
Differential Expression of LncRNA in Bladder Cancer Development
 ,
,  , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Enrollment, Sample Collection and Patient Evaluation
2.2. In Silico LncRNA Gene Expression Analysis
2.3. LncRNA Gene Expression Analysis in Tissue Biopsies
2.4. Statistical Analysis
3. Results
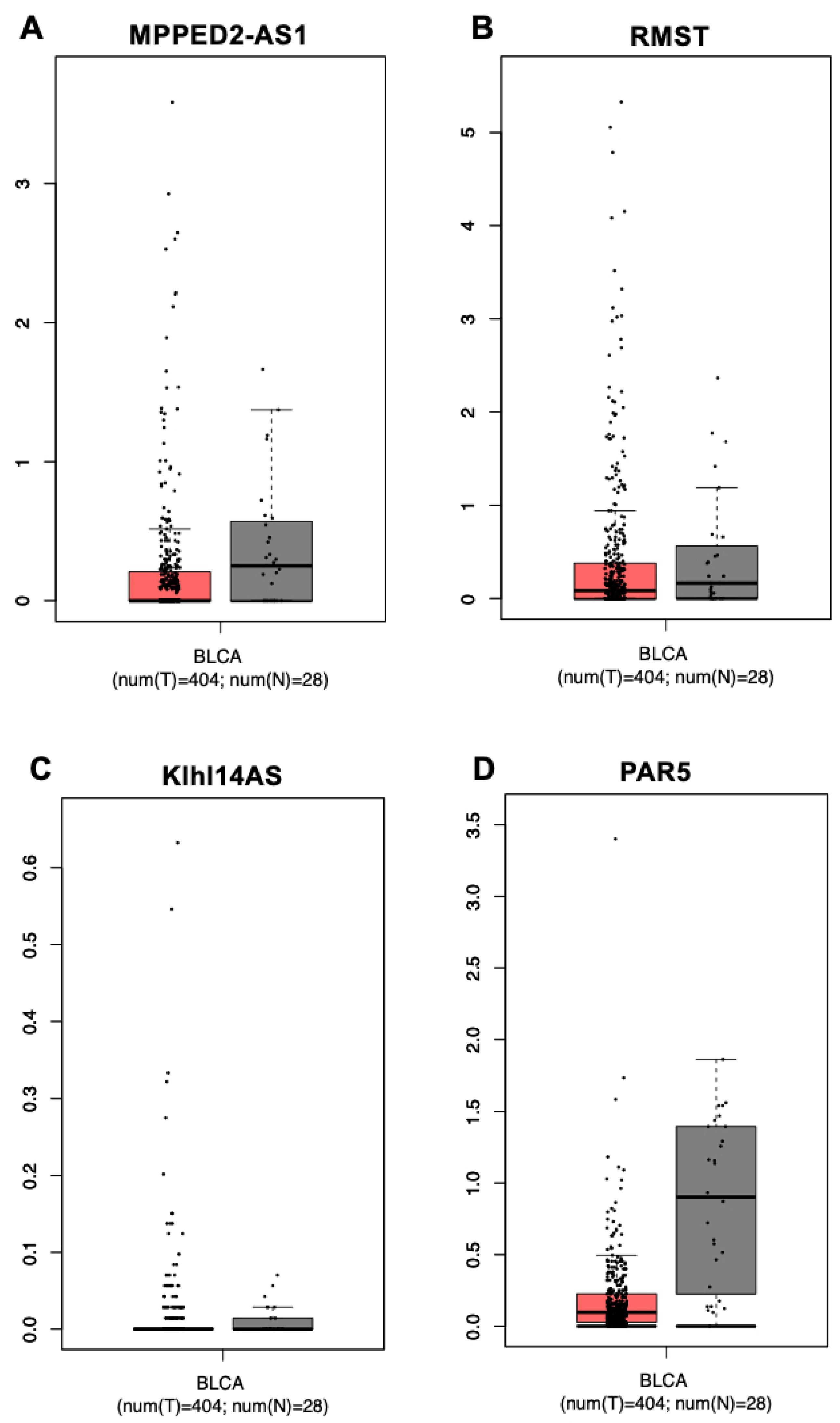
3.1. In Silico Analysis of Gene Expression of the lncRNAs MPPED2-AS1, RMST, KLHL14-AS and PAR5 in Bladder Urothelial Cancer
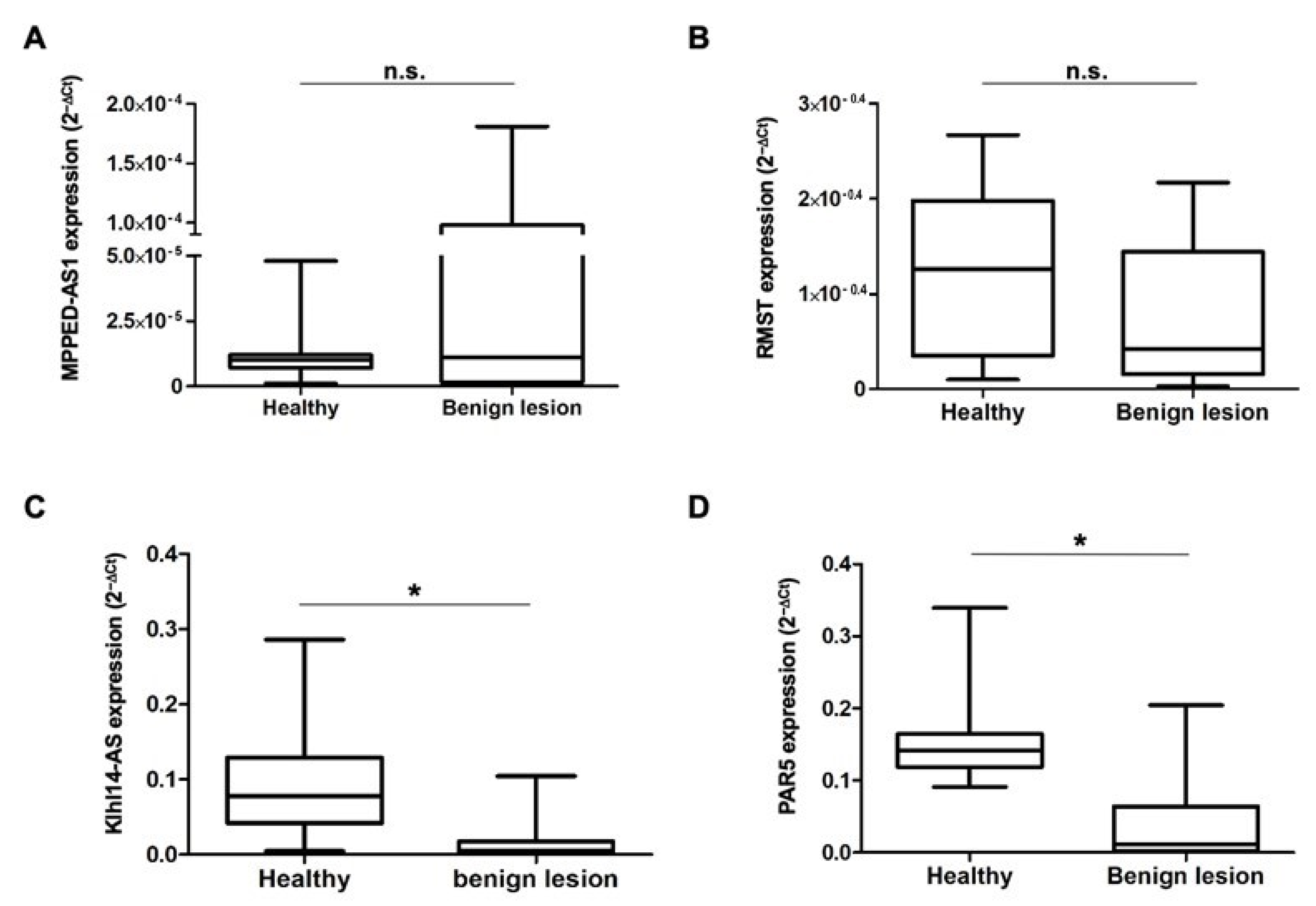
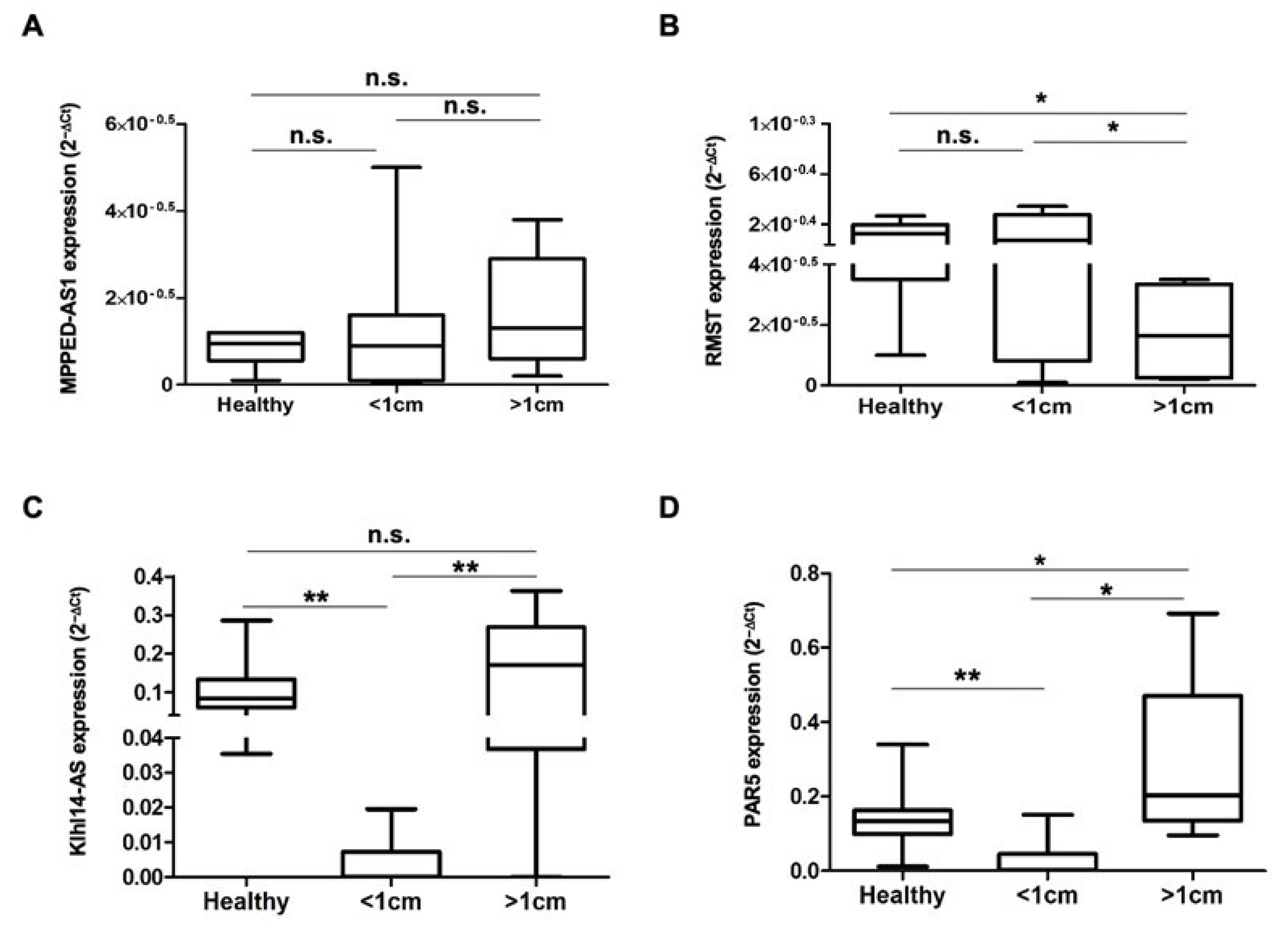
3.2. The lncRNAs MPPED2-AS1, RMST, Klhl14AS and PAR5 Show Differential Expression in Bladder Lesions of Different Degree of Malignancy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antoni, S.; Ferlay, J.; Soerjomataram, I.; Znaor, A.; Jemal, A.; Bray, F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur. Urol. 2017, 71, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Compérat, E.; Larré, S.; Rouprêt, M.; Neuzillet, Y.; Pignot, G.; Quintens, H.; Houede, N.; Roy, C.; Durand, X.; Varinot, J.; et al. Clinicopathological characteristics of urothelial bladder cancer in patients less than 40 years old. Virchows Arch. 2015, 466, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Burger, M.; Catto, J.W.F.; Dalbagni, G.; Grossman, H.B.; Herr, H.; Karakiewicz, P.; Kassouf, W.; Kiemeney, L.A.; La Vecchia, C.; Shariat, S.; et al. Epidemiology and Risk Factors of Urothelial Bladder Cancer. Eur. Urol. 2013, 63, 234–241. [Google Scholar] [CrossRef]
- Teoh, J.Y.-C.; Huang, J.; Ko, W.Y.-K.; Lok, V.; Choi, P.; Ng, C.-F.; Sengupta, S.; Mostafid, H.; Kamat, A.M.; Black, P.C.; et al. Global Trends of Bladder Cancer Incidence and Mortality, and Their Associations with Tobacco Use and Gross Domestic Product Per Capita. Eur. Urol. 2020, 78, 893–906. [Google Scholar] [CrossRef] [PubMed]
