
Importance of Potential New Biomarkers in Patient with Serouse Ovarian Cancer

 , and
, and
Abstract
:1. Introduction
Aim of the Study
2. Materials and Methods
2.1. Tissue Specimens
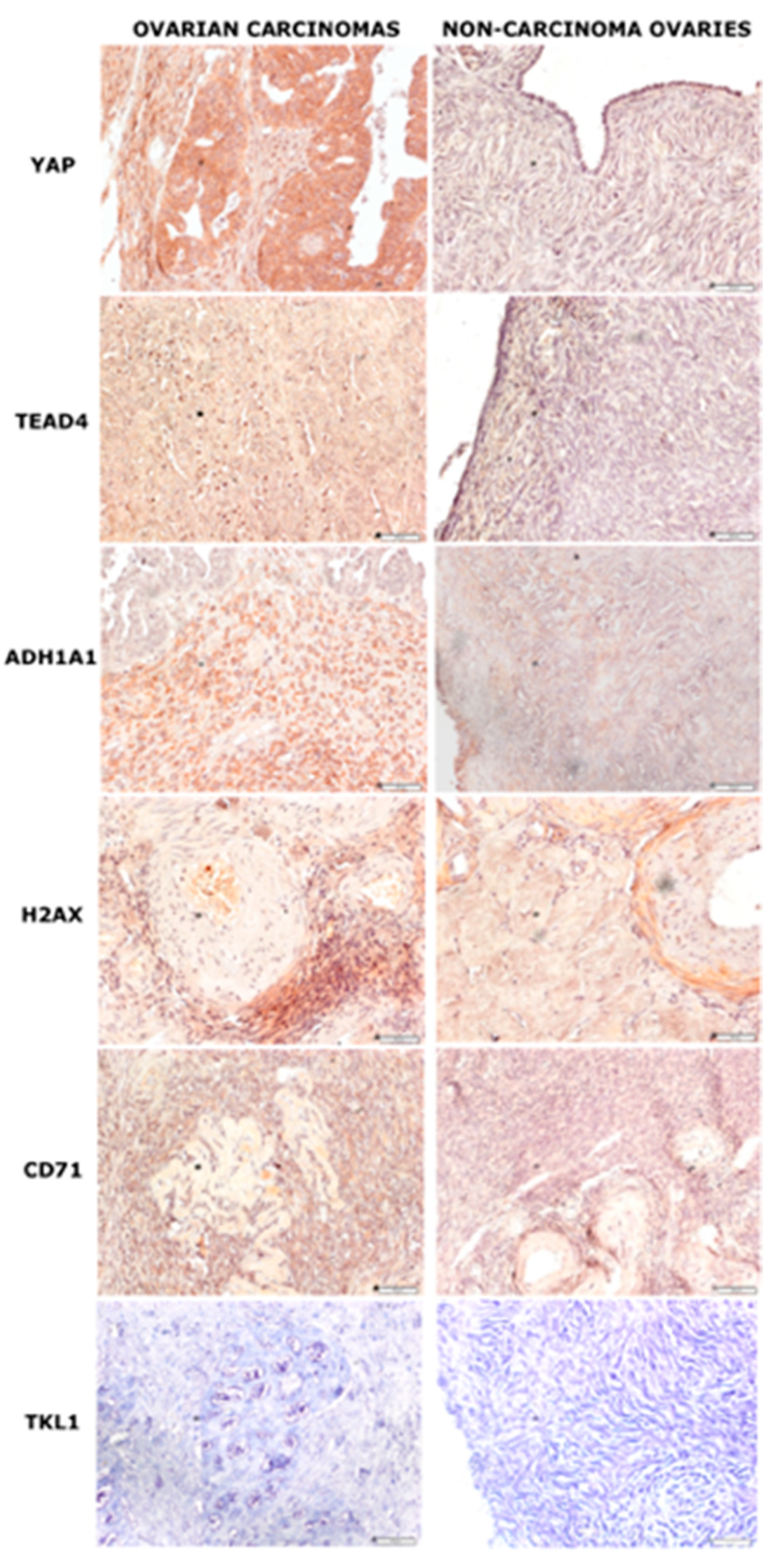
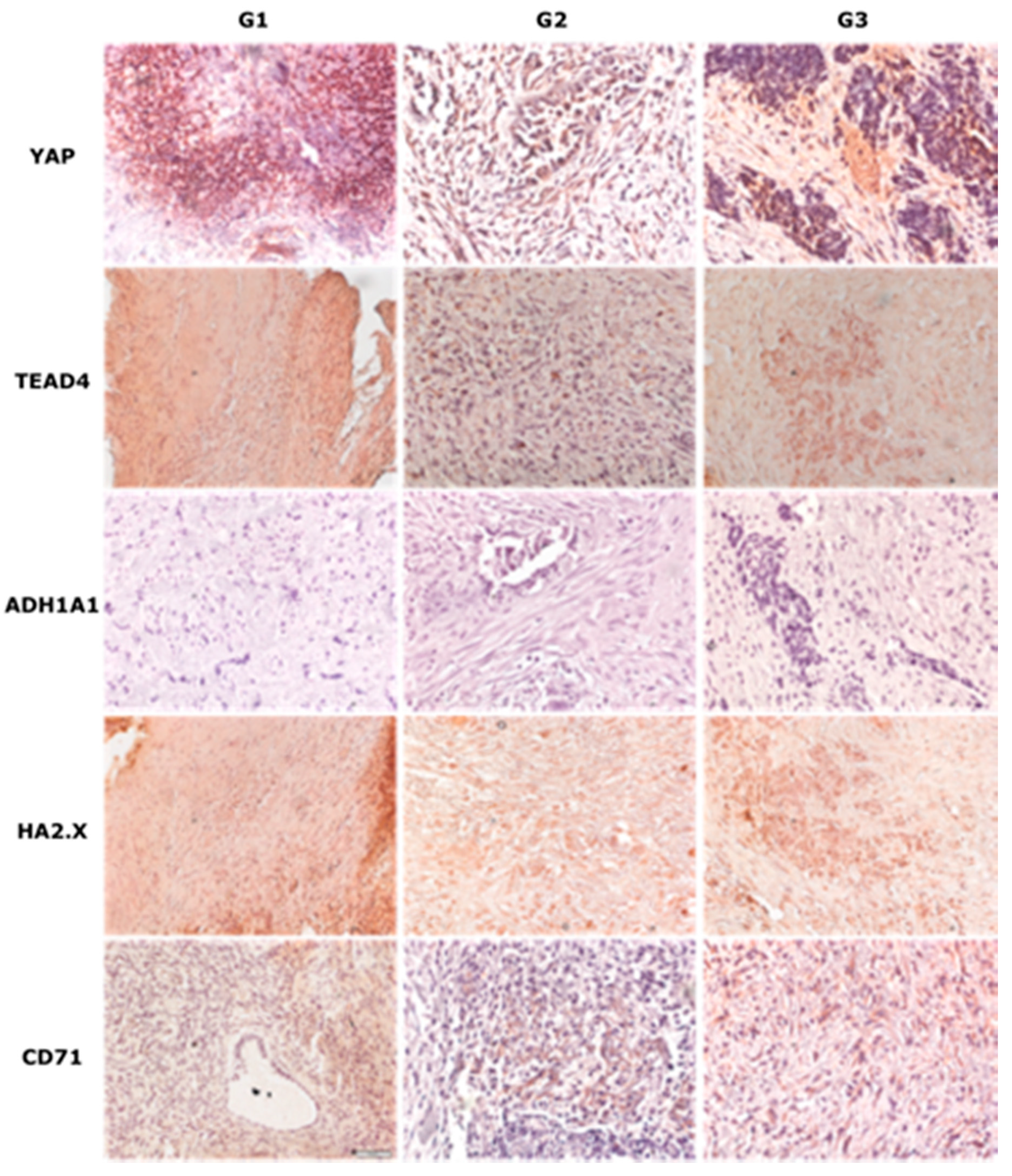
2.2. Immunohistochemical Analysis of Chosen Markers
2.3. Quantitative Real Time Polymerase Chain Reaction (qRT-PCR)
2.4. Statistical Analysis
3. Results
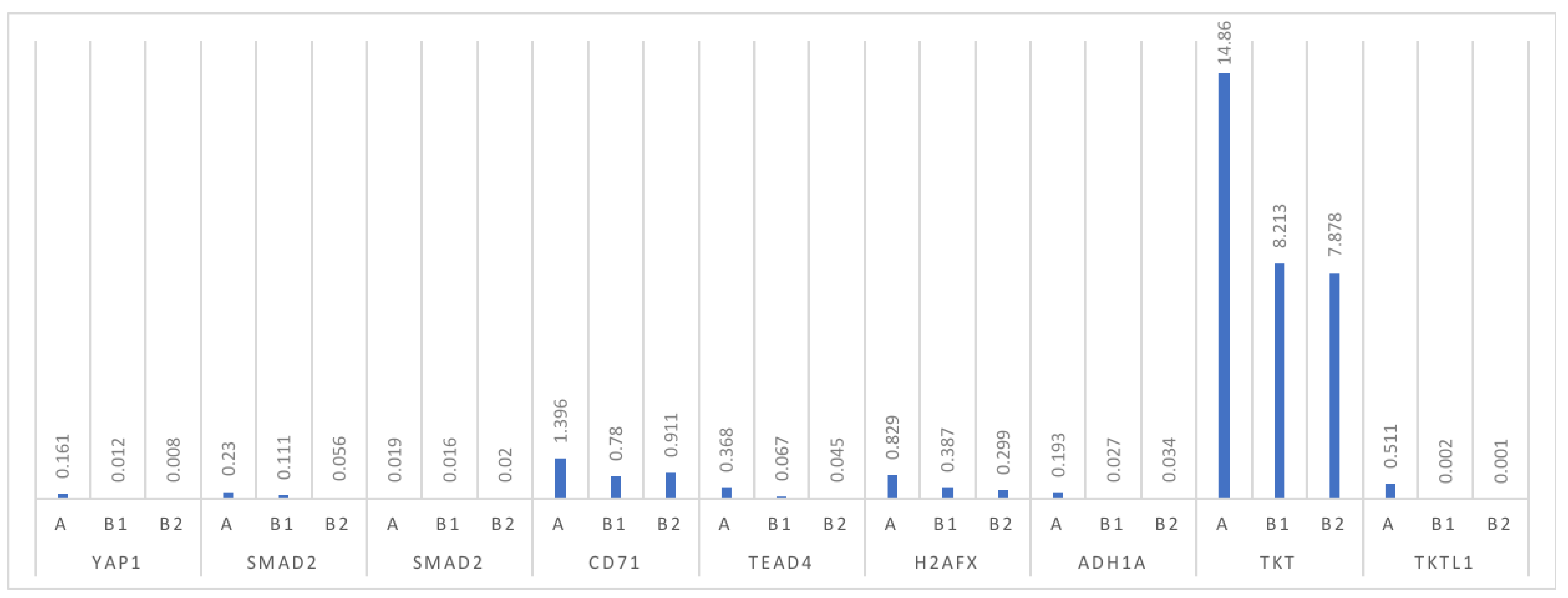
3.1. Analysis of mRNA Expression for Individual Proteins
3.2. Proteins as a Prognostic Factors
4. Discussion
4.1. Selected Factors Inducing DNA Damage
4.2. The Effect of Transferrin
4.3. Effect of Alcohol
4.4. Transketolase Family
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Liu, X.; Ren, X.; Tian, Y.; Chen, Z.; Xu, X.; Du, Y.; Jiang, C.; Fang, Y.; Liu, Z.; et al. Smad2 and Smad3 have differential sensitivity in relaying TGFβ signal-ing and inversely regulate early lineage specification. Sci. Rep. 2016, 6, 21602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roane, B.M.; Arend, R.C.; Birrer, M.J. Review: Targeting the Transforming Growth Factor-Beta Pathway in Ovarian Cancer. Cancers 2019, 11, 668. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, Y.; Zeng, Z.; Qiao, L.; Zhuang, L.; Gao, Q.; Ma, D.; Huang, X. Smad4 deletion in blood vessel endothelial cells promotes ovarian cancer metastasis. Int. J. Oncol. 2017, 50, 1693–1700. [Google Scholar] [CrossRef] [Green Version]
- Reid, B.M.; Permuth, J.B.; Sellers, T.A. Epidemiology of ovarian cancer: A review. Cancer Biol. Med. 2017, 14, 9–32. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Chang, T.; Wang, Y.; Liu, Y.; Li, W.; Li, M.; Fan, H.-Y. YAP Promotes Ovarian Cancer Cell Tumorigenesis and Is Indicative of a Poor Prognosis for Ovarian Cancer Patients. PLoS ONE 2014, 9, e91770. [Google Scholar] [CrossRef]
- Xie, Q.; Chen, J.; Feng, H.; Peng, S.; Adams, U.; Bai, Y.; Huang, L.; Li, J.; Huang, J.; Meng, S.; et al. YAP/TEAD–Mediated Transcription Controls Cellular Senescence. Cancer Res. 2013, 73, 3615–3624. [Google Scholar] [CrossRef] [Green Version]
- Yeh, K.-T.; Chen, T.-H.; Yang, H.-W.; Chou, J.-L.; Chen, L.-Y.; Yeh, C.-M.; Chen, Y.-H.; Lin, R.-I.; Su, H.-Y.; Chen, G.C.-W.; et al. Aberrant TGFβ/SMAD4 signaling contributes to epigenetic silencing of a putative tumor suppressor, RunX1T1, in ovarian cancer. Epigenetics 2011, 6, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; AOCS Study Group; George, J.M.; Deb, S.; Degoutin, J.L.; Takano, E.A.; Fox, S.B.; Bowtell, D.D.L.; Harvey, K.F. The Hippo pathway transcriptional co-activator, YAP, is an ovarian cancer oncogene. Oncogene 2011, 30, 2810–2822. [Google Scholar] [CrossRef] [Green Version]
