
Diagnostic Utility of Immunohistochemical Detection of MEOX2, SOX11, INSM1 and EGFR in Gliomas

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Selection of Cases
2.2. Tissue Handling and Construction of Tissue Microarrays
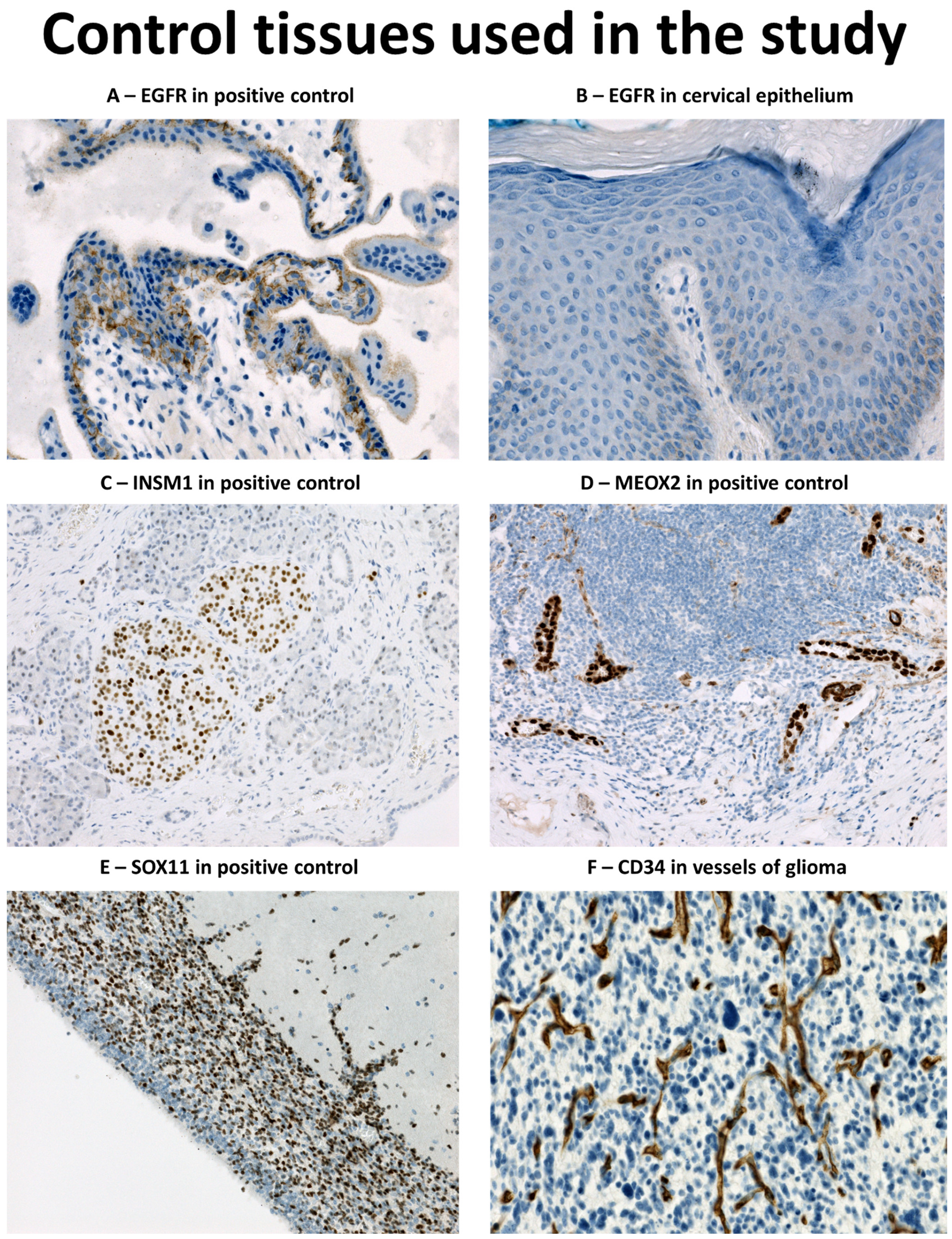
2.3. Immunohistochemical Analysis and In Situ Hybridisation (ISH) for EGFR
2.4. Morphological Evaluation
2.5. Statistical Analysis
3. Results
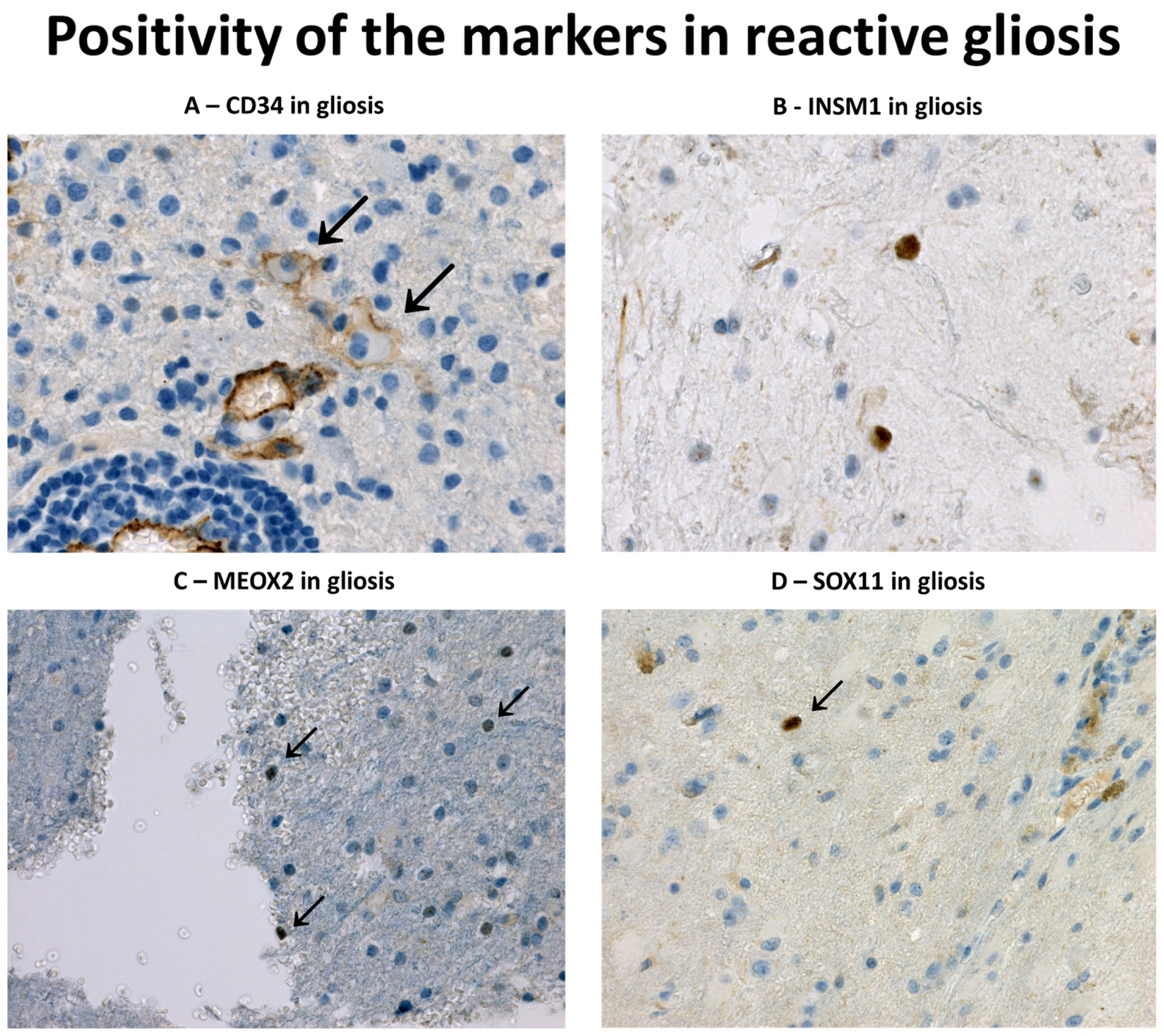
3.1. Immunohistochemical Profile of Reactive Gliosis
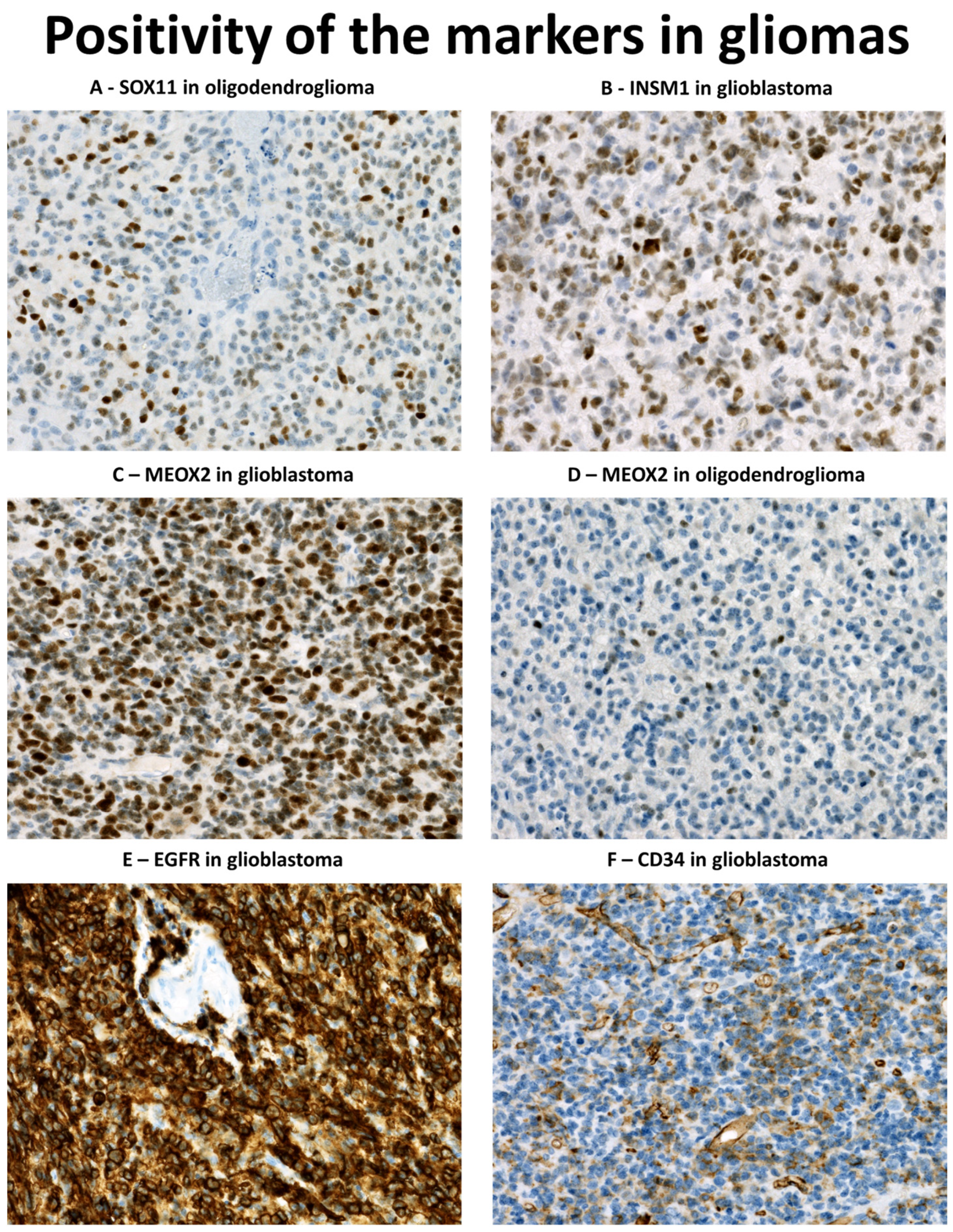
3.2. Immunohistochemical Profile of Diffuse Gliomas with IDH1 Mutation
3.3. Immunohistochemical Profile of Glioblastomas
3.4. Immunohistochemical Profile of Non-Diffuse Glial Tumours in the Study
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Capper, D.; Sahm, F.; Hartmann, C.; Meyermann, R.; von Deimling, A.; Schittenhelm, J. Application of Mutant IDH1 Antibody to Differentiate Diffuse Glioma From Nonneoplastic Central Nervous System Lesions and Therapy-induced Changes. Am. J. Surg. Pathol. 2010, 34, 1199–1204. [Google Scholar] [CrossRef]
- Hartmann, C.; Meyer, J.; Balss, J.; Capper, D.; Mueller, W.; Christians, A.; Felsberg, J.; Wolter, M.; Mawrin, C.; Wick, W.; et al. Type and frequency of IDH1 and IDH2 mutations are related to astrocytic and oli-godendroglial differentiation and age: A study of 1010 diffuse gliomas. Acta Neuropathol. 2009, 118, 469–474. [Google Scholar] [CrossRef] [Green Version]
- Balss, J.; Meyer, J.; Mueller, W.; Korshunov, A.; Hartmann, C.; von Deimling, A. Analysis of the IDH1 codon 132 mutation in brain tumors. Acta Neuropathol. 2008, 116, 597–602. [Google Scholar] [CrossRef] [PubMed]
- Parsons, D.W.; Jones, S.; Zhang, X.; Lin, J.C.-H.; Leary, R.J.; Angenendt, P.; Mankoo, P.; Carter, H.; Siu, I.-M.; Gallia, G.L.; et al. An integrated enomic analysis of human glioblastoma multiforme. Science 2008, 321, 1807–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glio-blastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncology 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Tanboon, J.; Williams, E.A.; Louis, D.N. The Diagnostic Use of Immunohistochemical Surrogates for Signature Molecular Genetic Alterations in Gliomas. J. Neuropathol. Exp. Neurol. 