
Caspase 9 and Caspase 3 Immunohistochemical Pattern in Skeletal and Cardiac Muscles at Different Times after Death: An Experimental Study on PMI Estimation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Immunohistochemistry
2.2. Image Analysis
2.3. Statistical Analysis
3. Results
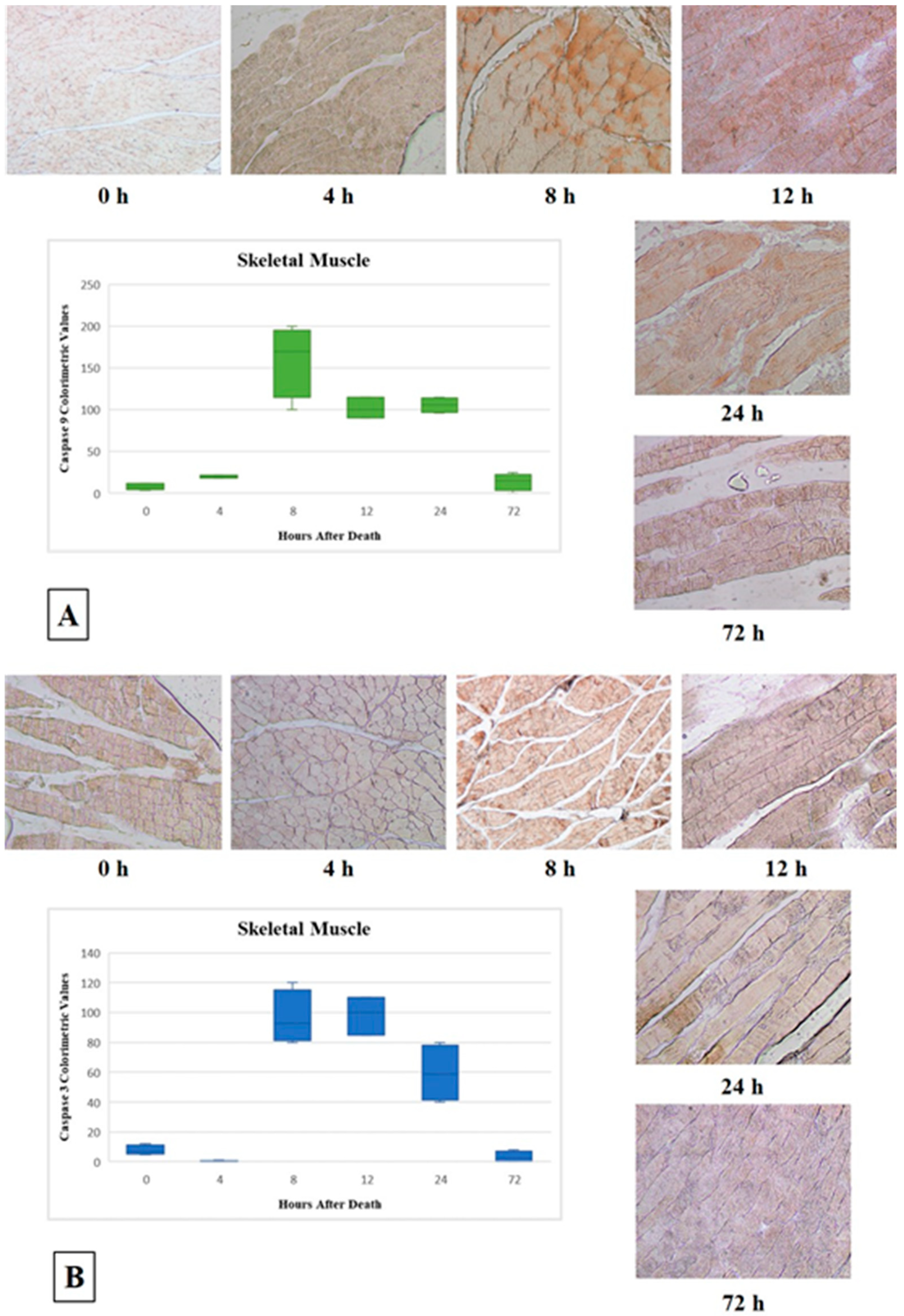
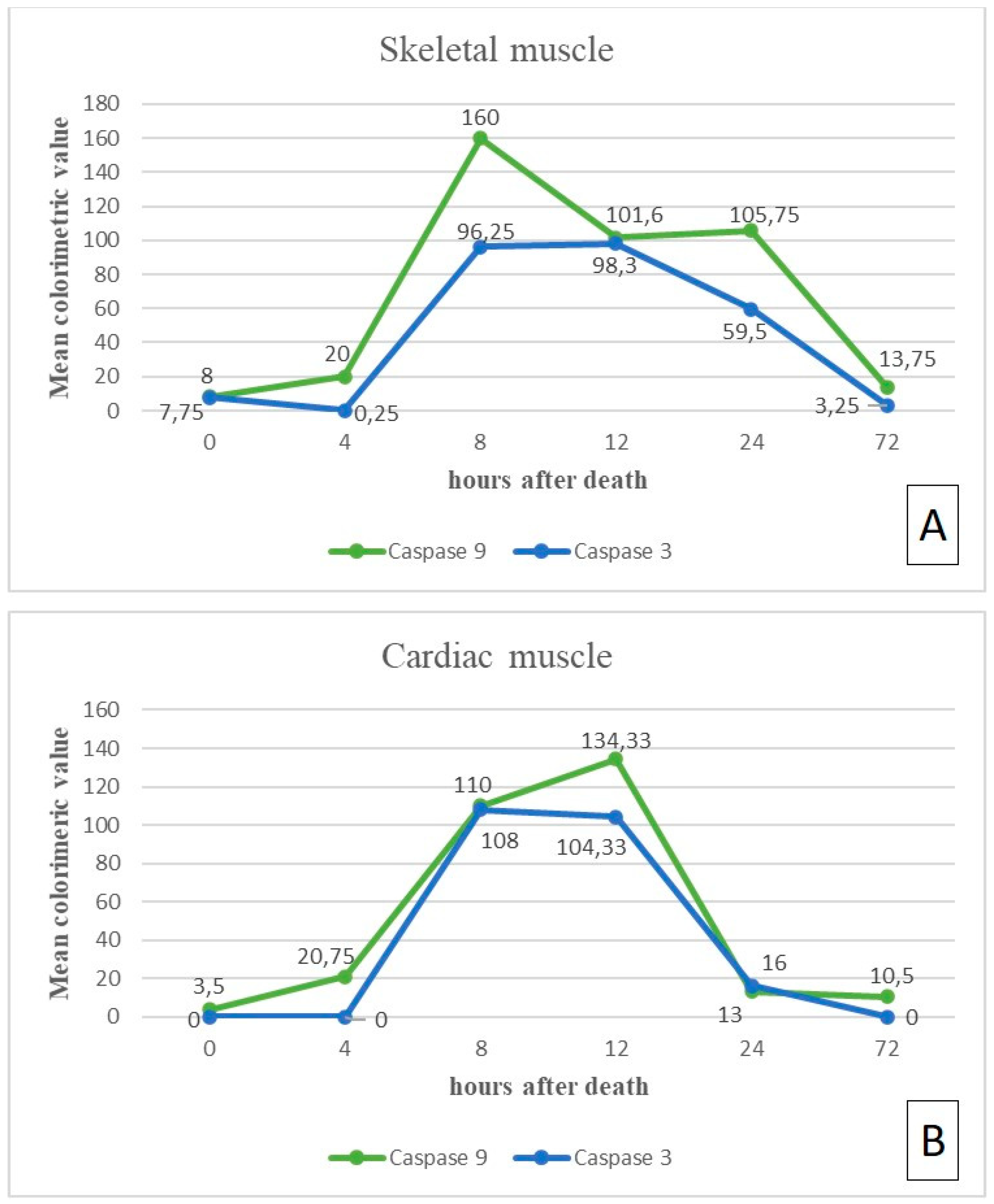
3.1. Skeletal Muscle
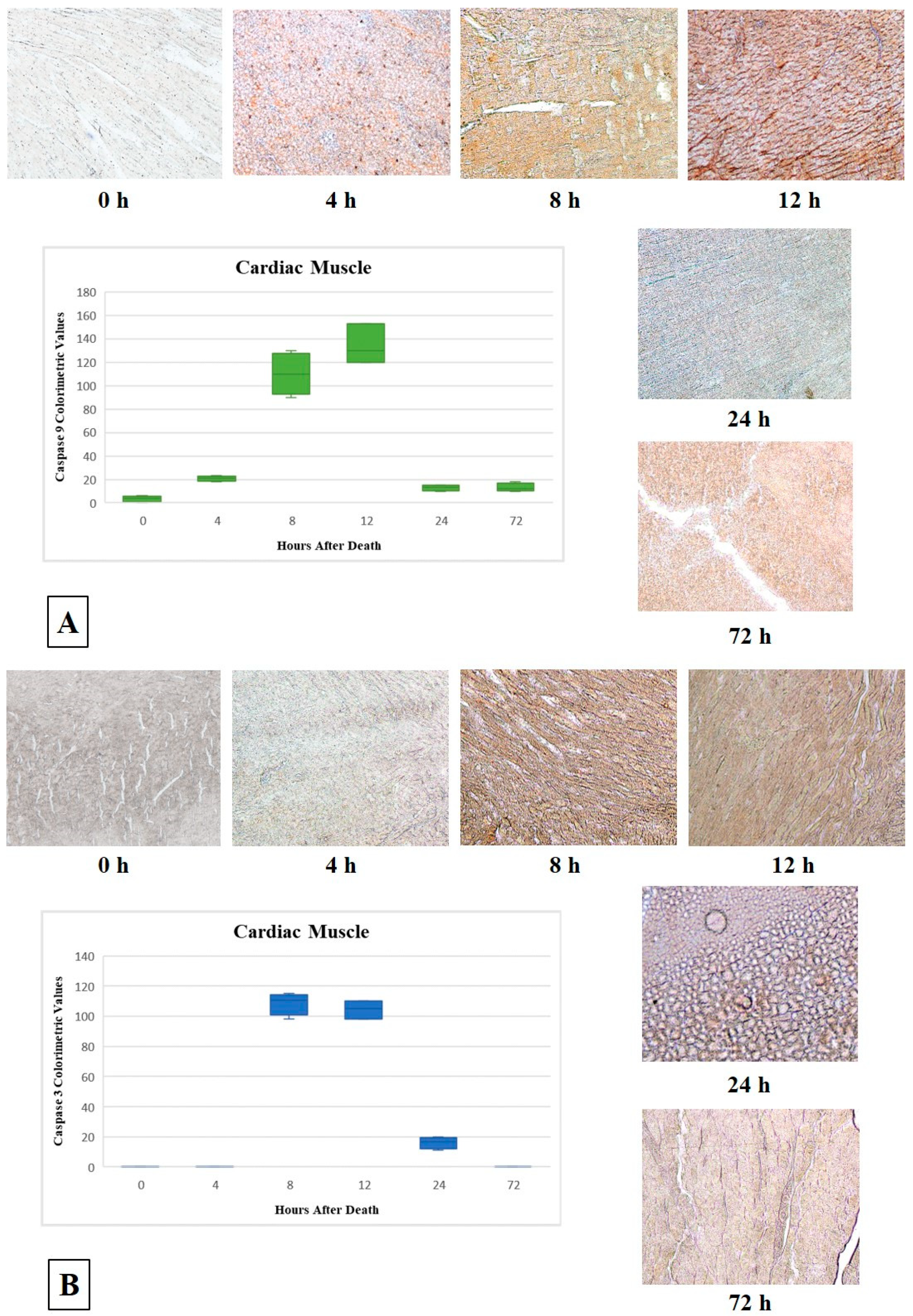
3.2. Cardiac Muscle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Henssge, C.; Althaus, L.; Bolt, J.; Freislederer, A.; Haffner, H.-T.; Hoppe, B.; Schneider, V. Experiences with a compound method for estimating the time since death. Int. J. Leg. Med. 2000, 113, 303–319. [Google Scholar] [CrossRef]
- Madea, B. Methods for determining time of death. Forensic Sci. Med. Pathol. 2016, 12, 451–485. [Google Scholar] [CrossRef]
- Mathur, A.; Agrawal, Y. An overview of methods used for estimation of time since death. Aust. J. Forensic Sci. 2011, 43, 275–285. [Google Scholar] [CrossRef]
- Madea, B.; Henssge, C. Electrical excitability of skeletal muscle postmortem in casework. Forensic Sci. Int. 1990, 47, 207–227. [Google Scholar] [CrossRef]
- Donaldson, A.E.; Lamont, I.L. Biochemistry Changes That Occur after Death: Potential Markers for Determining Post-Mortem Interval. PLoS ONE 2013, 8, 82011. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Lv, M.; Li, Z.; Tian, H.; Qu, S.; Jin, B.; Long, B.; Liang, W.; Zhang, L. Postmortem interval determination using mRNA markers and DNA normalization. Int. J. Leg. Med. 2019, 134, 149–157. [Google Scholar] [CrossRef]