- Turinetto, V.; Giachino, C. Multiple facets of histone variant H2AX: A DNA double-strand-break marker with several bio-logical functions. Nucleic Acids Res. 2015, 43, 2489–2498. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Capetillo, O.; Lee, A.; Nussenzweig, M.; Nussenzweig, A. H2AX: The histone guardian of the genome. DNA Repair 2004, 3, 959–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelski, W.; Chrostek, L.; Markiewicz, W.; Szmitkowski, M. Activity of alcohol dehydrogenase (adh) isoenzymes and aldehyde dehydrogenase (ALDH) in the sera of patients with breast cancer. J. Clin. Lab. Anal. 2006, 20, 105–108. [Google Scholar] [CrossRef]
- Althobiti, M.; El Ansari, R.; Aleskandarany, M.; Joseph, C.; Toss, M.S.; Green, A.R.; Rakha, E.A. The prognostic significance of ALDH1A1 expression in early invasive breast cancer. Histopathology 2020, 77, 437–448. [Google Scholar] [CrossRef]
- Shen, Y.; Li, X.; Dong, D.; Zhang, B.; Xue, Y.; Shang, P. Transferrin receptor 1 in cancer: A new sight for cancer therapy. Am. J. Cancer Res. 2018, 8, 916–931. [Google Scholar] [PubMed]
- Chan, K.T.; Choi, M.Y.; Lai, K.K.; Tan, W.; Tung, L.N.; Lam, H.Y.; Tong, D.K.; Lee, N.P.; Law, S. Overexpression of transferrin receptor CD71 and its tumorigenic properties in esophageal squamous cell carcinoma. Oncol. Rep. 2014, 31, 1296–1304. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Xiang, C.; Zhong, F.; Liu, Y.; Dong, Q.; Li, K.; Shi, W.; Ding, C.; Qin, L.; He, F. Transketolase (TKT) activity and nuclear localization promote hepatocellular carcinoma in a metabolic and a non-metabolic manner. J. Exp. Clin. Cancer Res. 2019, 38, 1–21. [Google Scholar] [CrossRef]
- Xu, I.M.-J.; Lai, R.K.-H.; Lin, S.-H.; Tse, A.P.-W.; Chiu, D.K.-C.; Koh, H.-Y.; Law, C.-T.; Wong, C.-M.; Cai, Z.; Wong, C.C.-L.; et al. Transketolase counteracts oxidative stress to drive cancer development. Proc. Natl. Acad. Sci. USA 2016, 113, E725–E734. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.W.; Lim, C.J.; Chen, L.; Chong, Y.F.; Huang, C.; Song, H.; Hong, W. The hippo pathway in biological control and cancer development. J. Cell. Physiol. 2011, 226, 928–939. [Google Scholar] [CrossRef]