- Colt, J.S.; Friesen, M.C.; Stewart, P.A.; Donguk, P.; Johnson, A.; Schwenn, M.; Karagas, M.R.; Armenti, K.; Waddell, R.; Verrill, C.; et al. A case-control study of occupational exposure to metalworking fluids and bladder cancer risk among men. Occup. Environ. Med. 2014, 71, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Pesch, B.; the UroScreen Study Group; Taeger, D.; Johnen, G.; Gawrych, K.; Bonberg, N.; Schwentner, C.; Wellhäußer, H.; Kluckert, M.; Leng, G.; et al. Screening for bladder cancer with urinary tumor markers in chemical workers with exposure to aromatic amines. Int. Arch. Occup. Environ. Health 2014, 87, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Yafi, F.A.; Brimo, F.; Steinberg, J.; Aprikian, A.G.; Tanguay, S.; Kassouf, W. Prospective analysis of sensitivity and specificity of urinary cytology and other urinary biomarkers for bladder cancer. Urol. Oncol. Semin. Orig. Investig. 2015, 33, 66.e25–66.e31. [Google Scholar] [CrossRef]
- Têtu, B. Diagnosis of urothelial carcinoma from urine. Mod. Pathol. 2009, 22 (Suppl. S2), S53–S59. [Google Scholar] [CrossRef]
- Marchese, F.P.; Huarte, M. Long non-coding RNAs and chromatin modifiers: Their place in the epigenetic code. Epigenetics 2014, 9, 21–26. [Google Scholar] [CrossRef]
- Balas, M.M.; Johnson, A.M. Exploring the mechanisms behind long noncoding RNAs and cancer. Non-Coding RNA Res. 2018, 3, 108–117. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ye, C.; Xiong, H.; Shen, Y.; Lu, Y.; Zhou, J.; Wang, L. Dysregulation of long non-coding RNA in breast cancer: An overview of mechanism and clinical implication. Oncotarget 2017, 8, 5508–5522. [Google Scholar] [CrossRef]