- Noshy, P.A. Postmortem expression of apoptosis-related genes in the liver of mice and their use for estimation of the time of death. Int. J. Leg. Med. 2021, 135, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Madea, B. Is there recent progress in the estimation of the postmortem interval by means of thanatochemistry? Forensic Sci. Int. 2005, 151, 139–149. [Google Scholar] [CrossRef]
- Tattoli, L.; Tsokos, M.; Sautter, J.; Anagnostopoulos, J.; Maselli, E.; Ingravallo, G.; Delia, M.; Solarino, B. Postmortem bone marrow analysis in forensic science: Study of 73 cases and review of the literature. Forensic Sci. Int. 2014, 234, 72–78. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, L.; Yin, Y.; Yang, E. Systematic Analysis of Gene Expression Patterns Associated with Postmortem Inter-val in Human Tissues. Sci. Rep. 2017, 7, 5435. [Google Scholar] [CrossRef]
- Scrivano, S.; Sanavio, M.; Tozzo, P.; Caenazzo, L. Analysis of RNA in the Estimation of Post-Mortem Interval: A Review of Cur-rent Evidence. Int. J. Legal Med. 2019, 133, 1629–1640. [Google Scholar] [CrossRef] [PubMed]
- Pigaiani, N.; Bertaso, A.; De Palo, E.F.; Bortolotti, F.; Tagliaro, F. Vitreous humor endogenous compounds analysis for post-mortem forensic investigation. Forensic Sci. Int. 2020, 310, 110235. [Google Scholar] [CrossRef]
- Locci, E.; Bazzano, G.; Chighine, A.; Locco, F.; Ferraro, E.; Demontis, R.; D’Aloja, E. Forensic NMR metabolomics: One more arrow in the quiver. Metabolomics 2020, 16, 1–16. [Google Scholar] [CrossRef]
- Maiese, A.; Scatena, A.; Costantino, A.; Di Paolo, M.; La Russa, R.; Turillazzi, E.; Frati, P.; Fineschi, V. MicroRNAs as Useful Tools to Estimate Time Since Death. A Systematic Review of Current Literature. Diagnostics 2021, 11, 64. [Google Scholar] [CrossRef]
- Zissler, A.; Stoiber, W.; Steinbacher, P.; Geissenberger, J.; Monticelli, F.C.; Pittner, S. Postmortem Protein Degradation as a Tool to Estimate the PMI: A Systematic Review. Diagnostics 2020, 10, 1014. [Google Scholar] [CrossRef]