- Allegra, A.; Pioggia, G.; Innao, V.; Musolino, C.; Gangemi, S. New Insights into YES-Associated Protein Signaling Pathways in Hematological Malignancies: Diagnostic and Therapeutic Challenges. Cancers 2021, 13, 1981. [Google Scholar] [CrossRef]
- He, C.; Lv, X.; Hua, G.; Lele, S.M.; Remmenga, S.W.; Dong, J.; Davis, J.S.; Wang, C. YAP forms autocrine loops with the ERBB pathway to regulate ovarian cancer initiation and progression. Oncogene 2015, 34, 6040–6054. [Google Scholar] [CrossRef] [Green Version]
- Liu-Chittenden, Y.; Huang, B.; Shim, J.S.; Chen, Q.; Lee, S.-J.; Anders, R.A.; Liu, J.O.; Pan, D. Genetic and pharmacological disruption of the TEAD-YAP complex suppresses the oncogenic activity of YAP. Genes Dev. 2012, 26, 1300–1305. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Jia, Y.; Lou, H.; Ma, J.; Huang, Q.; Meng, Y.; Sun, C.; Yang, Z.; Li, X.; Xu, S.; et al. Targeting YAP suppresses ovarian cancer progression through regulation of the PI3K/Akt/mTOR pathway. Oncol. Rep. 2019, 42, 2768–2776. [Google Scholar] [CrossRef]
- Kim, C.-L.; Choi, S.-H.; Mo, J.-S. Role of the Hippo Pathway in Fibrosis and Cancer. Cells 2019, 8, 468. [Google Scholar] [CrossRef] [Green Version]
- Moroishi, T.; Hayashi, T.; Pan, W.-W.; Fujita, Y.; Holt, M.V.; Qin, J.; Carson, D.A.; Guan, K.-L. The Hippo Pathway Kinases LATS1/2 Suppress Cancer Immunity. Cell 2016, 167, 1525–1539.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, J.; Cao, B.; Wang, Y.; Ma, C.; Zeng, Z.; Liu, L.; Li, X.; Tao, D.; Gong, J.; Xie, D. Hippo component YAP promotes focal adhesion and tumour aggressiveness via transcriptionally activating THBS1/FAK signalling in breast cancer. J. Exp. Clin. Cancer Res. 2018, 37, 175. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Guo, J.; Wu, Y.; Jin, D.; Jiang, H.; Liu, C.; Qin, C. Suppression of YAP by DDP disrupts colon tumor progression. Oncol. Rep. 2018, 39, 2114–2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Feng, Y.; Hu, Y.; He, C.; Xie, C.; Ouyang, Y.; Artim, S.C.; Huang, D.; Zhu, Y.; Luo, Z.; et al. Helicobacter pylori CagA promotes epithelial mesenchymal transition in gastric carcinogenesis via triggering oncogenic YAP pathway. J. Exp. Clin. Cancer Res. 2018, 37, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Macias, M.J.; Martin-Malpartida, P.; Massagué, J. Structural determinants of Smad function in TGF-β signaling. Trends Biochem. Sci. 2015, 40, 296–308. [Google Scholar] [CrossRef] [Green Version]
- Pardali, E.; Dijke, P.T. TGFβ Signaling and Cardiovascular Diseases. Int. J. Biol. Sci. 2012, 8, 195–213. [Google Scholar] [CrossRef]