- Liu, Q. The emerging roles of exosomal long non-coding RNAs in bladder cancer. J. Cell. Mol. Med. 2022, 26, 966–976. [Google Scholar] [CrossRef] [PubMed]
- Sepe, R.; Pellecchia, S.; Serra, P.; D’Angelo, D.; Federico, A.; Raia, M.; Penha, R.C.C.; Decaussin-Petrucci, M.; Del Vecchio, L.; Fusco, A.; et al. The Long Non-Coding RNA RP5-1024C24.1 and Its Associated-Gene MPPED2 Are Down-Regulated in Human Thyroid Neoplasias and Act as Tumour Suppressors. Cancers 2018, 10, 146. [Google Scholar] [CrossRef] [PubMed]
- De Martino, M.; Esposito, F.; Capone, M.; Pallante, P.; Fusco, A. Noncoding RNAs in Thyroid-Follicular-Cell-Derived Carcinomas. Cancers 2022, 14, 3079. [Google Scholar] [CrossRef]
- Credendino, S.C.; Bellone, M.L.; Lewin, N.; Amendola, E.; Sanges, R.; Basu, S.; Sepe, R.; Decaussin-Petrucci, M.; Tinto, N.; Fusco, A.; et al. A ceRNA Circuitry Involving the Long Noncoding RNA Klhl14-AS, Pax8, and Bcl2 Drives Thyroid Carcinogenesis. Cancer Res. 2019, 79, 5746–5757. [Google Scholar] [CrossRef]
- Pellecchia, S.; Sepe, R.; Decaussin-Petrucci, M.; Ivan, C.; Shimizu, M.; Coppola, C.; Testa, D.; Calin, G.A.; Fusco, A.; Pallante, P. The Long Non-Coding RNA Prader Willi/Angelman Region RNA5 (PAR5) Is Downregulated in Anaplastic Thyroid Carcinomas Where It Acts as a Tumor Suppressor by Reducing EZH2 Activity. Cancers 2020, 12, 235. [Google Scholar] [CrossRef]
- Pellecchia, S.; Sepe, R.; Federico, A.; Cuomo, M.; Credendino, S.C.; Pisapia, P.; Bellevicine, C.; Nicolau-Neto, P.; Ramundo, M.S.; Crescenzi, E.; et al. The Metallophosphoesterase-Domain-Containing Protein 2 (MPPED2) Gene Acts as Tumor Suppressor in Breast Cancer. Cancers 2019, 11, 797. [Google Scholar] [CrossRef]
- Ng, S.-Y.; Bogu, G.K.; Soh, B.S.; Stanton, L.W. The Long Noncoding RNA RMST Interacts with SOX2 to Regulate Neurogenesis. Mol. Cell 2013, 51, 349–359. [Google Scholar] [CrossRef]