2015, 75, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Yaziji, H.; Massarani-Wafai, R.; Gujrati, M.; Kuhns, J.G.; Martin, A.W.; Parker, J.C. Role of p53 Immunohistochemistry in Differentiating Reactive Gliosis From Malignant Astrocytic Lesions. Am. J. Surg. Pathol. 1996, 20, 1086–1090. [Google Scholar] [CrossRef]
- Pastrana, E.; Cheng, L.-C.; Doetsch, F. Simultaneous prospective purification of adult subventricular zone neural stem cells and their progeny. Proc. Natl. Acad. Sci. USA 2009, 106, 6387–6392. [Google Scholar] [CrossRef]
- Eskilsson, E.; Røsland, G.V.; Solecki, G.; Wang, Q.; Harter, P.N.; Graziani, G.; Verhaak, R.G.W.; Winkler, F.; Bjerkvig, R.; Miletic, H. EGFR heterogeneity and implications for therapeutic intervention in glioblastoma. Neuro-Oncology 2017, 20, 743–752. [Google Scholar] [CrossRef] [Green Version]
- Tachon, G.; Masliantsev, K.; Rivet, P.; Petropoulos, C.; Godet, J.; Milin, S.; Wager, M.; Guichet, P.-O.; Karayan-Tapon, L. Prognostic significance of MEOX2 in gliomas. Mod. Pathol. 2019, 32, 774–786. [Google Scholar] [CrossRef] [PubMed]
- Tachon, G.; Masliantsev, K.; Rivet, P.; Desette, A.; Milin, S.; Gueret, E.; Wager, M.; Karayan-Tapon, L.; Guichet, P.-O. MEOX2 Transcription Factor Is Involved in Survival and Adhesion of Glioma Stem-like Cells. Cancers 2021, 13, 5943. [Google Scholar] [CrossRef] [PubMed]
- Duggan, A.; Madathany, T.; de Castro, S.C.; Gerrelli, D.; Guddati, K.; García-Añoveros, J. Transient expression of the conserved zinc finger gene INSM1 in progenitors and nascent neurons throughout embryonic and adult neurogenesis. J. Comp. Neurol. 2008, 507, 1497–1520. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, L.; Lai, H.; Parada, L.F.; Lei, L. Transcription factor Sox11 is essential for both embryonic and adult neurogenesis. Dev. Dyn. 2013, 242, 638–653. [Google Scholar] [CrossRef]
