Inhibitors of Eicosanoid Biosynthesis Reveal that Multiple Lipid Signaling Pathways Influence Malaria Parasite Survival in Anopheles gambiae


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Rearing
2.2. Eicosanoid Inhibitors
2.3. Plasmodium berghei Infections
2.4. Gene-Silencing Experiments
2.5. Gene Expression Analysis
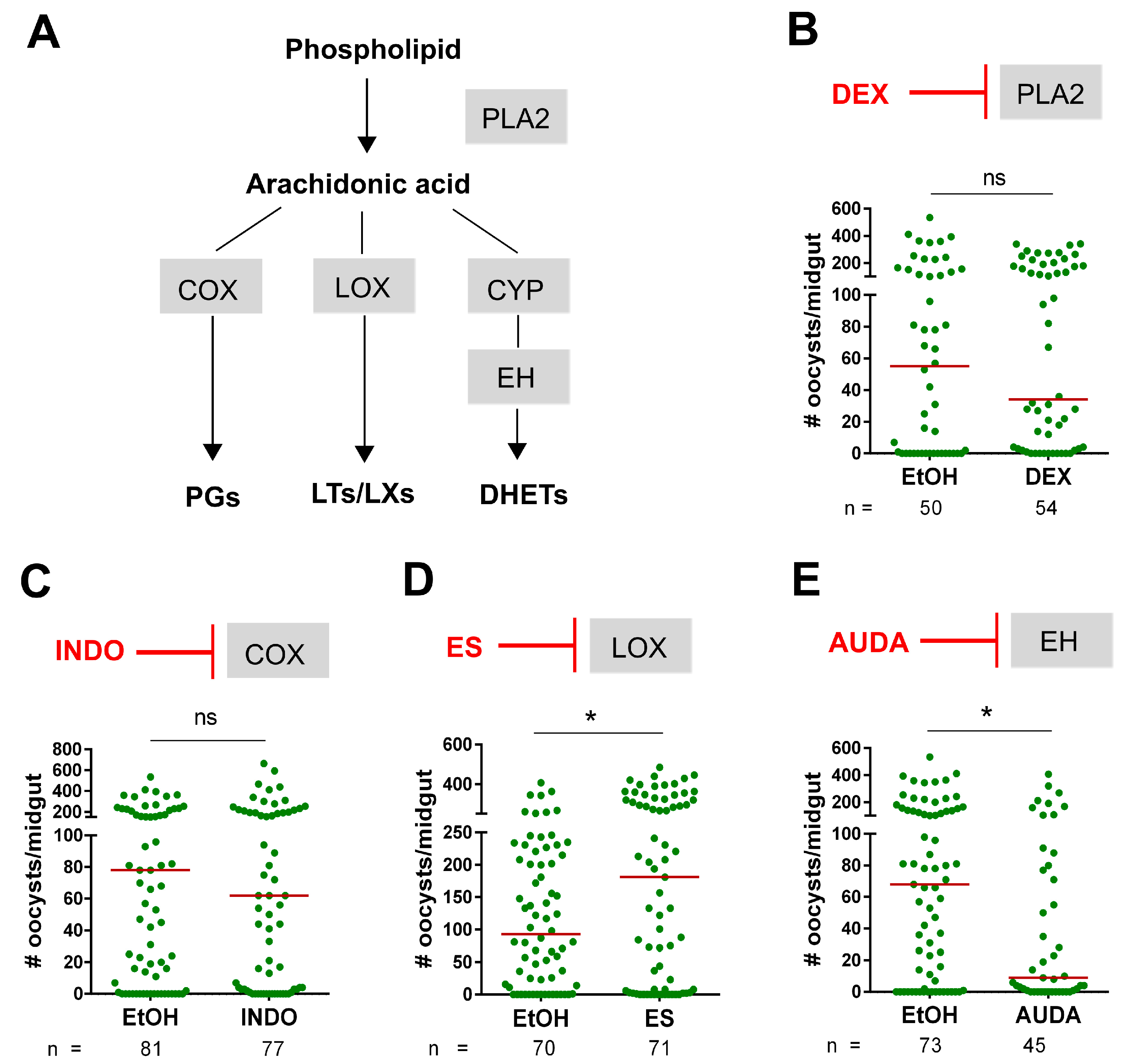
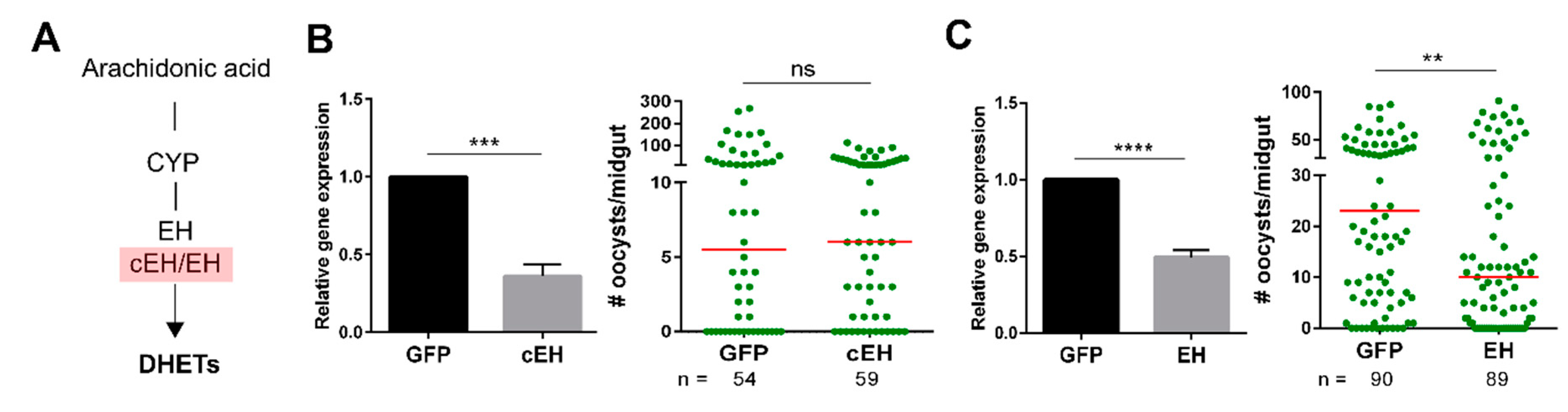
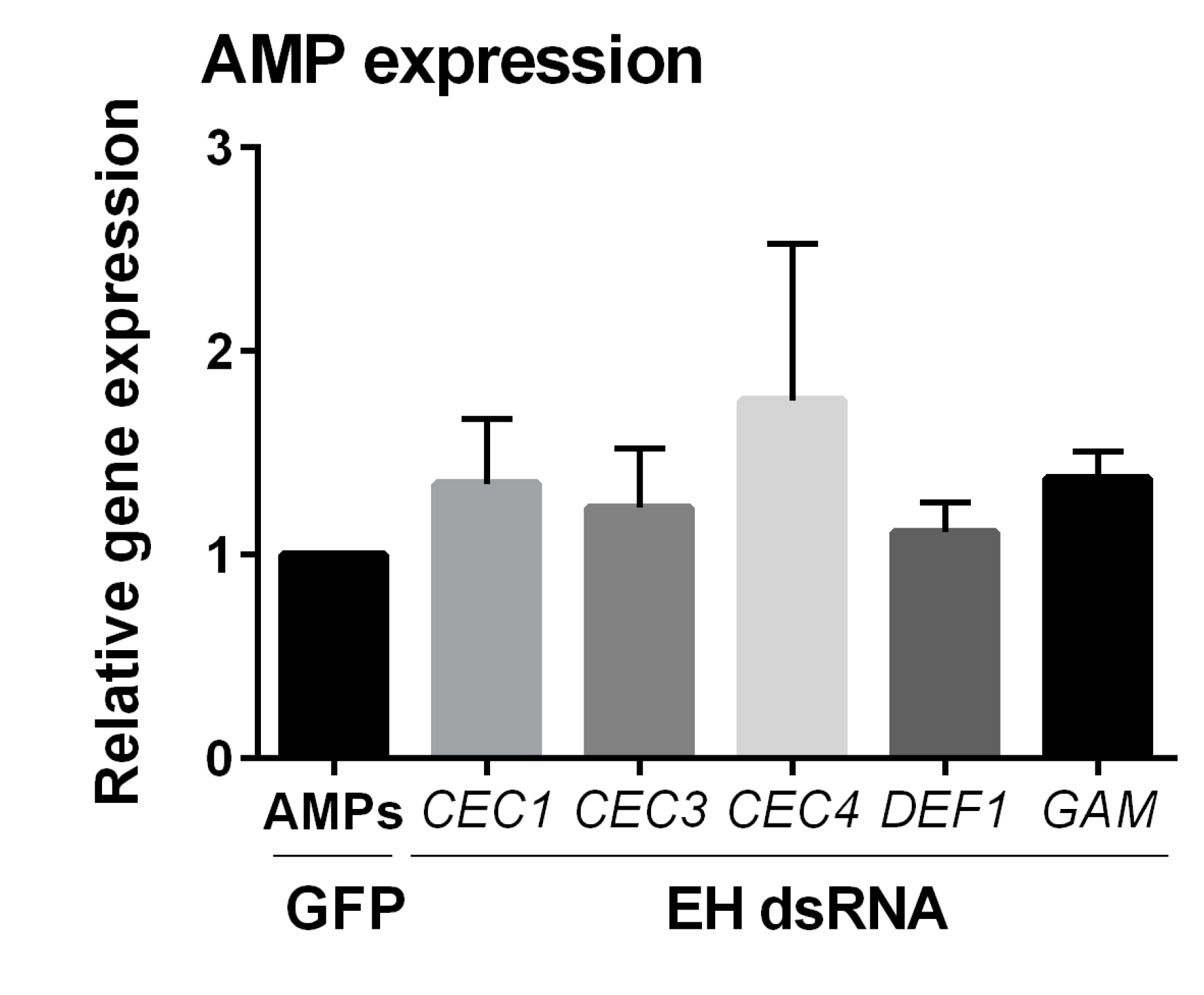
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jansen, C.C.; Beebe, N.W. The dengue vector Aedes aegypti: What comes next. Microb. Infect. 2010, 12, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Ashley, E.A.; Pyae Phyo, A.; Woodrow, C.J. Malaria. Lancet 2018, 391, 1608–1621. [Google Scholar] [CrossRef]
- World Malaria Report 2018; World Health Organization: Geneva, Switzerland, 2018; ISBN 978-92-4-156565-3.
- Smith, R.C.; Vega-Rodríguez, J.; Jacobs-Lorena, M. The Plasmodium bottleneck: Malaria parasite losses in the mosquito vector. Mem. Inst. Oswaldo. Cruz. 2014, 109, 644–661. [Google Scholar] [CrossRef] [PubMed]
- Tse, E.G.; Korsik, M.; Todd, M.H. The past, present and future of anti-malarial medicines. Malar J. 2019, 18, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.C.; Barillas-Mury, C. Plasmodium Oocysts: Overlooked targets of mosquito immunity. Trends Parasitol. 2016, 32, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Blandin, S.; Shiao, S.H.; Moita, L.F.; Janse, C.J.; Waters, A.P.; Kafatos, F.C.; Levashina, E.A. Complement-like protein TEP1 is a determinant of vectorial capacity in the malaria vector Anopheles gambiae. Cell 2004, 116, 661–670. [Google Scholar] [CrossRef]
- Fraiture, M.; Baxter, R.H.G.; Steinert, S.; Chelliah, Y.; Frolet, C.; Quispe-Tintaya, W.; Hoffmann, J.A.; Blandin, S.A.; Levashina, E.A. Two mosquito LRR proteins function as complement control factors in the TEP1-mediated killing of Plasmodium. Cell Host Microbe. 2009, 5, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Povelones, M.; Waterhouse, R.M.; Kafatos, F.C.; Christophides, G.K. Leucine-rich repeat protein complex activates mosquito complement in defense against Plasmodium parasites. Science 2009, 324, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Gupta, L.; Molina-Cruz, A.; Kumar, S.; Rodrigues, J.; Dixit, R.; Zamora, R.E.; Barillas-Mury, C. The STAT pathway mediates late-phase immunity against Plasmodium in the mosquito Anopheles gambiae. Cell Host Microbe. 2009, 5, 498–507. [Google Scholar] [CrossRef]