- Wang, P.; Shang, J.; Zhao, J.; Wang, K.; Guo, L.; Gu, J.; Wang, W. SRY-related HMG box-2 role in anaplastic thyroid cancer aggressiveness is related to the fibronectin 1 and PI3K/AKT pathway. Mol. Med. Rep. 2020, 21, 1201–1207. [Google Scholar] [CrossRef]
- Credendino, S.C.; Lewin, N.; de Oliveira, M.; Basu, S.; D’andrea, B.; Amendola, E.; Di Guida, L.; Nardone, A.; Sanges, R.; De Felice, M.; et al. Tissue- and Cell Type-Specific Expression of the Long Noncoding RNA Klhl14-AS in Mouse. Int. J. Genom. 2017, 2017, 9769171. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Zhang, H.S.; Liu, Y.; Zhang, Z.G.; Du, G.Y.; Li, H.; Yu, X.-Y.; Huang, Y.-H. Loss of TET1 facilitates DLD1 colon cancer cell migration via H3K27me3-mediated down-regulation of E-cadherin. J. Cell Physiol. 2018, 233, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed]
- Seligson, D.B.; Pantuck, A.J.; Liu, X.; Huang, Y.; Horvath, S.; Bui, M.H.; Han, K.-R.; Correa, A.J.L.; Eeva, M.; Tze, S.; et al. Epithelial cell adhesion molecule (KSA) expression: Pathobiology and its role as an independent predictor of survival in renal cell carcinoma. Clin. Cancer Res. 2004, 10, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Xiang, W.; Lyu, L.; Huang, T.; Zheng, F.; Yuan, J.; Zhang, C.; Zhang, G. The long non-coding RNA SNHG1 promotes bladder cancer progression by interacting with miR-143-3p and EZH2. J. Cell Mol. Med. 2020, 24, 11858–11873. [Google Scholar] [CrossRef]
