
Histological Profiling of the Human Umbilical Cord: A Potential Alternative Cell Source in Tissue Engineering

 , ,
, ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Situ Histological and Histochemical Characterization of the Human Umbilical Cord
2.2. Generation of Primary Ex Vivo Cultures of Human Umbilical Cord Mesenchymal Stem Cells (hUC-MSC)
2.3. In Situ Immunohistochemical Characterization of the Human Umbilical Cord
2.4. Ex Vivo Immunofluorescence Analysis of hUC-MSC
2.5. Quantification and Statistical Analysis
3. Results
3.1. In Situ Analysis of hUC Histology and ECM Histochemistry
3.2. Immunohistochemical Analysis of MSC Markers in hUC and Isolated hUC-MSC
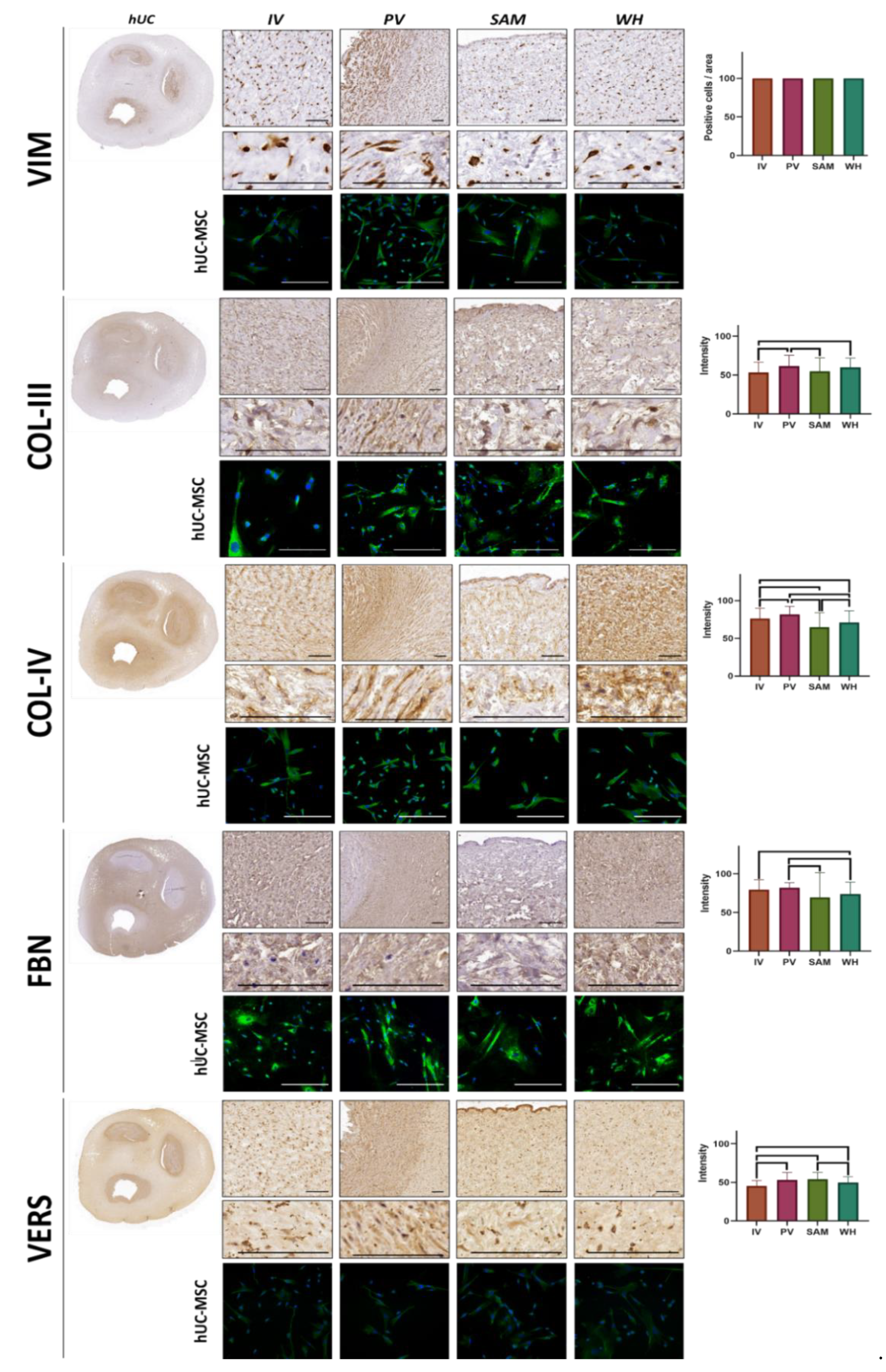
3.3. Immunohistochemical Analysis of Vimentin and ECM Components in hUC and Isolated hUC-MSC
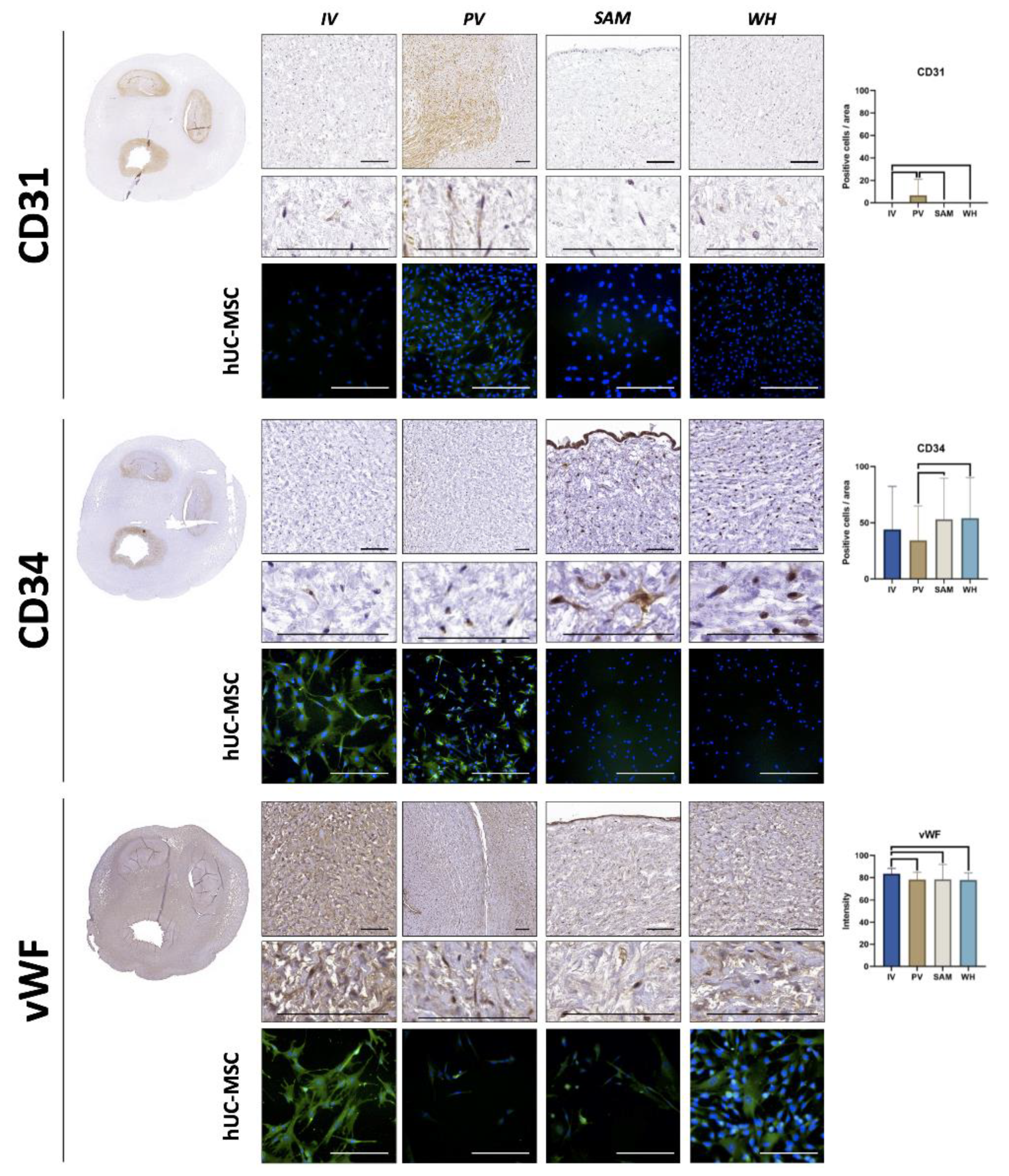
3.4. Immunohistochemical Analysis of Vascular Cell Markers in hUC and Isolated hUC-MSC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B.; et al. Perinatal Derivatives: Where Do We Stand? A Roadmap of the Human Placenta and Consensus for Tissue and Cell Nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 610544. [Google Scholar] [CrossRef] [PubMed]
- Gervaso, F.; Boschetti, F.; Pennati, G. Evaluation of the Wharton’s Jelly Poroelastic Parameters through Compressive Tests on Placental and Foetal Ends of Human Umbilical Cords. J. Mech. Behav. Biomed. Mater. 2014, 35, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Fong, C.-Y.; Biswas, A.; Bongso, A. Comparative Characterization of Cells from the Various Compartments of the Human Umbilical Cord Shows That the Wharton’s Jelly Compartment Provides the Best Source of Clinically Utilizable Mesenchymal Stem Cells. PLoS ONE 2015, 10, e0127992. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, J.; Wang, S.; Deng, Q.; Wang, K.; Dai, X.; An, Y.; Dong, G.; Ke, W.; Chen, F.; et al. Single-Cell Transcriptome Profiling Reveals Molecular Heterogeneity in Human Umbilical Cord Tissue and Culture-Expanded Mesenchymal Stem Cells. FEBS J. 2021, 288, 5311–5330. [Google Scholar] [CrossRef]