- Riedl, S.J.; Shi, Y. Molecular mechanisms of caspase regulation during apoptosis. Nat. Rev. Mol. Cell Biol. 2004, 5, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.M. Caspases: The executioners of apoptosis. Biochem. J. 1997, 326, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabucedo, A.J.; Furton, K.G. Estimation of postmortem interval using the protein marker cardiac Troponin I. Forensic Sci. Int. 2003, 134, 11–16. [Google Scholar] [CrossRef]
- Jellinghaus, K.; Urban, P.K.; Hachmann, C.; Bohnert, M.; Hotz, G.; Rosendahl, W.; Wittwer-Backofen, U. Collagen deg-radation as a possibility to determine the post-mortem interval (PMI) of human bones in a forensic context—A survey. Leg. Med. 2019, 36, 96–102. [Google Scholar] [CrossRef]
- Pittner, S.; Monticelli, F.C.; Pfisterer, A.; Zissler, A.; Sänger, A.M.; Stoiber, W.; Steinbacher, P. Postmortem degradation of skeletal muscle proteins: A novel approach to determine the time since death. Int. J. Leg. Med. 2015, 130, 421–431. [Google Scholar] [CrossRef]
- Prieto-Bonete, G.; Pérez-Cárceles, M.D.; López, A.M.; Pérez-Martínez, C.; Luna, A. Association between protein profile and postmortem interval in human bone remains. J. Proteom. 2019, 192, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.-J.; Han, L.-H.; Cong, R.-S.; Liang, J. Caspase Family Proteases and Apoptosis. Acta Biochim. Biophys. Sin. 2005, 37, 719–727. [Google Scholar] [CrossRef] [Green Version]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase Functions in Cell Death and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, 008656. [Google Scholar] [CrossRef]