- Went, P.; Dirnhofer, S.; Salvisberg, T.; Amin, M.B.; Lim, S.D.; Diener, P.A.; Moch, H. Expression of epithelial cell adhesion molecule (EpCam) in renal epithelial tumors. Am. J. Surg. Pathol. 2005, 29, 83–88. [Google Scholar] [CrossRef]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Luo, H.; Xu, C.; Le, W.; Ge, B.; Wang, T. lncRNA CASC11 promotes cancer cell proliferation in bladder cancer through miRNA-150. J. Cell. Biochem. 2019, 120, 13487–13493. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, W.; Lin, J.; Xiao, J.; Tian, Y. lncRNA CCAT1 promotes bladder cancer cell proliferation, migration and invasion. Int. Braz. J. Urol. 2019, 45, 549–559. [Google Scholar] [CrossRef]
- Huang, Y.; Xie, B.; Cao, M.; Lu, H.; Wu, X.; Hao, Q.; Zhou, X. LncRNA RNA Component of Mitochondrial RNA-Processing Endoribonuclease Promotes AKT-Dependent Breast Cancer Growth and Migration by Trapping MicroRNA-206. Front. Cell Dev. Biol. 2021, 9, 730538. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, M.; Liu, J. Kelch-like protein 14 promotes proliferation and migration of ovarian cancer cells. Int. J. Clin. Exp. Pathol. 2020, 13, 2950–2961. [Google Scholar] [PubMed]
- Choi, J.; Phelan, J.D.; Wright, G.W.; Haupl, B.; Huang, D.W.; Shaffer, A.L., 3rd; Young, R.M.; Wang, Z.; Zhao, H.; Yu, X.; et al. Regulation of B cell receptor-dependent NF-kappaB signaling by the tumor suppressor KLHL14. Proc. Natl. Acad. Sci. USA 2020, 117, 6092–6102. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, R.; Hong, X.; Chen, C.; Ding, Y. KLHL14: A Novel Prognostic Biomarker and Therapeutic Target for Ovarian Cancer. J. Oncol. 2022, 2022, 9799346. [Google Scholar] [CrossRef] [PubMed]
- Ropri, A.S.; DeVaux, R.S.; Eng, J.; Chittur, S.V.; Herschkowitz, J.I. Cis-acting super-enhancer lncRNAs as biomarkers to early-stage breast cancer. Breast Cancer Res. 2021, 23, 101. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Zhan, Y.; Chen, Z.; He, S.; Gong, Y.; He, A.; Li, Y.; Zhang, L.; Zhang, X.; Fang, D.; Li, X.; et al. Long non-coding RNA SOX2OT promotes the stemness phenotype of bladder cancer cells by modulating SOX2. Mol. Cancer 2020, 19, 25. [Google Scholar] [CrossRef]