- Smith, R.C.; Barillas-Mury, C.; Jacobs-Lorena, M. Hemocyte differentiation mediates the mosquito late-phase immune response against Plasmodium in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2015, 112, E3412–E3420. [Google Scholar] [CrossRef]
- Kwon, H.; Arends, B.R.; Smith, R.C. Late-phase immune responses limiting oocyst survival are independent of TEP1 function yet display strain specific differences in Anopheles gambiae. Parasit. Vectors 2017, 10, 369. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Smith, R.C. Chemical depletion of phagocytic immune cells in Anopheles gambiae reveals dual roles of mosquito hemocytes in anti-Plasmodium immunity. Proc. Natl. Acad. Sci. USA 2019, 116, 14119–14128. [Google Scholar] [CrossRef]
- Oliveira, G.D.A.; Lieberman, J.; Barillas-Mury, C. Epithelial nitration by a Peroxidase/NOX5 system mediates mosquito antiplasmodial immunity. Science 2012, 335, 856–859. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Thompson, J.; Kafatos, F.C.; Barillas-Mury, C. Molecular interactions between Anopheles stephensi midgut cells and Plasmodium berghei: The time bomb theory of ookinete invasion. EMBO J. 2000, 19, 6030–6040. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.C.; Beatriz, A.; Ferreira, B.; Trisnadi, N.; Barillas-Mury, C. Activation of mosquito complement antiplasmodial response requires cellular immunity. Sci. Immunol. 2017, 2, eaal1505. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.G.; Padilla, J.; Koumas, L.; Ray, D.; Phipps, R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002, 23, 144–150. [Google Scholar] [CrossRef]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley-Samuelson, D.W.; Peloquin, J.J.; Loher, W. Egg-laying in response to prostaglandin injections in the Australian field cricket, Teleogryllus commodus. Physiol. Entomol. 1986, 11, 213–219. [Google Scholar] [CrossRef]