- Haslinger, A.; Schwarz, T.J.; Covic, M.; Lie, D.C. Expression of Sox11 in adult neurogenic niches suggests a stage-specific role in adult neurogenesis. Eur. J. Neurosci. 2009, 29, 2103–2114. [Google Scholar] [CrossRef]
- Ames, H.M.; Rooper, L.M.; Laterra, J.J.; Eberhart, C.G.; Rodriguez, F.J. INSM1 Expression Is Frequent in Primary Central Nervous System Neoplasms but Not in the Adult Brain Parenchyma. J. Neuropathol. Exp. Neurol. 2018, 77, 374–382. [Google Scholar] [CrossRef]
- Farkas, L.M.; Haffner, C.; Giger, T.; Khaitovich, P.; Nowick, K.; Birchmeier, C.; Pääbo, S.; Huttner, W.B. Insulinoma-Associated 1 Has a Panneurogenic Role and Promotes the Generation and Expansion of Basal Progenitors in the Developing Mouse Neocortex. Neuron 2008, 60, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Korkolopoulou, P.; Levidou, G.; El-Habr, E.A.; Adamopoulos, C.; Fragkou, P.; Boviatsis, E.; Themistocleous, M.S.; Petraki, K.; Vrettakos, G.; Sakalidou, M.; et al. Sox11 expression in astrocytic gliomas: Correlation with nes-tin/c-Met/IDH1-R132H expression phenotypes, p-Stat-3 and survival. Br. J. Cancer 2013, 108, 2142–2152. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Dong, Y.; Huo, Z.; Yu, L.; Xue, J.; Wang, G.; Duan, Y. SOX11: A potentially useful marker in surgical pathology: A systematic analysis of SOX11 expression in epithelial and non-epithelial tumours. Histopathology 2018, 74, 391–405. [Google Scholar] [CrossRef]
- Hide, T.; Takezaki, T.; Nakatani, Y.; Nakamura, H.; Kuratsu, J.I.; Kondo, T. Sox11 prevents tumorigenesis of glioma-initiating cells by inducing neuronal differ-entiation. Cancer Res. 2009, 69, 7953–7959. [Google Scholar] [CrossRef] [Green Version]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise Review: Evidence for CD34 as a Common Marker for Diverse Progenitors. Stem Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef]