- Bolha, L.; Ravnik-Glavač, M.; Glavač, D. Long Noncoding RNAs as Biomarkers in Cancer. Dis. Markers 2017, 2017, 7243968. [Google Scholar] [CrossRef]
- Qian, Y.; Shi, L.; Luo, Z. Long Non-coding RNAs in Cancer: Implications for Diagnosis, Prognosis, and Therapy. Front. Med. 2020, 7, 612393. [Google Scholar] [CrossRef]
- Creta, M.; Celentano, G.; Napolitano, L.; La Rocca, R.; Capece, M.; Califano, G.; Collà Ruvolo, C.; Mangiapia, F.; Morra, S.; Turco, C.; et al. Inhibition of Androgen Signalling Improves the Outcomes of Therapies for Bladder Cancer: Results from a Systematic Review of Preclinical and Clinical Evidence and Meta-Analysis of Clinical Studies. Diagnostics 2021, 11, 351. [Google Scholar] [CrossRef]
- Manfredi, C.; Spirito, L.; Calace, F.P.; Balsamo, R.; Terribile, M.; Stizzo, M.; Romano, L.; Napolitano, L.; Califano, G.; Cirillo, L.; et al. Oral Preparation of Hyaluronic Acid, Chondroitin Sulfate, Curcumin, and Quercetin (Ialuril((R)) Soft Gels) for the Prevention of LUTS after Intravesical Chemotherapy. Pathophysiology 2022, 29, 365–373. [Google Scholar] [CrossRef]
- Capece, M.; Spirito, L.; La Rocca, R.; Napolitano, L.; Buonopane, R.; Di Meo, S.; Sodo, M.; Bracale, U.; Longo, N.; Palmieri, A.; et al. Hexaminolevulinate blue light cystoscopy (Hal) assisted transurethral resection of the bladder tumour vs white light transurethral resection of the bladder tumour in non-muscle invasive bladder cancer (NMIBC): A retrospective analysis. Arch. Ital. Urol. Androl. 2020, 92, 17–20. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′ → 3′) | Reverse Primer (5′ → 3′) |
|---|---|---|