- Nanaev, A.K.; Kohnen, G.; Milovanov, A.P.; Domogatsky, S.P.; Kaufmann, P. Stromal Differentiation and Architecture of the Human Umbilical Cord. Placenta 1997, 18, 53–64. [Google Scholar] [CrossRef]
- Garzón, I.; Alfonso-Rodríguez, C.A.; Martínez-Gómez, C.; Carriel, V.; Martin-Piedra, M.A.; Fernández-Valadés, R.; Sánchez-Quevedo, M.C.; Alaminos, M. Expression of Epithelial Markers by Human Umbilical Cord Stem Cells. A Topographical Analysis. Placenta 2014, 35, 994–1000. [Google Scholar] [CrossRef]
- Bharti, D.; Shivakumar, S.B.; Park, J.-K.; Ullah, I.; Subbarao, R.B.; Park, J.-S.; Lee, S.-L.; Park, B.-W.; Rho, G.-J. Comparative Analysis of Human Wharton’s Jelly Mesenchymal Stem Cells Derived from Different Parts of the Same Umbilical Cord. Cell Tissue Res. 2018, 372, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.-C.; Chang, Y.-H.; Shyu, W.-C.; Lin, S.-Z. Human Umbilical Cord Mesenchymal Stem Cells: A New Era for Stem Cell Therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef]
- Kassem, D.H.; Kamal, M.M. Therapeutic Efficacy of Umbilical Cord-Derived Stem Cells for Diabetes Mellitus: A Meta-Analysis Study. Stem Cell Res. Ther. 2020, 11, 484. [Google Scholar] [CrossRef]
- Bae, J.-Y.; Choi, S.J.; Kim, J.K. Comparison of Mesenchymal Stem Cell Attachment Efficiency in Acellular Neural Graft for Peripheral Nerve Regeneration. J. Plast. Reconstr. Aesthetic Surg. JPRAS 2021, in press. [Google Scholar] [CrossRef]
- Chen, C.-F.; Chen, Y.-C.; Fu, Y.-S.; Tsai, S.-W.; Wu, P.-K.; Chen, C.-M.; Chang, M.-C.; Chen, W.-M. Characterization of Osteogenesis and Chondrogenesis of Human Decellularized Allogeneic Bone with Mesenchymal Stem Cells Derived from Bone Marrow, Adipose Tissue, and Wharton’s Jelly. Int. J. Mol. Sci. 2021, 22, 8987. [Google Scholar] [CrossRef] [PubMed]
- Garzón, I.; Miyake, J.; González-Andrades, M.; Carmona, R.; Carda, C.; del Sánchez-Quevedo, M.C.; Campos, A.; Alaminos, M. Wharton’s Jelly Stem Cells: A Novel Cell Source for Oral Mucosa and Skin Epithelia Regeneration. Stem Cells Transl. Med. 2013, 2, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Garzón, I.; Chato-Astrain, J.; González-Gallardo, C.; Ionescu, A.; de la Cardona, J.C.; Mateu, M.; Carda, C.; Pérez, M.D.M.; Martín-Piedra, M.Á.; Alaminos, M. Long-Term in Vivo Evaluation of Orthotypical and Heterotypical Bioengineered Human Corneas. Front. Bioeng. Biotechnol. 2020, 8, 681. [Google Scholar] [CrossRef] [PubMed]