- Blümcke, I.; Giencke, K.; Wardelmann, E.; Beyenburg, S.; Kral, T.; Sarioglu, N.; Pietsch, T.; Wolf, H.K.; Schramm, J.; Elger, C.E.; et al. The CD34 epitope is expressed in neoplastic and malformative lesions associated with chronic, focal epilepsies. Acta Neuropathol. 1999, 97, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Giulioni, M.; Marucci, G.; Cossu, M.; Tassi, L.; Bramerio, M.; Barba, C.; Buccoliero, A.M.; Vornetti, G.; Zenesini, C.; Consales, A.; et al. CD34 Expression in Low-Grade Epilepsy-Associated Tumors: Relationships with Clinicopathologic Features. World Neurosurg. 2018, 121, e761–e768. [Google Scholar] [CrossRef] [PubMed]
- Burel-Vandenbos, F.; Benchetrit, M.; Miquel, C.; Fontaine, D.; Auvergne, R.; Lebrun-Frenay, C.; Cardot-Leccia, N.; Michiels, J.-F.; Paquis-Flucklinger, V.; Virolle, T. EGFR immunolabeling pattern may discriminate low-grade gliomas from gliosis. J. Neuro-Oncol. 2010, 102, 171–178. [Google Scholar] [CrossRef]
- Conroy, S.; Kruyt, F.A.E.; Joseph, J.V.; Balasubramaniyan, V.; Bhat, K.P.; Wagemakers, M.; Enting, R.H.; Walenkamp, A.M.E.; Dunnen, W.F.A.D. Subclassification of Newly Diagnosed Glioblastomas through an Immunohistochemical Approach. PLoS ONE 2014, 9, e115687. [Google Scholar] [CrossRef]
- Guillaudeau, A.; Durand, K.; Pommepuy, I.; Robert, S.; Chaunavel, A.; Lacorre, S.; DeArmas, R.; Bourtoumieux, S.; El Demery, M.; Moreau, J.-J.; et al. Determination of EGFR status in gliomas: Usefulness of immunohistochem-istry and fluorescent in situ hybridization. Appl. Immunohistochem. Mol. Morphol. 2009, 17, 220–226. [Google Scholar] [CrossRef]
- Popova, S.N.; Bergqvist, M.; Dimberg, A.; Edqvist, P.-H.; Ekman, S.; Hesselager, G.; Ponten, F.; Smits, A.; Sooman, L.; Alafuzoff, I. Subtyping of gliomas of various WHO grades by the application of immuno-histochemistry. Histopathology 2014, 64, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Jiang, S.; Lu, C.; Ji, T.; Yang, W.; Li, T.; Lv, J.; Hu, W.; Yang, Y.; Jin, Z. SOX11: Friend or foe in tumor prevention and carcinogenesis? Ther. Adv. Med. Oncol. 2019, 11, 1758835919853449. [Google Scholar] [CrossRef] [Green Version]
