971, 122, 647–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevara, J.M.; Moncayo, M.A.; Vaca-Gonzalez, J.J.; Gutierrez, M.L.; Barrera, L.A.; Garzon-Alvarado, D.A. Growth plate stress distribution implications during bone development: A simple framework computational approach. Comput. Methods Programs Biomed. 2015, 118, 59–68. [Google Scholar] [CrossRef]
- Piszczatowski, S. Geometrical aspects of growth plate modelling using Carter’s and Stokes’s approaches. Acta Bioeng. Biomech. 2012, 14, 93–106. [Google Scholar]
- De Oliveira, P.G.; Baldo, G.; Mayer, F.Q.; Martinelli, B.; Meurer, L.; Giugliani, R.; Matte, U.; Xavier, R.M. Characterization of joint disease in mucopolysaccharidosis type I mice. Int. J. Exp. Pathol. 2013, 94, 305–311. [Google Scholar] [CrossRef]
- Haskins, M.; Casal, M.; Ellinwood, N.M.; Melniczek, J.; Mazrier, H.; Giger, U. Animal models for mucopolysaccharidoses and their clinical relevance. Acta Paediatr. Suppl. 2002, 91, 88–97. [Google Scholar] [CrossRef]
- Simonaro, C.M.; Ge, Y.; Eliyahu, E.; He, X.; Jepsen, K.J.; Schuchman, E.H. Involvement of the Toll-like receptor 4 pathway and use of TNF-alpha antagonists for treatment of the mucopolysaccharidoses. Proc. Natl. Acad. Sci. USA 2010, 107, 222–227. [Google Scholar] [CrossRef] [Green Version]
- Tomatsu, S.; Orii, K.O.; Vogler, C.; Grubb, J.H.; Snella, E.M.; Gutierrez, M.; Dieter, T.; Holden, C.C.; Sukegawa, K.; Orii, T.; et al. Production of MPS VII mouse (Gus(tm(hE540A x mE536A)Sly)) doubly tolerant to human and mouse beta-glucuronidase. Hum. Mol. Genet. 2003, 12, 961–973. [Google Scholar] [CrossRef] [Green Version]
- Alini, M.; Roughley, P.J. Changes in leucine-rich repeat proteoglycans during maturation of the bovine growth plate. Matrix Biol. 2001, 19, 805–813. [Google Scholar] [CrossRef]
- Martel-Pelletier, J.; Boileau, C.; Pelletier, J.P.; Roughley, P.J. Cartilage in normal and osteoarthritis conditions. Best Pract. Res. Clin. Rheumatol. 2008, 22, 351–384. [Google Scholar] [CrossRef] [PubMed]
- Gentili, C.; Cancedda, R. Cartilage and bone extracellular matrix. Curr. Pharm. Des. 2009, 15, 1334–1348. [Google Scholar] [CrossRef] [PubMed]
- Ruhland, C.; Schonherr, E.; Robenek, H.; Hansen, U.; Iozzo, R.V.; Bruckner, P.; Seidler, D.G. The glycosaminoglycan chain of decorin plays an important role in collagen fibril formation at the early stages of fibrillogenesis. FEBS J. 2007, 274, 4246–4255. [Google Scholar] [CrossRef]
- Nikitovic, D.; Aggelidakis, J.; Young, M.F.; Iozzo, R.V.; Karamanos, N.K.; Tzanakakis, G.N. The biology of small leucine-rich proteoglycans in bone pathophysiology. J. Biol. Chem. 2012, 287, 33926–33933. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, S.; Hosaka, Y.; Iwasaki, T.; Yamamoto, K.; Nagayasu, A.; Ueda, H.; Kokai, Y.; Takehana, K. The modulation of collagen fibril assembly and its structure by decorin: An electron microscopic study. Arch. Histol. Cytol. 2008, 71, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Ezura, Y.; Chervoneva, I.; Robinson, P.S.; Beason, D.P.; Carine, E.T.; Soslowsky, L.J.; Iozzo, R.V.; Birk, D.E. Decorin regulates assembly of collagen fibrils and acquisition of biomechanical properties during tendon development. J. Cell. Biochem. 2006, 98, 1436–1449. [Google Scholar] [CrossRef]
- Olsen, B.R. Role of cartilage collagens in formation of the skeleton. Ann. N. Y. Acad. Sci. 1996, 785, 124–130. [Google Scholar] [CrossRef]
- Gustafsson, E.; Aszodi, A.; Ortega, N.; Hunziker, E.B.; Denker, H.W.; Werb, Z.; Fassler, R. Role of collagen type II and perlecan in skeletal development. Ann. N. Y. Acad. Sci. 2003, 995, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Arita, M.; Fertala, J.; Hou, C.; Steplewski, A.; Fertala, A. Mechanisms of Aberrant Organization of Growth Plates in Conditional Transgenic Mouse Model of Spondyloepiphyseal Dysplasia Associated with the R992C Substitution in Collagen II. Am. J. Pathol. 2015, 185, 214–229. [Google Scholar] [CrossRef]
