The Influence of New Surveillance Data on Predictive Species Distribution Modeling of Aedes aegypti and Aedes albopictus in the United States


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Data Collection and Processing 2016–2017
2.2. Additional Ae. aegypti and Ae. albopictus Data
2.3. Bioclimatic Variables
2.4. Species Distribution Modeling Using MaxEnt in R Studio
2.5. Background Data
2.6. Covariate Selection Process
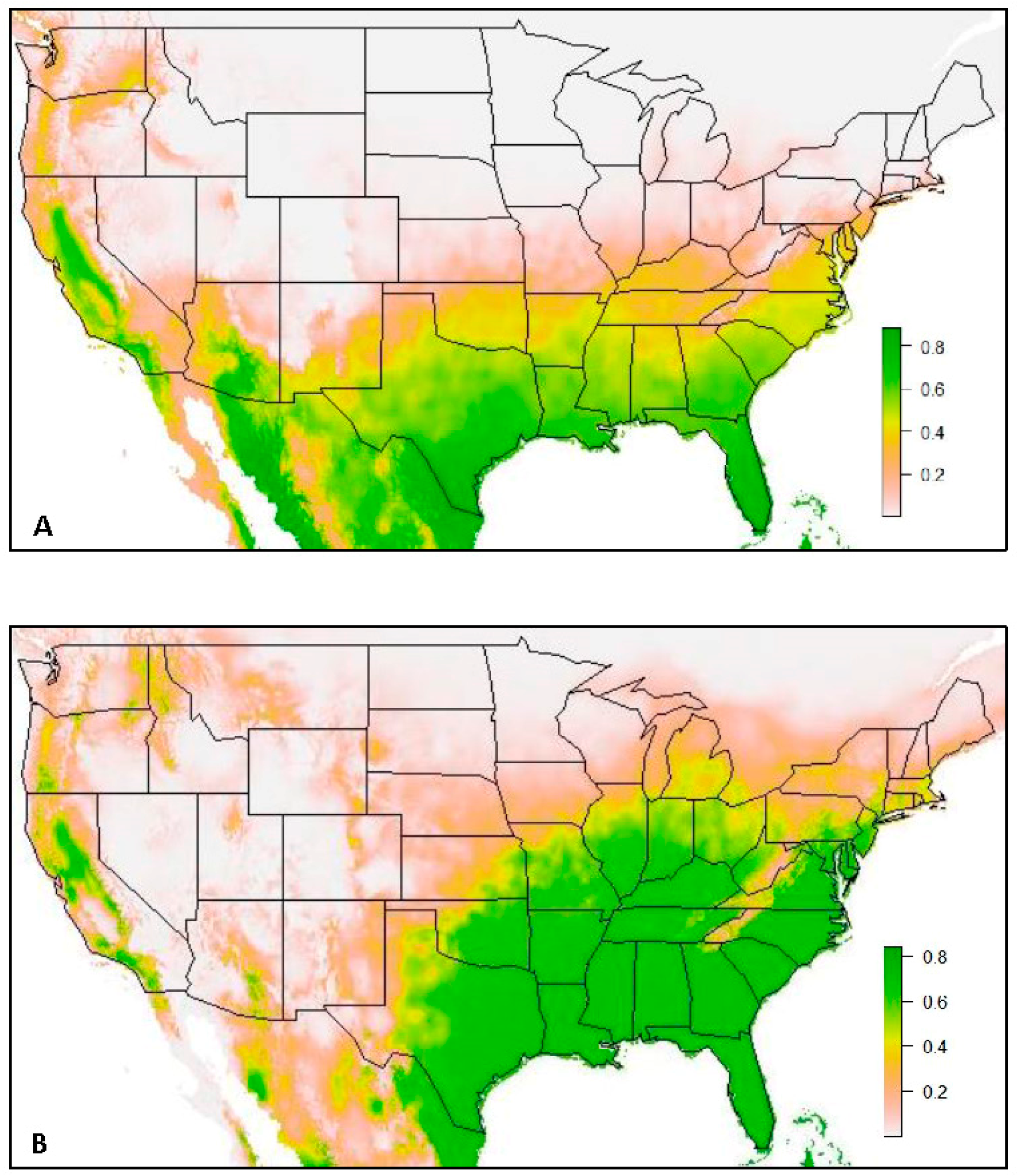
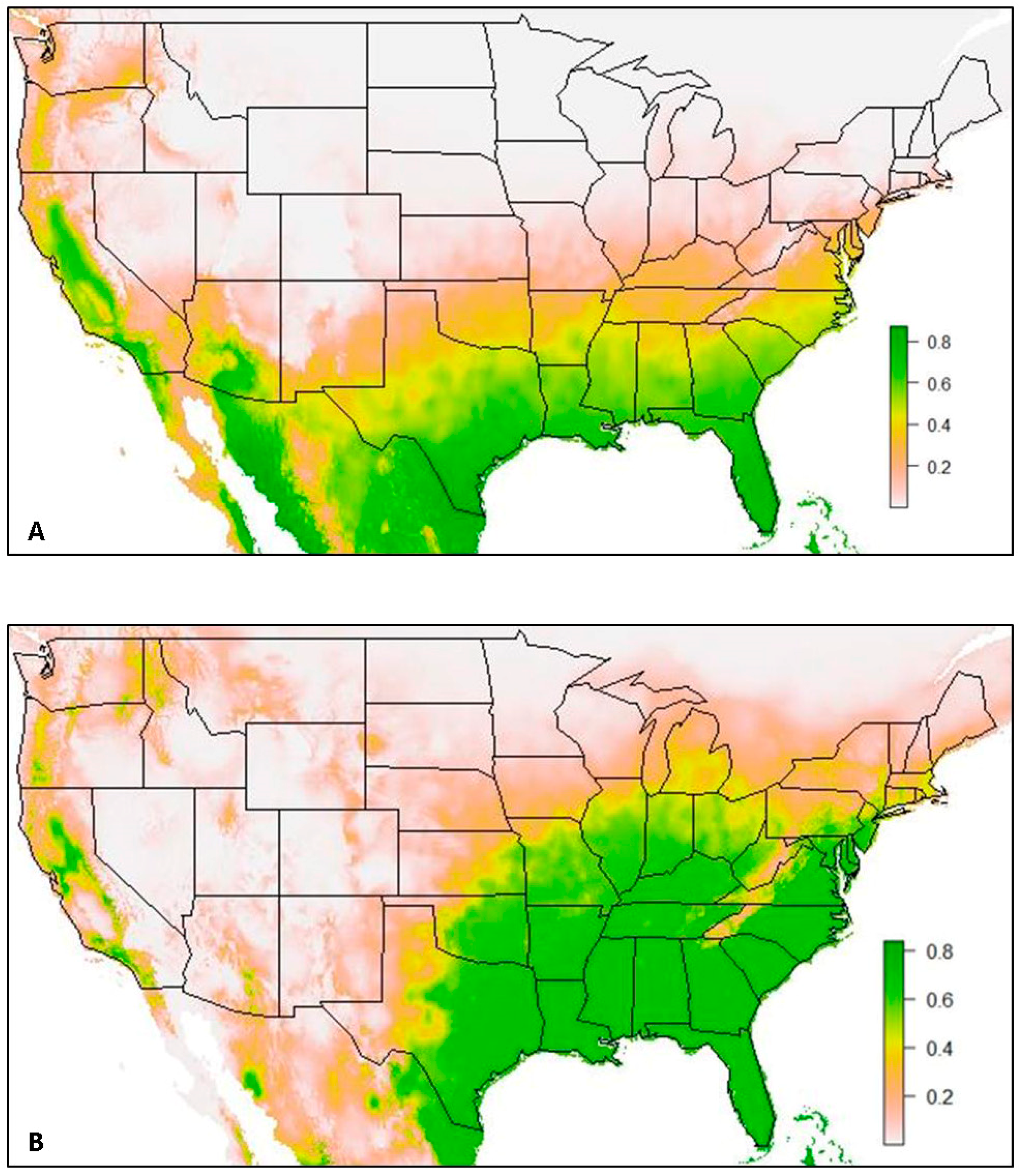
3. Results