- Pittner, S.; Ehrenfellner, B.; Zissler, A.; Racher, V.; Trutschnig, W.; Bathke, A.C.; Sänger, A.M.; Stoiber, W.; Steinbacher, P.; Monticelli, F.C. First application of a protein-based approach for time since death estimation. Int. J. Leg. Med. 2016, 131, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.-M.; Zissler, A.; Kim, E.; Ehrenfellner, B.; Cho, E.; Lee, S.-I.; Steinbacher, P.; Na Yun, K.; Shin, J.H.; Kim, J.Y.; et al. Postmortem proteomics to discover biomarkers for forensic PMI estimation. Int. J. Leg. Med. 2019, 133, 899–908. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-G.; Yang, K.E.; Hwang, J.W.; Kang, H.-S.; Lee, S.-Y.; Choi, S.; Shin, J.; Jang, I.-S.; An, H.J.; Chung, H.; et al. Degradation of Kidney and Psoas Muscle Proteins as Indicators of Post-Mortem Interval in a Rat Model, with Use of Lateral Flow Technology. PLoS ONE 2016, 11, 0160557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zissler, A.; Ehrenfellner, B.; Foditsch, E.E.; Monticelli, F.C.; Pittner, S. Does altered protein metabolism interfere with postmortem degradation analysis for PMI estimation? Int. J. Leg. Med. 2018, 132, 1349–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foditsch, E.E.; Saenger, A.M.; Monticelli, F.C. Skeletal muscle proteins: A new approach to delimitate the time since death. Int. J. Leg. Med. 2015, 130, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Mondello, C.; Cardia, L.; Bartoloni, G.; Asmundo, A.; Spagnolo, E.V. Immunohistochemical study on dystrophin expression in CAD-related sudden cardiac death: A marker of early myocardial ischaemia. Int. J. Leg. Med. 2018, 132, 1333–1339. [Google Scholar] [CrossRef]
- Spagnolo, E.V.; Mondello, C.; Cardia, L.; Minutoli, L.; Puzzolo, D.; Asmundo, A.; Macaione, V.; Alibrandi, A.; Malta, C.; Baldino, G.; et al. Post-Mortem Immunohistochemical Evidence of β2-Adrenergic Receptor Expression in the Adrenal Gland. Int. J. Mol. Sci. 2019, 20, 3065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stassi, C.; Mondello, C.; Baldino, G.; Spagnolo, E.V. Post-Mortem Investigations for the Diagnosis of Sepsis: A Review of Literature. Diagnostics 2020, 10, 849. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | PMI | N | Skeletal Muscle | Cardiac Muscle | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Caspase 9 | Mean CV * | Caspase 3 | Mean CV * | Caspase 9 | Mean CV * | Caspase 3 | Mean CV * | |||
| G1 | T0 = 0 h | 1 | CV * 4 (+) | 8 (+) | CV * 5 (+) | 7.75 (+) | CV * 6 (+) | 3.5 (+) | 0 | 0 |
| 2 | CV * 6 (+) | CV * 8 (+) | CV * 4 (+) | 0 | ||||||
| 3 | CV * 12 (+) | CV * 6 (+) | CV * 4 (+) | 0 | ||||||
| 4 | CV * 10 (+) | CV * 12 (+) | 0 | 0 | ||||||
| G2 | T1 = 4 h | 5 | CV * 20 (+) | 20 (+) | 0 | 0.25 (+) | CV * 22 (+) | 20.75 (+) | 0 | 0 |
| 6 | CV * 22 (+) | CV * 1 (+) | CV * 18 (+) | 0 | ||||||
| 7 | CV * 18 (+) | 0 | CV * 23 (+) | 0 | ||||||
| 8 | CV * 20 (+) | 0 | CV * 20 (+) | 0 | ||||||
| G3 | T2 = 8 h | 9 | CV * 200 (+ + + +) | 160 (+ + + +) | CV * 120 (+ + +) | 96.25 (+ +) | CV * 100 (+ +) | 110 (+ + +) | CV * 110 (+ + +) | 108.5 (+ + +) |
| 10 | CV * 160 (+ + + +) | CV * 80 (+ +) | CV * 130 (+ + +) | CV * 115 (+ + +) | ||||||
| 11 | CV * 180 (+ + + +) | CV * 85 (+ +) | CV * 90 (+ +) | CV * 98 (+ +) | ||||||
| 12 | CV * 100 (+ +) | CV * 100 (+ +) | CV * 120 (+ + +) | CV * 111 (+ + +) | ||||||
| G4 | T3 = 12 h | 13 | CV * 115 (+ + +) | 101.66 (+ + +) | CV * 100 (+ +) | 98.33 (+ +) | CV * 120 (+ + +) | 134.33 (+ + +) | CV * 110 (+ + +) | 104.33 (+ + +) |
| 14 | CV * 100 (+ +) | CV * 110 (+ + +) | CV * 153 (+ + + +) | CV * 98 (+ +) | ||||||
| 15 | CV * 90 (+ +) | CV * 85 (+ +) | CV * 130 (+ + +) | CV * 105 (+ + +) | ||||||
| G5 | T4 = 24 h | 16 | CV * 100 (+ +) | 105.75 (+ + +) | CV * 80 (+ +) | 59.5 (+ +) | CV * 10 (+) | 13 (+) | CV * 15 (+) | 16 (+) |