- Lan, M.S.; Breslin, M.B. Structure, expression, and biological function of INSM1 transcription factor in neuroendocrine differentiation. FASEB J. 2009, 23, 2024–2033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, Z.S.; Zhang, C.B.; Wang, H.J.; Yan, W.; Liu, Y.W.; Li, M.Y.; Zhang, W. Whole-genome mRNA expression profiling identifies functional and prognostic sig-natures in patients with mesenchymal glioblastoma multiforme. CNS Neurosci. Ther. 2013, 19, 714–720. [Google Scholar] [CrossRef]
- Lohkamp, L.-N.; Schinz, M.; Gehlhaar, C.; Guse, K.; Thomale, U.-W.; Vajkoczy, P.; Heppner, F.L.; Koch, A. MGMT Promoter Methylation and BRAF V600E Mutations Are Helpful Markers to Discriminate Pleomorphic Xanthoastrocytoma from Giant Cell Glioblastoma. PLoS ONE 2016, 11, e0156422. [Google Scholar] [CrossRef] [Green Version]
- Galloway, M. CD34 expression in glioblastoma and giant cell glioblastoma. Clin. Neuropathol. 2010, 29, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Burel-Vandenbos, F.; Turchi, L.; Benchetrit, M.; Fontas, E.; Pedeutour, Z.; Rigau, V.; Almairac, F.; Ambrosetti, D.; Michiels, J.-F.; Virolle, T. Cells with intense EGFR staining and a high nuclear to cytoplasmic ratio are specific for infiltrative glioma: A useful marker in neuropathological practice. Neuro-Oncology 2013, 15, 1278–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hölzl, D.; Hutarew, G.; Zellinger, B.; Alinger-Scharinger, B.; Schlicker, H.U.; Schwartz, C.; Sotlar, K.; Kraus, T.F.J. EGFR Amplification Is a Phenomenon of IDH Wildtype and TERT Mutated High-Grade Glioma: An Integrated Analysis Using Fluorescence In Situ Hybridization and DNA Methylome Profiling. Biomedicines 2022, 10, 794. [Google Scholar] [CrossRef]
- Li, K.K.-W.; Shi, Z.-F.; Malta, T.M.; Chan, A.K.-Y.; Cheng, S.; Kwan, J.S.H.; Yang, R.R.; Poon, W.S.; Mao, Y.; Noushmehr, H.; et al. Identification of subsets of IDH-mutant glioblastomas with distinct epigenetic and copy number alterations and stratified clinical risks. Neuro-Oncol. Adv. 2019, 1. [Google Scholar] [CrossRef] [Green Version]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Sharain, R.F.; Gown, A.M.; Greipp, P.T.; Folpe, A.L. Immunohistochemistry for TFE3 lacks specificity and sensitivity in the diagnosis of TFE3-rearranged neoplasms: A comparative, 2-laboratory study. Hum. Pathol. 2019, 87, 65–74. [Google Scholar] [CrossRef]