| 18S | TGCGAGTACTCAACACCAACA | TTGGTGAGGTCAATGTCTGC |
| Klhl14-AS | CTTCGCTGCTGAGCTGAAAC | TTGTCATCTCTGTGAGACAGC |
| MPPED2-AS1 | TGGTGCAGGGATTGTTGCAT | TGAACGACTGCAACTGCTTTG |
| RMST | GCCCAGCCCATTTCATTCAC | GCTCTGTCGTTCCACCTTGA |
| PAR5 | GGTAGGAGGAGGGTTGGCTT | TTAGCAAGGCTGGACCTCAC |
| Gender | Male: 20 (83.3%) Female: 4 (16.7%) |
|---|---|
| Age | Years: 70.54 ± 7.82 |
| Size | <1 cm: 13 (54.2%) ≥1 cm: 11 (45.8%) |
| Focality pattern | Monofocal: 17 (70.8%) Multifocal: 7 (29.2%) |
| Histology | Urothelial: 23 (95.8%) Squamous: 1 (4.2%) |
| Grade | Low: 18 (75.0%) High: 6 (25.0%) |
| Growth | Papillary: 20 (83.3%) Flat: 1 (4.2%) Solid: 3 (12.5%) |
| TNM | T0: 9 (37.5%) Ta:10 (41.7%) Tis:1 (4.2%) T1:4 (16.7%) T2-4: 0 (0%) N0: 24 (100%) M0: 24 (100%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spirito, L.; Maturi, R.; Credendino, S.C.; Manfredi, C.; Arcaniolo, D.; De Martino, M.; Esposito, F.; Napolitano, L.; Di Bello, F.; Fusco, A.; et al. Differential Expression of LncRNA in Bladder Cancer Development. Diagnostics 2023, 13, 1745. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics13101745
Spirito L, Maturi R, Credendino SC, Manfredi C, Arcaniolo D, De Martino M, Esposito F, Napolitano L, Di Bello F, Fusco A, et al. Differential Expression of LncRNA in Bladder Cancer Development. Diagnostics. 2023; 13(10):1745. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics13101745
Chicago/Turabian StyleSpirito, Lorenzo, Rufina Maturi, Sara Carmela Credendino, Celeste Manfredi, Davide Arcaniolo, Marco De Martino, Francesco Esposito, Luigi Napolitano, Francesco Di Bello, Alfredo Fusco, and et al. 2023. "Differential Expression of LncRNA in Bladder Cancer Development" Diagnostics 13, no. 10: 1745. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics13101745