| 17 | CV * 112 (+ + +) | CV * 73 (+ +) | CV * 12 (+) | CV * 18 (+) | ||||||
| 18 | CV * 96 (+ +) | CV * 45 (+) | CV * 15 (+) | CV * 20 (+) | ||||||
| 19 | CV * 115 (+ + +) | CV * 40 (+) | CV * 15 (+) | CV * 11 (+) | ||||||
| G6 | T5 = 72 h | 20 | 0 | 13.75 (+) | 0 | 3.25 (+) | CV * 10 (+) | 13 (+) | 0 | 0 |
| 21 | CV * 15 (+) | 0 | CV * 18 (+) | 0 | ||||||
| 22 | CV * 15 (+) | CV * 8 (+) | CV * 12 (+) | 0 | ||||||
| 23 | CV * 25 (+) | CV * 5 (+) | CV * 12 (+) | 0 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mondello, C.; Stassi, C.; Minutoli, L.; Baldino, G.; Alibrandi, A.; Spatola, G.F.; Uzzo, M.L.; Micali, A.; Puzzolo, D.; Asmundo, A.; et al. Caspase 9 and Caspase 3 Immunohistochemical Pattern in Skeletal and Cardiac Muscles at Different Times after Death: An Experimental Study on PMI Estimation. Diagnostics 2021, 11, 1062. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11061062
Mondello C, Stassi C, Minutoli L, Baldino G, Alibrandi A, Spatola GF, Uzzo ML, Micali A, Puzzolo D, Asmundo A, et al. Caspase 9 and Caspase 3 Immunohistochemical Pattern in Skeletal and Cardiac Muscles at Different Times after Death: An Experimental Study on PMI Estimation. Diagnostics. 2021; 11(6):1062. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11061062
Chicago/Turabian StyleMondello, Cristina, Chiara Stassi, Letteria Minutoli, Gennaro Baldino, Angela Alibrandi, Giovanni Francesco Spatola, Maria Laura Uzzo, Antonio Micali, Domenico Puzzolo, Alessio Asmundo, and et al. 2021. "Caspase 9 and Caspase 3 Immunohistochemical Pattern in Skeletal and Cardiac Muscles at Different Times after Death: An Experimental Study on PMI Estimation" Diagnostics 11, no. 6: 1062. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11061062