- Teo, W.-Y.; Sekar, K.; Seshachalam, P.; Shen, J.; Chow, W.-Y.; Lau, C.C.; Yang, H.; Park, J.; Kang, S.-G.; Li, X.; et al. Relevance of a TCGA-derived Glioblastoma Subtype Gene-Classifier among Patient Populations. Sci. Rep. 2019, 9, 7442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swiss, V.A.; Nguyen, T.; Dugas, J.; Ibrahim, A.; Barres, B.; Androulakis, I.P.; Casaccia, P. Identification of a gene regulatory network necessary for the initiation of oligoden-drocyte differentiation. PLoS ONE 2011, 6, e18088. [Google Scholar] [CrossRef] [Green Version]
- Camacho-Urkaray, E.; Santos-Juanes, J.; Gutiérrez-Corres, F.B.; García, B.; Quirós, L.M.; Guerra-Merino, I.; Aguirre, J.J.; Fernández-Vega, I. Establishing cut-off points with clinical relevance for bcl-2, cyclin D1, p16, p21, p27, p53, Sox11 and WT1 expression in glioblastoma - a short report. Cell. Oncol. 2017, 41, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Vengoechea, J.; Zheng, S.; Sloan, A.E.; Chen, Y.; Brat, D.J.; O’neill, B.P.; de Groot, J.; Yust-Katz, S.; Yung, W.-K.A.; et al. Molecular Subtypes of Glioblastoma Are Relevant to Lower Grade Glioma. PLoS ONE 2014, 9, e91216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motomura, K.; Natsume, A.; Watanabe, R.; Ito, I.; Kato, Y.; Momota, H.; Nishikawa, R.; Mishima, K.; Nakasu, Y.; Abe, T.; et al. Immunohistochemical analysis-based proteomic subclassification of newly diagnosed glioblastomas. Cancer Sci. 2012, 103, 1871–1879. [Google Scholar] [CrossRef] [PubMed]
- McTigue, D.M.; Tripathi, R.B. The life, death, and replacement of oligodendrocytes in the adult CNS. J. Neurochem. 2008, 107, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lu, I.-N.; Dobersalske, C.; Rauschenbach, L.; Teuber-Hanselmann, S.; Steinbach, A.; Ullrich, V.; Prasad, S.; Blau, T.; Kebir, S.; Siveke, J.T.; et al. Tumor-associated hematopoietic stem and progenitor cells positively linked to glioblastoma progression. Nat. Commun. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Kovacs, M.; Trias, E.; Varela, V.; Ibarburu, S.; Beckman, J.S.; Moura, I.C.; Hermine, O.; King, P.H.; Si, Y.; Kwon, Y.; et al. CD34 Identifies a Subset of Proliferating Microglial Cells Associated with Degenerating Motor Neurons in ALS. Int. J. Mol. Sci. 2019, 20, 3880. [Google Scholar] [CrossRef] [Green Version]
- Soldini, D.; Valera, A.; Solé, C.; Palomero, J.; Amador, V.; Martin-Subero, J.I.; Ribera-Cortada, I.; Royo, C.; Salaverria, I.; Beà, S.; et al. Assessment of SOX11 expression in routine lymphoma tissue sections: Characterization of new monoclonal antibodies for diagnosis of mantle cell lymphoma. Am. J. Surg. Pathol. 2014, 38, 86–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torlakovic, E.E.; Nielsen, S.; Vyberg, M.; Taylor, C.R. Getting controls under control: The time is now for immunohistochemistry. J. Clin. Pathol. 2015, 68, 879–882. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Antibody | Clone | Dilution | Manufacturer | Pretreatment | Incubation Time | Visualisation | Positive Control |
|---|---|---|---|---|---|---|---|
| GFAP | EP672Y | RTU | Ventana, Basel, Switzerland | Ventana CC1, 36 min. | 16 min | Ventana ultraView | Brain |
| MEOX2 | 6A5 | 1:1000 | Sigma Aldrich s.r.o., Prague, CR | Ventana CC1, 56 min. | 32 min | Ventana OptiView | Appendix—HEV * |
| EGFR | 31G7 | 1:100 | Novus Biologicals, Ontario, Canada | Protease 3, 32 min | 32 min | Ventana OptiView | Placenta |
| SOX11 | MRQ-58 | 1:50 | Cell Marque, Rocklin, CA, USA | Ventana CC1, 56 min. | 32 min | Ventana OptiView | Foetal brain—subventricular zone |
| INSM1 | A-8 | 1:100 | SCBT Inc., Dallas, TX, USA | Dako PT Link High pH, 20 min. | 30 min | DAKO EnVision FLEX | Pancreas |
| CD34 | QBEnd10 | 1:50 | Cell Marque, Rocklin, CA, USA | Ventana CC1, 20 min. | 28 min | Ventana ultraView | Appendix |
| Gliosis | Gliomas | p | ||||||
|---|---|---|---|---|---|---|---|---|
| GBM | IDH1mt AA | OO | PA | EP | GG | |||