- Myllyharju, J. Extracellular matrix and developing growth plate. Curr. Osteoporos. Rep. 2014, 12, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Thery, M.; Racine, V.; Pepin, A.; Piel, M.; Chen, Y.; Sibarita, J.B.; Bornens, M. The extracellular matrix guides the orientation of the cell division axis. Nat. Cell Biol. 2005, 7, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Amini, S.; Mortazavi, F.; Sun, J.; Levesque, M.; Hoemann, C.D.; Villemure, I. Stress relaxation of swine growth plate in semi-confined compression: Depth dependent tissue deformational behavior versus extracellular matrix composition and collagen fiber organization. Biomech. Model. Mechanobiol. 2013, 12, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Cowin, S.C.; Doty, S.B. Cartilage. In Tissue Mechanics; Springer: Berlin, Germany, 2007; pp. 471–505. [Google Scholar]
- Smith, L.J.; Martin, J.T.; Szczesny, S.E.; Ponder, K.P.; Haskins, M.E.; Elliott, D.M. Altered lumbar spine structure, biochemistry, and biomechanical properties in a canine model of mucopolysaccharidosis type VII. J. Orthop. Res. 2010, 28, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.; Simonaro, C.M.; Schuchman, E.H.; Ge, Y.; Laudier, D.M.; Iatridis, J.C. Structural, compositional, and biomechanical alterations of the lumbar spine in rats with mucopolysaccharidosis type VI (Maroteaux-Lamy syndrome). J. Orthop. Res. 2013, 31, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Stokes, I.A. Mechanical effects on skeletal growth. J. Musculoskelet. Neuronal. Interact. 2002, 2, 277–280. [Google Scholar] [PubMed]
- Villemure, I.; Stokes, I.A. Growth plate mechanics and mechanobiology. A survey of present understanding. J. Biomech. 2009, 42, 1793–1803. [Google Scholar] [CrossRef] [Green Version]
- Vaca-Gonzalez, J.J.; Guevara, J.M.; Moncayo, M.A.; Castro-Abril, H.; Hata, Y.; Garzon-Alvarado, D.A. Biophysical Stimuli: A Review of Electrical and Mechanical Stimulation in Hyaline Cartilage. Cartilage 2019, 10, 157–172. [Google Scholar] [CrossRef]
- Platt, F.M.; Boland, B.; van der Spoel, A.C. The cell biology of disease: Lysosomal storage disorders: The cellular impact of lysosomal dysfunction. J. Cell Biol. 2012, 199, 723–734. [Google Scholar] [CrossRef] [Green Version]
- Ballabio, A.; Gieselmann, V. Lysosomal disorders: From storage to cellular damage. Biochim. Biophys. Acta 2009, 1793, 684–696. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Parameter | Wild Type | MPS VI |
|---|---|---|---|
| 4 days | Growth plate thickness (µm) | 659.04 ± 96.28 | 539.5 ± 136.26 |
| Resting-proliferative zone thickness (µm) | 461.83 ± 87.24 | 319.15 ± 118.35 | |
| Hypertrophic zone thickness (µm) | 197.11 ± 29.74 | 220.35 ± 33.02 | |
| 1 month | Growth plate thickness (µm) | 466.62 ± 46.16 | 381.68 ± 81.23 |
| Resting-proliferative zone thickness (µm) | 225.17 ± 34.63 | 241.24 ± 59.75 | |
| Hypertrophic zone thickness (µm) | 241.11 ± 31.61 | 140.43 ± 41.74 * | |
| 3 months | Growth plate thickness (µm) | 224.49 ± 33.98 | 244 ± 78.4 |
| Age | Growth Plate Zone | Column Orientation Angle (α) *,§ | Variation Coefficient of α (%) * | ||
|---|---|---|---|---|---|
| Wild Type | MPS VI | Wild Type | MPS VI | ||
| 4 days | Proliferative | 86.77 | 89.38 | 33 | 28 |
| Pre-hypertrophic | 88.53 | 85.99 | 28 | 39 | |
| Hypertrophic | 90.24 | 91.18 | 22 | 22 | |
| 1 month | Proliferative | 93.82 | 84.05 | 15 | 20 |
| Pre-hypertrophic | 92.15 | 82.48 | 17 | 23 | |
| Hypertrophic | 89.4 | 81.51 | 13 | 21 | |
| 3 months | Total growth plate | 92.23 | 86.69 | 17 | 24 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guevara-Morales, J.M.; Frohbergh, M.; Castro-Abril, H.; Vaca-González, J.J.; Barrera, L.A.; Garzón-Alvarado, D.A.; Schuchman, E.; Simonaro, C. Growth Plate Pathology in the Mucopolysaccharidosis Type VI Rat Model—An Experimental and Computational Approach. Diagnostics 2020, 10, 360. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10060360
Guevara-Morales JM, Frohbergh M, Castro-Abril H, Vaca-González JJ, Barrera LA, Garzón-Alvarado DA, Schuchman E, Simonaro C. Growth Plate Pathology in the Mucopolysaccharidosis Type VI Rat Model—An Experimental and Computational Approach. Diagnostics. 2020; 10(6):360. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10060360
Chicago/Turabian StyleGuevara-Morales, Johana M., Michael Frohbergh, Hector Castro-Abril, Juan J. Vaca-González, Luis A. Barrera, Diego A. Garzón-Alvarado, Edward Schuchman, and Calogera Simonaro. 2020. "Growth Plate Pathology in the Mucopolysaccharidosis Type VI Rat Model—An Experimental and Computational Approach" Diagnostics 10, no. 6: 360. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics10060360