- Alaminos, M.; Pérez-Köhler, B.; Garzón, I.; García-Honduvilla, N.; Romero, B.; Campos, A.; Buján, J. Transdifferentiation Potentiality of Human Wharton’s Jelly Stem Cells towards Vascular Endothelial Cells. J. Cell. Physiol. 2010, 223, 640–647. [Google Scholar] [CrossRef]
- Marino, L.; Castaldi, M.A.; Rosamilio, R.; Ragni, E.; Vitolo, R.; Fulgione, C.; Castaldi, S.G.; Serio, B.; Bianco, R.; Guida, M.; et al. Mesenchymal Stem Cells from the Wharton’s Jelly of the Human Umbilical Cord: Biological Properties and Therapeutic Potential. Int. J. Stem Cells 2019, 12, 218–226. [Google Scholar] [CrossRef]
- Liau, L.L.; Ruszymah, B.H.I.; Ng, M.H.; Law, J.X. Characteristics and Clinical Applications of Wharton’s Jelly-Derived Mesenchymal Stromal Cells. Curr. Res. Transl. Med. 2020, 68, 5–16. [Google Scholar] [CrossRef]
- Safari, F.; Fani, N.; Eglin, D.; Alini, M.; Stoddart, M.J.; Baghaban Eslaminejad, M. Human Umbilical Cord-Derived Scaffolds for Cartilage Tissue Engineering. J. Biomed. Mater. Res. A 2019, 107, 1793–1802. [Google Scholar] [CrossRef]
- Semenova, E.; Grudniak, M.P.; Machaj, E.K.; Bocian, K.; Chroscinska-Krawczyk, M.; Trochonowicz, M.; Stepaniec, I.M.; Murzyn, M.; Zagorska, K.E.; Boruczkowski, D.; et al. Mesenchymal Stromal Cells from Different Parts of Umbilical Cord: Approach to Comparison & Characteristics. Stem Cell Rev. Rep. 2021, 17, 1780–1795. [Google Scholar] [CrossRef]
- Carriel, V.; Garzón, I.; Campos, A.; Cornelissen, M.; Alaminos, M. Differential Expression of GAP-43 and Neurofilament during Peripheral Nerve Regeneration through Bio-Artificial Conduits. J. Tissue Eng. Regen. Med. 2017, 11, 553–563. [Google Scholar] [CrossRef]
- Wilke, D.J.; Denier, J.P.; Khong, T.Y.; Mattner, T.W. Pressure and Flow in the Umbilical Cord. J. Biomech. 2018, 79, 78–87. [Google Scholar] [CrossRef]