- Petzel, D.H.; Parrish, A.K.; Ogg, C.L.; Witters, N.A.; Howard, R.W.; Stanley-Samuelson, D.W. Arachidonic acid and prostaglandin E2 in malpighian tubules of female yellow fever mosquitoes. Insect Biochem. Mol. Biol. 1993, 23, 431–437. [Google Scholar] [CrossRef]
- Howard, R.W.; Stanley-Samuelson, D.W. Fatty acid composition of fat body and malpighian tubules of the tenebrionid beetle, Zophobas atratus: Significance in eicosanoid-mediated physiology. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1996, 115, 429–437. [Google Scholar] [CrossRef]
- Stanley, D.; Kim, Y. Prostaglandins and their receptors in insect biology. Front. Endocrinol. 2011, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; de Almeida Oliveira, G.; Calvo, E.; Dalli, J.; Colas, R.A.; Serhan, C.N.; Ribeiro, J.M.; Barillas-Mury, C. A mosquito lipoxin/lipocalin complex mediates innate immune priming in Anopheles gambiae. Nat. Commun. 2015, 6, 7403. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Stanley, D.; Kim, Y. An insect prostaglandin E2 synthase acts in immunity and reproduction. Front. Physiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ferreira Barletta, A.B.; Trisnadi, N.; Ramirez, J.L.; Barillas-Mury, C. Mosquito midgut prostaglandin release establishes systemic immune priming. Science 2019, 19, 54–62. [Google Scholar]
- Brune, K.; Patrignani, P. New insights into the use of currently available non-steroidal anti-inflammatory drugs. J. Pain Res. 2015, 8, 105. [Google Scholar] [CrossRef] [PubMed]
- Stanley-Samuelson, D.; Ogg, C. Prostaglandin biosynthesis by fat body from the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. 1994, 24, 481–491. [Google Scholar] [CrossRef]
- Shrestha, S.; Kim, Y. Eicosanoids mediate prophenoloxidase release from oenocytoids in the beet armyworm Spodoptera exigua. Insect Biochem. Mol. Biol. 2008, 38, 99–112. [Google Scholar] [CrossRef]
- Miller, J.S.; Nguyen, T.; Stanley-Samuelson, D.W. Eicosanoids mediate insect nodulation responses to bacterial infections. Proc. Natl. Acad. Sci. USA 1994, 91, 12418–12422. [Google Scholar] [CrossRef]
- Stanley, D.W.; Miller, J.S. Eicosanoid actions in insect cellular immune functions. Entomol. Exp. Appl. 2006, 119, 1–13. [Google Scholar] [CrossRef]