- Yu, W.; Chai, H. Inhibition of BAMBI reduces the viability and motility of colon cancer via activating TGF-β/Smad pathway in vitro and in vivo. Oncol. Lett. 2017, 14, 4793–4799. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, B.A.; Deatherage, D.E.; Gu, F.; Tang, B.; Chan, M.W.Y.; Nephew, K.P.; Huang, T.H.-M.; Jin, V.X. ChIP-seq Defined Genome-Wide Map of TGFβ/SMAD4 Targets: Implications with Clinical Outcome of Ovarian Cancer. PLoS ONE 2011, 6, e22606. [Google Scholar] [CrossRef] [Green Version]
- Wakahara, K.; Kobayashi, H.; Yagyu, T.; Matsuzaki, H.; Kondo, T.; Kurita, N.; Sekino, H.; Inagaki, K.; Suzuki, M.; Kanayama, N.; et al. Transforming growth factor-?1-dependent activation of Smad2/3 and up-regulation of PAI-1 expression is negatively regulated by Src in SKOV-3 human ovarian cancer cells. J. Cell. Biochem. 2004, 93, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Shigeta, S.; Toyoshima, M.; Kitatani, K.; Ishibashi, M.; Usui, T.; Yaegashi, N. Transferrin facilitates the formation of DNA double-strand breaks via transferrin receptor 1: The possible involvement of transferrin in carcinogenesis of high-grade serous ovarian cancer. Oncogene 2016, 35, 3577–3586. [Google Scholar] [CrossRef] [PubMed]
- Helt, C.E.; Cliby, W.A.; Keng, P.C.; Bambara, R.A.; O’Reilly, M.A. Ataxia Telangiectasia Mutated (ATM) and ATM and Rad3-related Protein Exhibit Selective Target Specificities in Response to Different Forms of DNA Damage. J. Biol. Chem. 2005, 280, 1186–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kastan, M.B. DNA Damage Responses: Mechanisms and Roles in Human Disease. Mol. Cancer Res. 2008, 6, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Opławski, M.; Nowakowski, R.; Średnicka, A.; Ochnik, D.; Grabarek, B.; Boroń, D. Molecular Landscape of the Epithelial–Mesenchymal Transition in Endometrioid Endometrial Cancer. J. Clin. Med. 2021, 10, 1520. [Google Scholar] [CrossRef]
- Tanaka, T.; Halicka, H.D.; Huang, X.; Traganos, F.; Darzynkiewicz, Z. Constitutive Histone H2AX Phosphorylation and ATM Activation, the Reporters of DNA Damage by Endogenous Oxidants. Cell Cycle 2006, 5, 1940–1945. [Google Scholar] [CrossRef]
- Escargueil, A.E.; Soares, D.G.; Salvador, M.; Larsen, A.K.; Henriques, J.A.P. What histone code for DNA repair? Mutat. Res. Mutat. Res. 2008, 658, 259–270. [Google Scholar] [CrossRef]