- Kasiteropoulou, D.; Topalidou, A.; Downe, S. A Computational Fluid Dynamics Modelling of Maternal-Fetal Heat Exchange and Blood Flow in the Umbilical Cord. PLoS ONE 2020, 15, e0231997. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Batsali, A.K.; Kastrinaki, M.-C.; Papadaki, H.A.; Pontikoglou, C. Mesenchymal Stem Cells Derived from Wharton’s Jelly of the Umbilical Cord: Biological Properties and Emerging Clinical Applications. Curr. Stem Cell Res. Ther. 2013, 8, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Ranjbaran, H.; Abediankenari, S.; Mohammadi, M.; Jafari, N.; Khalilian, A.; Rahmani, Z.; Momeninezhad Amiri, M.; Ebrahimi, P. Wharton’s Jelly Derived-Mesenchymal Stem Cells: Isolation and Characterization. Acta Med. Iran. 2018, 56, 28–33. [Google Scholar]
- Satelli, A.; Li, S. Vimentin in Cancer and Its Potential as a Molecular Target for Cancer Therapy. Cell. Mol. Life Sci. CMLS 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Bolten, Z.T.; Wagner, D.R.; Hsieh, A.H. Deformability of Human Mesenchymal Stem Cells Is Dependent on Vimentin Intermediate Filaments. Ann. Biomed. Eng. 2017, 45, 1365–1374. [Google Scholar] [CrossRef] [Green Version]
- Song, H.-H.G.; Rumma, R.T.; Ozaki, C.K.; Edelman, E.R.; Chen, C.S. Vascular Tissue Engineering: Progress, Challenges, and Clinical Promise. Cell Stem Cell 2018, 22, 340–354. [Google Scholar] [CrossRef] [Green Version]
- Buján, J.; García-Honduvilla, N.; Bellón, J.M. Engineering Conduits to Resemble Natural Vascular Tissue. Biotechnol. Appl. Biochem. 2004, 39, 17–27. [Google Scholar] [CrossRef]
- Blanco-Elices, C.; Chato-Astrain, J.; Oyonarte, S.; Bermejo-Casares, F.; España-López, A.; Fernández-Valadés, R.; Sánchez-Quevedo, M.D.C.; Alaminos, M.; Martín-Piedra, M.A.; Garzón, I. Generation of a Novel Model of Bioengineered Human Oral Mucosa with Increased Vascularization Potential. J. Periodontal Res. 2021, 56, 1116–1131. [Google Scholar] [CrossRef]
- Amirsadeghi, A.; Jafari, A.; Eggermont, L.J.; Hashemi, S.-S.; Bencherif, S.A.; Khorram, M. Vascularization Strategies for Skin Tissue Engineering. Biomater. Sci. 2020, 8, 4073–4094. [Google Scholar] [CrossRef]