- Park, Y.; Stanley, D.W.; Kim, Y. Eicosanoids up-regulate production of reactive oxygen species by NADPH-dependent oxidase in Spodoptera exigua phagocytic hemocytes. J. Insect Physiol. 2015, 79, 63–72. [Google Scholar] [CrossRef]
- Park, J.A.; Kim, Y. Toll recognition signal activates oenocytoid cell lysis via a crosstalk between plasmatocyte-spreading peptide and eicosanoids in response to a fungal infection. Cell. Immunol. 2012, 279, 117–123. [Google Scholar] [CrossRef] [PubMed]
- García Gil de Muñoz, F.L.; Martínez-Barnetche, J.; Lanz-Mendoza, H.; Rodríguez, M.H.; Hernández-Hernández, F.C. Prostaglandin E2 modulates the expression of antimicrobial peptides in the fat body and midgut of Anopheles albimanus. Arch. Insect Biochem. Physiol. 2008, 68, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Carton, Y.; Frey, F.; Stanley, D.W.; Vass, E.; Nappi, A.J. Dexamethasone inhibition of the cellular immune response of Drosophila melanogaster against a parasitoid. J. Parasitol. 2002, 88, 405. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Morisseau, C.; Hammock, B.D. Expression and characterization of an epoxide hydrolase from Anopheles gambiae with high activity on epoxy fatty acids. Insect Biochem. Mol. Biol. 2014, 54, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Morisseau, C.; Yang, J.; Mamatha, D.M.; Hammock, B.D. Epoxide hydrolase activities and epoxy fatty acids in the mosquito Culex quinquefasciatus. Insect Biochem. Mol. Biol. 2015, 59, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Morisseau, C.; Yang, J.; Lee, K.S.S.; Kamita, S.G.; Hammock, B.D. Ingestion of the epoxide hydrolase inhibitor AUDA modulates immune responses of the mosquito, Culex quinquefasciatus during blood feeding. Insect Biochem. Mol. Biol. 2016, 76, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.J. Eicosanoid receptors: Targets for the treatment of disrupted intestinal epithelial homeostasis. Eur. J. Pharmacol. 2017, 796, 7–19. [Google Scholar] [CrossRef]
- Woodward, D.F.; Jones, R.L.; Narumiya, S. International Union of Basic and Clinical Pharmacology. LXXXIII: Classification of prostanoid receptors, updating 15 years of progress. Pharmacol. Rev. 2011, 63, 471–538. [Google Scholar] [CrossRef]