- Novik, K.L.; Spinelli, J.J.; MacArthur, A.C.; Shumansky, K.; Sipahimalani, P.; Leach, S.; Lai, A.; Connors, J.M.; Gascoyne, R.D.; Gallagher, R.P.; et al. Genetic Variation in H2AFX Contributes to Risk of Non–Hodgkin Lymphoma. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Gorgoulis, V.G.; Vassiliou, L.-V.F.; Karakaidos, P.; Zacharatos, P.; Kotsinas, A.; Liloglou, T.; Venere, M.; Ditullio, R.A., Jr.; Kastrinakis, N.G.; Levy, B.; et al. Activation of the DNA damage checkpoint and genomic instability in human precancerous lesions. Nat. Cell Biol. 2005, 434, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Orywal, K.; Jelski, W.; Zdrodowski, M.; Szmitkowski, M. The diagnostic value of alcohol dehydrogenase isoenzymes and aldehyde dehydrogenase measurement in the sera of patients with endometrial cancer. Anticancer Res. 2013, 33, 3725–3730. [Google Scholar]
- Orywal, K.; Jelski, W.; Zdrodowski, M.; Szmitkowski, M. The activity of class I, II, III and IV alcohol dehydrogenase isoenzymes and aldehyde dehydrogenase in ovarian cancer and ovarian cysts. Adv. Med. Sci. 2013, 58, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Akçay, T.; Dinçer, Y.; Alademir, Z.; Aydınlı, K.; Arvas, M.; Demirkıran, F.; Kösebay, D. Significance of the O6-methylguanine-DNA methyltransferase and glutathione S-transferase activity in the sera of patients with malignant and benign ovarian tumors. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 119, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Orywal, K.; Szmitkowski, M. Alcohol dehydrogenase and aldehyde dehydrogenase in malignant neoplasms. Clin. Exp. Med. 2017, 17, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.G.; Liu, S.; et al. ALDH1 Is a Marker of Normal and Malignant Human Mammary Stem Cells and a Predictor of Poor Clinical Outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, E.; Wei, H.; Liao, X.; Xu, Y.; Li, S.; Zeng, X. Prognostic value of alcohol dehydrogenase mRNA expression in gastric cancer. Oncol. Lett. 2018, 15, 5505–5516. [Google Scholar] [CrossRef] [Green Version]
- Bery, A.; Leung, F.; Smith, C.R.; Diamandis, E.; Kulasingam, V. Deciphering the ovarian cancer ascites fluid peptidome. Clin. Proteom. 2014, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Ricciardelli, C.; Lokman, N.; Cheruvu, S.; Tan, I.A.; Ween, M.P.; Pyragius, C.E.; Ruszkiewicz, A.; Hoffmann, P.; Oehler, M.K. Transketolase is upregulated in metastatic peritoneal implants and promotes ovarian cancer cell proliferation. Clin. Exp. Metastasis 2015, 32, 441–455. [Google Scholar] [CrossRef]