- Hu, M.; Zhang, H.; Liu, Q.; Hao, Q. Structural Basis for Human PECAM-1-Mediated Trans-Homophilic Cell Adhesion. Sci. Rep. 2016, 6, 38655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusztaszeri, M.P.; Seelentag, W.; Bosman, F.T. Immunohistochemical Expression of Endothelial Markers CD31, CD34, von Willebrand Factor, and Fli-1 in Normal Human Tissues. J. Histochem. Cytochem. 2006, 54, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naß, J.; Terglane, J.; Gerke, V. Weibel Palade Bodies: Unique Secretory Organelles of Endothelial Cells That Control Blood Vessel Homeostasis. Front. Cell Dev. Biol. 2021, 9, 813995. [Google Scholar] [CrossRef] [PubMed]
- Garzon, I.; Chato-Astrain, J.; Campos, F.; Fernandez-Valades, R.; Sanchez-Montesinos, I.; Campos, A.; Alaminos, M.; D’Souza, R.N.; Martin-Piedra, M.A. Expanded Differentiation Capability of Human Wharton’s Jelly Stem Cells Toward Pluripotency: A Systematic Review. Tissue Eng. Part B Rev. 2020, 26, 301–312. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IV | PV | SAM | WH | IV vs. PV | IV vs. SAM | IV vs. WH | PV vs. SAM | PV vs. WH | SAM vs. WH | |
|---|---|---|---|---|---|---|---|---|---|---|
| AB | 88.44 ± 28.69 | 82.08 ± 24.38 | 76.08 ± 23.83 | 83.37 ± 24.84 | 0.1330 | 0.0012 | 0.0822 | 0.0974 | 0.5827 | 0.0754 |
| PAS | 58.96 ± 30.41 | 69.89 ± 32.66 | 52.98 ± 29.03 | 55.88 ± 26.77 | 0.0035 | 0.0837 | 0.5050 | <0.0001 | 0.0013 | 0.3365 |
| PR | 123.79 ± 26.74 | 125.89 ± 30.24 | 91.44 ± 27.74 | 99.06 ± 21.76 | 0.3616 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0296 |
| CD73 | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 | 1.0000 | 1.0000 | 1.0000 | 1.0000 | 1.0000 | 1.0000 |
| CD90 | 33.4 ± 35.41 | 14.42 ± 20.78 | 73.97 ± 22.14 | 36.95 ± 33.2 | 0.0010 | <0.0001 | 0.4978 | <0.0001 | <0.0001 | <0.0001 |
| CD105 | 4.18 ± 11.41 | 6.15 ± 16.53 | 1.79 ± 8.73 | 3.86 ± 9.69 | 0.5498 | 0.3032 | 0.9238 | 0.0963 | 0.6164 | 0.2514 |
| VIM | 100 ± 0 | 100 ± 0 | 100 ± 0 | 100 ± 0 | 1.0000 | 1.0000 | 1.0000 | 1.0000 | 1.0000 | 1.0000 |
| COL-III | 53.32 ± 13.38 | 61.39 ± 14.08 | 54.69 ± 17.41 | 59.95 ± 11.8 | 0.0001 | 0.5594 | 0.0064 | 0.0074 | 0.0721 | 0.0546 |
| COL-IV | 76.35 ± 13.46 | 81.8 ± 10.49 | 64.71 ± 19.33 | 70.99 ± 15.59 | 0.0332 | <0.0001 | 0.0238 | <0.0001 | <0.0001 | 0.0127 |