- Ramos, S.; Custódio, A.; Silveira, H. Anopheles gambiae eicosanoids modulate Plasmodium berghei survival from oocyst to salivary gland invasion. Mem. Inst. Oswaldo. Cruz. 2014, 109, 668–671. [Google Scholar] [CrossRef]
- Burke, J.E.; Dennis, E.A. Phospholipase A 2 structure/function, mechanism, and signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef] [PubMed]
- Merchant, D.; Ertl, R.L.; Rennard, S.I.; Stanley, D.W.; Miller, J.S. Eicosanoids mediate insect hemocyte migration. J. Insect Physiol. 2008, 54, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Park, Y.; Kim, Y.; Hwang, J.; Lee, D. An entomopathogenic bacterium, Xenorhabdus nematophila, suppresses expression of antimicrobial peptides controlled by Toll and Imd pathways by blocking eicosanoid biosynthesis. Arch. Insect Biochem. Physiol. 2013, 83, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Kim, Y. Activation of immune-associated phospholipase A2 is functionally linked to Toll/Imd signal pathways in the red flour beetle, Tribolium castaneum. Dev. Comp. Immunol. 2010, 34, 530–537. [Google Scholar] [CrossRef] [PubMed]
- Van Zyl, A.; Louw, A. Inhibition of peroxidase activity by some non-steroidal anti-inflammatory drugs. Biochem. Pharmacol. 1979, 28, 2753–2759. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, H.; Smith, R.C. Inhibitors of Eicosanoid Biosynthesis Reveal that Multiple Lipid Signaling Pathways Influence Malaria Parasite Survival in Anopheles gambiae. Insects 2019, 10, 307. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10100307
Kwon H, Smith RC. Inhibitors of Eicosanoid Biosynthesis Reveal that Multiple Lipid Signaling Pathways Influence Malaria Parasite Survival in Anopheles gambiae. Insects. 2019; 10(10):307. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10100307
Chicago/Turabian StyleKwon, Hyeogsun, and Ryan C. Smith. 2019. "Inhibitors of Eicosanoid Biosynthesis Reveal that Multiple Lipid Signaling Pathways Influence Malaria Parasite Survival in Anopheles gambiae" Insects 10, no. 10: 307. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10100307