- Zhao, M.; Ye, M.; Zhou, J.; Zhu, X. Prognostic values of transketolase family genes in ovarian cancer. Oncol. Lett. 2019, 18, 4845–4857. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Histopathology Type | n Patient | Age Mean (Years) (SD) | p |
|---|---|---|---|
| Papillary serous carcinoma | 38 | 63.8 (1.6–2.9) | NS |
| Benign changes | 41 | 50.3 (1.1–1.9) | |
| Simple cysts | 21 | 44.9 (0.7–1.8) | NS |
| Normal ovary | 20 | 58.2 (1.4–3.1) |
| n Patient | Age Mean (Years) (SD) | |
|---|---|---|
| Papillary serous adenocarcinoma | 38 | 63.8 (2.1–3.0) |
| G1 | 6 | 66.6 (1.7–3.2) |
| G2 | 14 | 61.5 (2.8–4.1) |
| G3 | 18 | 62.3 (2.0–2.9) |
| FIGO I–II | 2 | 60.2 (1.6–3.2) |
| FIGO III–IV | 36 | 67.1 (2.4–3.4) |
| Variable | Statistics Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| n Patient | Mean | −95% CI | +95% CI | Median | Minimum | Maximum | Standard Deviation | p | |
| YAP1 group A | 41 | 0.164 | 0.054 | 0.272 | 0.048 | 0.001 | 1.561 | 0.345 | 0.003 |
| YAP1 group B | 40 | 0.011 | 0.005 | 0.016 | 0.005 | 0.000 | 0.091 | 0.0016 | |
| SMAD2 group A | 41 | 0.232 | 0.052 | 0.413 | 0.061 | 0.001 | 3.334 | 0.579 | 0.053 |
| SMAD2 group B | 40 | 0.059 | 0.001 | 0.116 | 0.019 | 0.000 | 1.000 | 0.171 | |
| SMAD3 group A | 41 | 0.018 | 0.011 | 0.026 | 0.010 | 0.001 | 0.351 | 0.092 | NS |
| SMAD3 group B | 40 | 0.081 | –0.064 | 0.225 | 0.005 | 0.000 | 0.106 | 0.002 | |
| CD71 group A | 41 | 1.339 | 0.752 | 1.925 | 0.378 | 0.017 | 7.501 | 1.880 | 0.048 |
| CD71 group B | 40 | 0.887 | 0.403 | 1.371 | 0.167 | 0.006 | 7.412 | 1.533 | |
| TEAD4 group A | 41 | 0.371 | 0.200 | 0.542 | 0.088 | 0.011 | 2.351 | 0.548 | 0.006 |
| TEAD4 group B | 40 | 0.061 | 0.042 | 0.171 | 0.016 | 0.000 | 0.999 | 0.202 | |
| H2AX group A | 41 | 0.826 | 0.424 | 1.217 | 0.222 | 0.013 | 5.864 | 1.258 | 0.023 |
| H2AXgroup B | 40 | 0.511 | 0.195 | 0.829 | 0.110 | 0.017 | 5.671 | 1.002 | |
| ADH1A group A | 41 | 0.184 | 0.083 | 0.284 | 0.086 | 0.001 | 1.700 | 0.313 | 0.003 |
| ADH1A group B | 40 | 0.039 | 0.017 | 0.061 | 0.006 | 0.000 | 0.305 | 0.066 | |
| TKT group A | 41 | 14.861 | 9.241 | 20.497 | 7.564 | 1.108 | 162.113 | 18.066 | 0.003 |
| TKT group B | 40 | 7.933 | –0.241 | 16.092 | 1.842 | 0.008 | 6.280 | 25.885 | |
| TKTL1 group A | 41 | 0.508 | –0.053 | 1.071 | 0.001 | 0.000 | 6.681 | 1.411 | 0.016 |
| TKTL1group B | 40 | 0.002 | –0.001 | 0.006 | 0.000 | 0.000 | 0.051 | 0.018 | |
| Variable | Statistics Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| n Patient | Mean | −95% CI | +95% CI | Median | Minimum | Maximum | Standard Deviation | p | |