| FBN | 79.63 ± 12.88 | 82.12 ± 6.49 | 69.48 ± 32.11 | 73.76 ± 15.6 | 0.8967 | 0.0564 | 0.0059 | 0.0177 | 0.0016 | 0.2123 |
| VERS | 45.23 ± 7.13 | 52.88 ± 9.76 | 53.92 ± 8.96 | 49.69 ± 7.67 | <0.0001 | <0.0001 | 0.0001 | 0.0744 | 0.0997 | 0.0007 |
| CD31 | 0 ± 0 | 8.03 ± 17.34 | 0 ± 0 | 0 ± 0 | 0.0203 | 0.9989 | 0.9989 | 0.0203 | 0.0203 | 0.9989 |
| CD34 | 43.93 ± 38.43 | 34.3 ± 30.62 | 53.05 ± 36.53 | 67.86 ± 136.18 | 0.1670 | 0.2683 | 0.0858 | 0.0012 | 0.0004 | 0.6613 |
| vWF | 83.52 ± 4.84 | 78.19 ± 6.57 | 78.38 ± 13.6 | 77.88 ± 6.37 | <0.0001 | 0.0304 | <0.0001 | 0.2965 | 0.6144 | 0.1806 |
| IV | PV | SAM | WH | |
|---|---|---|---|---|
| VIM | ++ | ++ | ++ | ++ |
| COL-III | +++ | +++ | +++ | +++ |
| COL-IV | ++ | ++ | ++ | ++ |
| FBN | +++ | +++ | +++ | +++ |
| VERS | + | + | + | + |
| CD31 | − | +/− | − | − |
| CD34 | ++ | ++ | − | − |
| vWF | +++ | + | + | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco-Elices, C.; Chato-Astrain, J.; González-González, A.; Sánchez-Porras, D.; Carriel, V.; Fernández-Valadés, R.; Sánchez-Quevedo, M.d.C.; Alaminos, M.; Garzón, I. Histological Profiling of the Human Umbilical Cord: A Potential Alternative Cell Source in Tissue Engineering. J. Pers. Med. 2022, 12, 648. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm12040648
Blanco-Elices C, Chato-Astrain J, González-González A, Sánchez-Porras D, Carriel V, Fernández-Valadés R, Sánchez-Quevedo MdC, Alaminos M, Garzón I. Histological Profiling of the Human Umbilical Cord: A Potential Alternative Cell Source in Tissue Engineering. Journal of Personalized Medicine. 2022; 12(4):648. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm12040648
Chicago/Turabian StyleBlanco-Elices, Cristina, Jesús Chato-Astrain, Alberto González-González, David Sánchez-Porras, Víctor Carriel, Ricardo Fernández-Valadés, María del Carmen Sánchez-Quevedo, Miguel Alaminos, and Ingrid Garzón. 2022. "Histological Profiling of the Human Umbilical Cord: A Potential Alternative Cell Source in Tissue Engineering" Journal of Personalized Medicine 12, no. 4: 648. https://0-doi-org.brum.beds.ac.uk/10.3390/jpm12040648