| INSM1 | 2.3% (1/44) | 81.6% (62/76) | 35.3% (6/17) | 82.4% (14/17) | 12.5% (1/8) | 0% (0/10) | 80% (4/5) | <0.001 (χ2) |
| EGFR | 0% (0/44) | 50% (39/78) | 23.5% (4/17) | 17.6% (3/17) | 0% (0/8) | 0% (0/10) | 0% (0/5) | <0.001 (χ2) |
| MEOX2 | 2.3% (1/43) | 81% (64/79) | 5.9% (1/17) | 5.9% (1/17) | 0% (0/8) | 0% (0/10) | 20% (1/5) | <0.001 (χ2) |
| SOX11 | 20.4% (9/44) | 75% (57/76) | 100% (17/17) | 100% (17/17) | 12.5% (1/8) | 10% (1/10) | 0% (0/4) | <0.001 (χ2) |
| CD34 | 11.6% (5/43) | 25% (20/80) | 0% (0/17) | 0% (0/17) | 12.5% (1/8) | 20% (2/10) | 60% (3/5) | 0.39 (χ2) |
| Gliomas | ||||
|---|---|---|---|---|
| GBM | IDH1mt AA | OO | GG | |
| INSM1 | 9 (2–23) | 0 (0–0) | 13.5 (±18.2) | 3.9 (±3.5) |
| EGFR | 0 (0–93.8) | 0 (0–0) | 0 (0–0) | 0 (0–0) |
| MEOX2 | 7.25 (1–70.6) | 0 (0–0) | 0 (0–0) | 0 (0–0) |
| SOX11 | 2 (0–17.9) | 22.5 (2.5–45) | 34.3 (±29.9) | 0 (0–0) |
| CD34 | 0 (0–1) | 0 (0-0) | 0 (0-0) | 7.4 (±10.5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soukup, J.; Gerykova, L.; Rachelkar, A.; Hornychova, H.; Bartos, M.C.; Krupa, P.; Vitovcova, B.; Pleskacova, Z.; Kasparova, P.; Dvorakova, K.; et al. Diagnostic Utility of Immunohistochemical Detection of MEOX2, SOX11, INSM1 and EGFR in Gliomas. Diagnostics 2023, 13, 2546. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics13152546
Soukup J, Gerykova L, Rachelkar A, Hornychova H, Bartos MC, Krupa P, Vitovcova B, Pleskacova Z, Kasparova P, Dvorakova K, et al. Diagnostic Utility of Immunohistochemical Detection of MEOX2, SOX11, INSM1 and EGFR in Gliomas. Diagnostics. 2023; 13(15):2546. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics13152546
Chicago/Turabian StyleSoukup, Jiri, Lucie Gerykova, Anjali Rachelkar, Helena Hornychova, Michael Christian Bartos, Petr Krupa, Barbora Vitovcova, Zuzana Pleskacova, Petra Kasparova, Katerina Dvorakova, and et al. 2023. "Diagnostic Utility of Immunohistochemical Detection of MEOX2, SOX11, INSM1 and EGFR in Gliomas" Diagnostics 13, no. 15: 2546. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics13152546