| YAP1 group B1 | 21 | 0.012 | 0.021 | 0.040 | 0.009 | 0.001 | 1.002 | 0.021 | NS |
| YAP1 group B2 | 20 | 0.008 | 0.024 | 0.031 | 0.006 | 0.000 | 0.893 | 0.009 | |
| SMAD2 group B1 | 21 | 0.111 | 0.001 | 0.116 | 0.019 | 0.000 | 1.000 | 0.171 | 0.04 |
| SMAD2 group B2 | 20 | 0.056 | 0.001 | 0.078 | 0.009 | 0.000 | 1.002 | 0.087 | |
| SMAD3 group B1 | 21 | 0.016 | 0.012 | 0.033 | 0.010 | 0.000 | 0.311 | 0.078 | NS |
| SMAD3 group B2 | 20 | 0.020 | 0.014 | 0.061 | 0.014 | 0.001 | 0.408 | 0.068 | |
| CD71 group B1 | 21 | 0.780 | 0.356 | 1.206 | 0.423 | 0.006 | 7.112 | 1.432 | NS |
| CD71 group B2 | 20 | 0.911 | 0.423 | 1.523 | 0.622 | 0.008 | 7.004 | 1.321 | |
| TEAD4 group B1 | 21 | 0.067 | 0.042 | 0.171 | 0.016 | 0.000 | 0.999 | 0.202 | NS |
| TEAD4 group B2 | 20 | 0.045 | 0.039 | 0.121 | 0.023 | 0.002 | 1.003 | 0.187 | |
| H2AX group B1 | 21 | 0.387 | 0.109 | 0.912 | 0.110 | 0.012 | 5.076 | 0.976 | NS |
| H2AX group B2 | 20 | 0.299 | 0.123 | 0.867 | 0.132 | 0.001 | 5.231 | 0.954 | |
| ADH1A group B1 | 21 | 0.027 | 0.020 | 0.049 | 0.016 | 0.000 | 0.298 | 0.066 | NS |
| ADH1A group B2 | 20 | 0.034 | 0.022 | 0.061 | 0.023 | 0.001 | 0.311 | 0.052 | |
| TKT group B1 | 21 | 8.213 | 0.876 | 17.344 | 1.997 | 0.009 | 7.243 | 23.435 | NS |
| TKT group B2 | 20 | 7.878 | 0.767 | 15.231 | 1,878 | 0.013 | 7.006 | 20.878 | |
| TKTL1 group B1 | 21 | 0.002 | 0.001 | 0.005 | 0.000 | 0.000 | 0.051 | 0.018 | NS |
| TKTL1 group B2 | 20 | 0.001 | 0.001 | 0.006 | 0.000 | 0.000 | 0.043 | 0.020 | |
| Ovarian Cancer | Non Ovarian Cancer | p | |
|---|---|---|---|
| YAP | POS + + + | POS - | <0.05 |
| NEG − | NEG + | ||
| TEAD4 | POS +++ | POS − | <0.05 |
| NEG −x | NEG + | ||
| ADH1A1 | POS +++ | POS + | <0.5 |
| NEG − | NEG + | ||
| H2AX | POS ++ | POS ++ | NS |
| NEG + | NEG + | ||
| CD71 | POS ++ | POS + | NS |
| NEG + | NEG + | ||
| TKL1 | POS + | POS + | NS |
| NEG + | NEG − |
| Proteins | Age | p | Grade | p | FIGO | p | Metastasized | p |
|---|---|---|---|---|---|---|---|---|
| YAP | <55 | NS | G1 | NS | I,II | 0.01 | Yes | 0.04 |
| >55 | G3 | III,IV | No | |||||
| TEAD4 | <55 | NS | G1 | NS | I,II | 0.01 | Yes | 0.02 |
| >55 | G3 | III,IV | No | |||||
| ADH1A1 | <55 | NS | G1 | NS | I,II | NS | Yes | NS |
| >55 | G3 | III,IV | No | |||||
| H2AX | <55 | NS | G1 | NS | I,II | NS | Yes | NS |
| >55 | G3 | III,IV | No | |||||
| CD71 | <55 | NS | G1 | NS | I,II | NS | Yes | NS |
| >55 | G3 | III,IV | No | |||||
| TKL1 | <55 | NS | G1 | NS | I,II | NS | Yes | NS |
| >55 | G3 | III,IV | No |
| Univariate Analysis (Cox Regression Model) | ||||||
|---|---|---|---|---|---|---|
| PFS | OS | |||||
| HR | 95% CI | p-Value | HR | 95% CI | p-Value | |
| Age | 1.019 | 0.78–1.22 | 0.061 | 1.21 | 0.99–1.26 | 0.037 |
| Grade 1 vs 3 | 2.04 | 1.68–2.43 | 0.049 | 2.18 | 1.74–2.25 | 0.008 |
| YAP median | 1.43 | 1.21–1.62 | 0.031 | 1.40 | 1.33–1.48 | 0.02 |
| TEAD4 median | 1.28 | 1.11–1.47 | 0.026 | 1.09 | 0.97–1.30 | 0.017 |
| SMAD2 median | 1.06 | 1.02–1.21 | 0.048 | 1.16 | 1.04–1.27 | NS |
| SMAD3 median | 0.98 | 0.90–1.13 | NS | 0.89 | 0.72–0.99 | NS |
| H2A.X | 0.94 | 0.85–1.17 | NS | 0.97 | 0.76–1.08 | NS |
| ADH1.A | 1.23 | 1.07–1.32 | NS | 1.24 | 1.12–1.31 | 0.043 |
| CD71 | 1.11 | 0.82–1.30 | 0.053 | 1.37 | 1.18–145 | 0.024 |
| TKT | 1.08 | 0.88–1.21 | 0.015 | 1.15 | 0.99–1.17 | 0.028 |
| TKTL1 | 1.37 | 1.14–1.44 | NS | 1.26 | 1.18–1.38 | NS |
| Multivariate Analysis (Cox Regression Model) | ||||||
| PFS | OS | |||||
| HR | 95% CI | p-Value | HR | 95% CI | p-Value | |
| YAP median | 1.22 | 1.06–1.31 | 0.027 | 1.19 | 0.98–1.24 | 0.041 |
| YAP 95 percentile | 1.49 | 1.36–1.59 | 0.046 | 1.31 | 1.26–1.39 | 0.039 |
| TEAD4 median | 1.10 | 0.95–1.27 | 0.007 | 1.17 | 1.08–1.32 | NS |
| TEAD4 95 percentile | 1.34 | 1.26–1.43 | NS | 1.02 | 0.94–1.22 | NS |
| CD71 median | 1.21 | 1.11–1.36 | 0.033 | 1.37 | 1.24–1.43 | NS |
| CD71 95 percentile | 1.08 | 0.96–1.21 | NS | 1.19 | 1.12–1.34 | 0.018 |
| TKT median | 0.99 | 0.90–1.17 | NS | 1.07 | 0.97–1.22 | NS |
| TKT 95 percentile | 1.12 | 1.01–1.28 | 0.004 | 1.28 | 1.13–1.32 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cymbaluk-Płoska, A.; Chudecka, K.; Chudecka-Głaz, A.; Piotrowska, K.; Kwiatkowski, S.; Tarnowski, M. Importance of Potential New Biomarkers in Patient with Serouse Ovarian Cancer. Diagnostics 2021, 11, 1026. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11061026
Cymbaluk-Płoska A, Chudecka K, Chudecka-Głaz A, Piotrowska K, Kwiatkowski S, Tarnowski M. Importance of Potential New Biomarkers in Patient with Serouse Ovarian Cancer. Diagnostics. 2021; 11(6):1026. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11061026
Chicago/Turabian StyleCymbaluk-Płoska, Aneta, Karolina Chudecka, Anita Chudecka-Głaz, Katarzyna Piotrowska, Sebastian Kwiatkowski, and Maciej Tarnowski. 2021. "Importance of Potential New Biomarkers in Patient with Serouse Ovarian Cancer" Diagnostics 11, no. 6: 1026. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11061026