3.1. Model 1. All USA Data
3.2. Model 2. VZL Data Excluded
4. Discussion
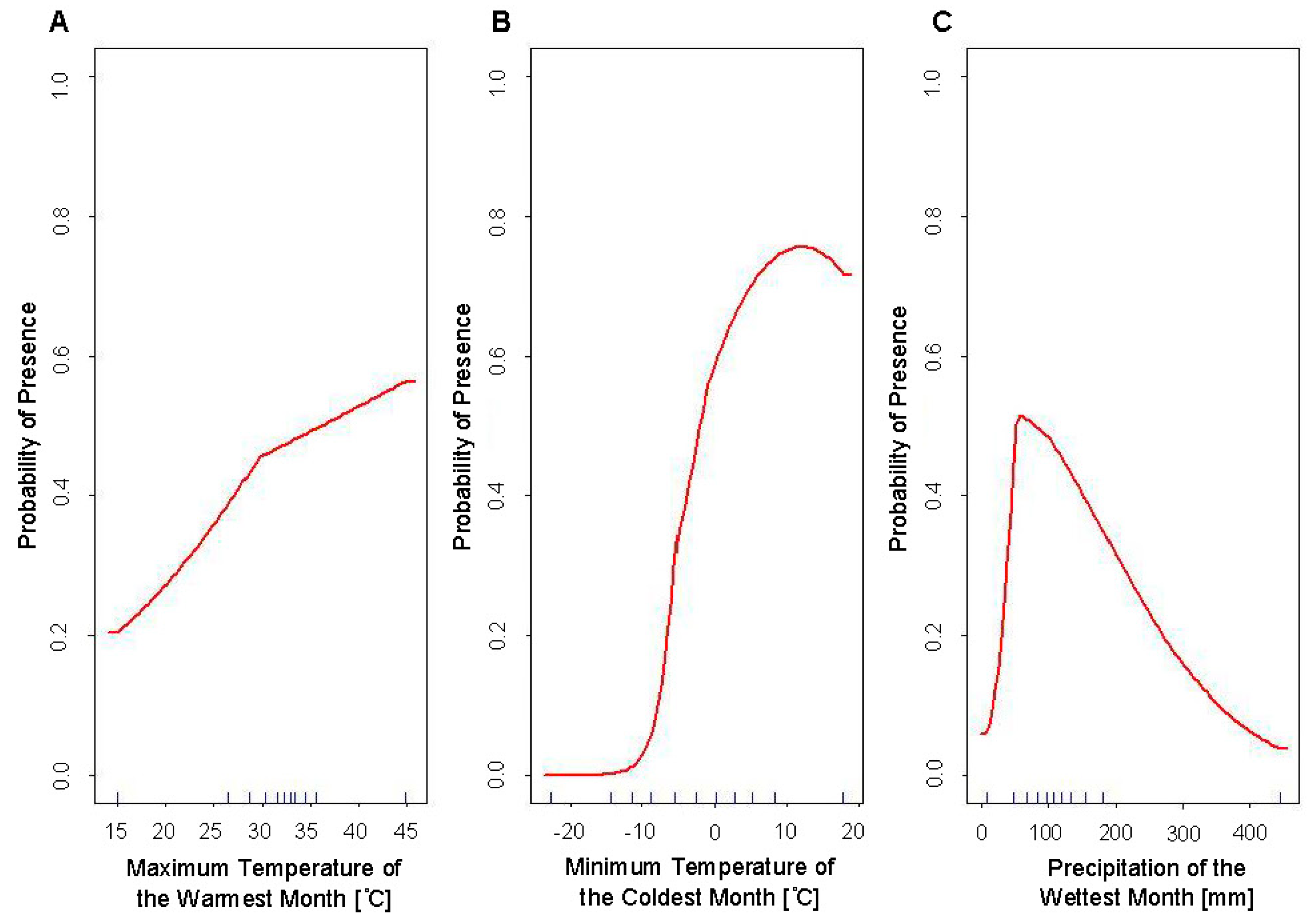
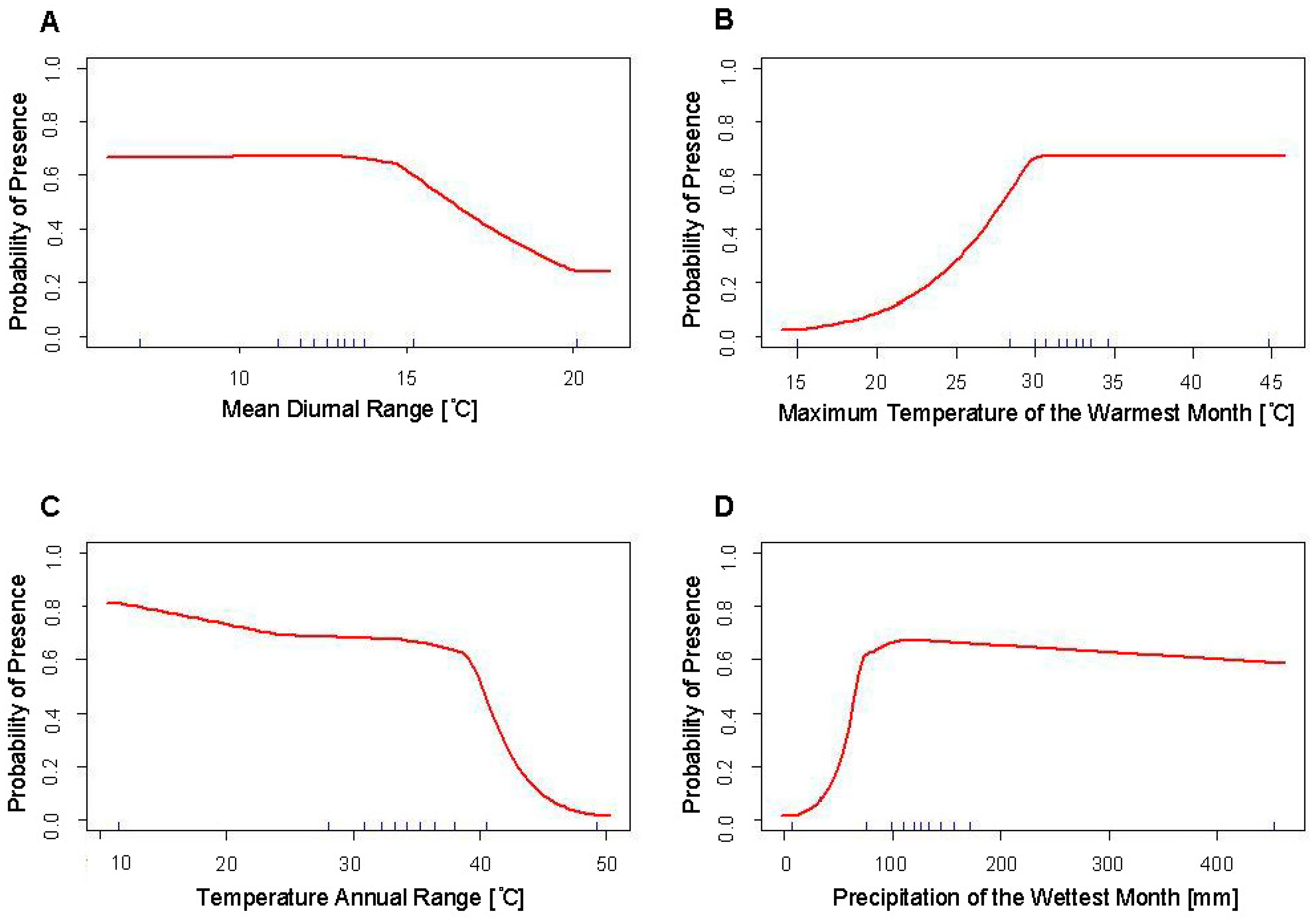
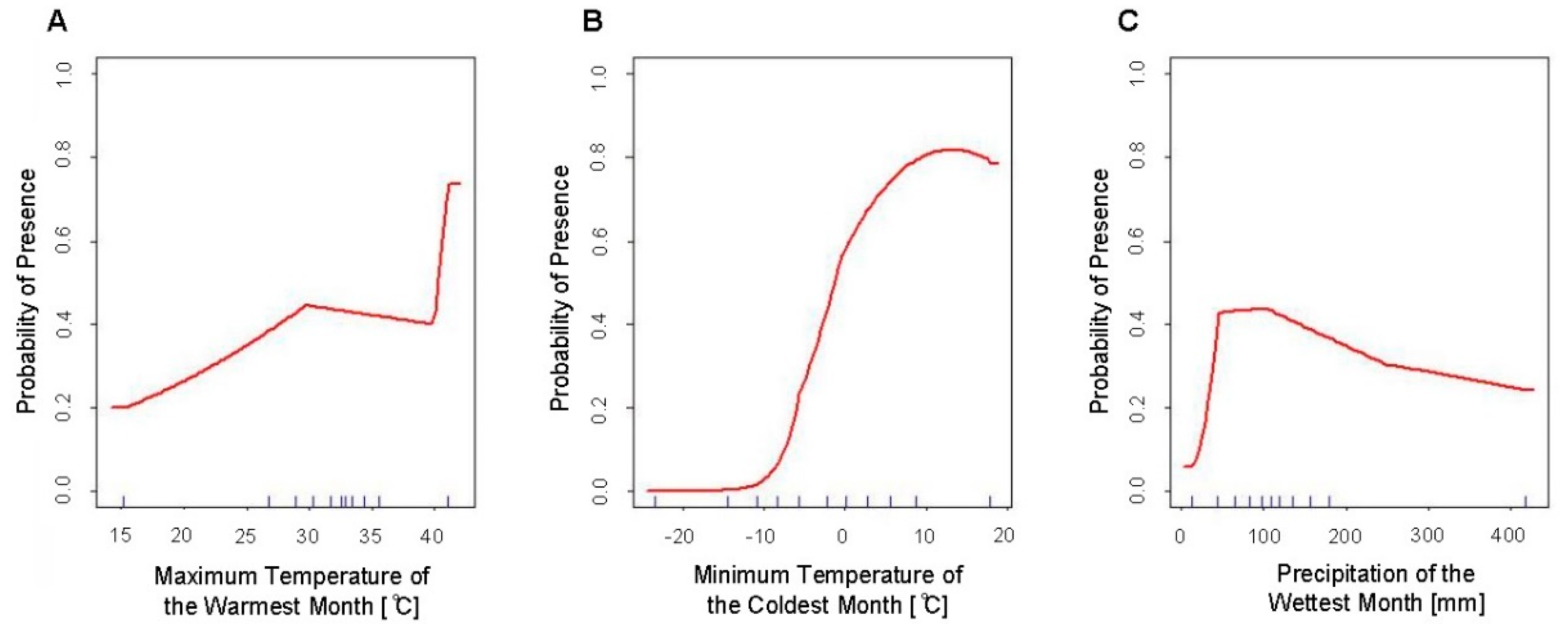
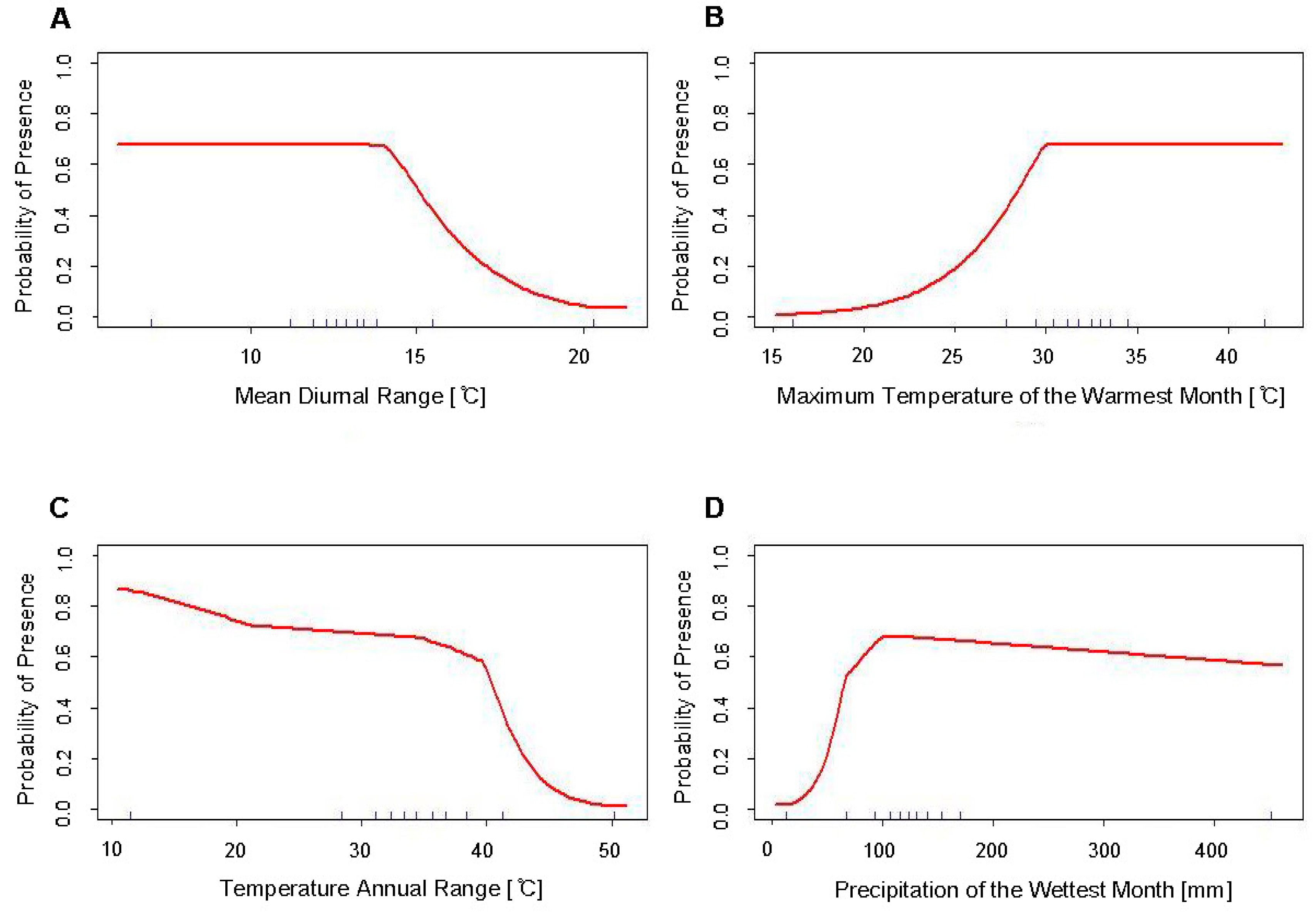
4.1. Response Curves
4.2. Evaluation and Comparison of SDMs
4.3. Data along Margins of Current Known Distributions
4.4. Potential Limitations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Bortel, W.V.; et al. The Global Distribution of the Arbovirus Vectors Aedes Aegypti and Ae. Albopictus. eLife 2015, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.B.; Eisen, R.J.; Eisen, L.; Boegler, K.A.; Moore, C.G.; McAllister, J.; Savage, H.M.; Mutebi, J.-P. Reported Distribution of Aedes (Stegomyia) Aegypti and Aedes (Stegomyia) Albopictus in the United States, 1995-2016 (Diptera: Culicidae). J. Med. Entomol. 2016, 53, 1169–1175. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.L.; Haque, U.; Monaghan, A.J.; Eisen, L.; Hahn, M.B.; Hayden, M.H.; Savage, H.M.; McAllister, J.; Mutebi, J.-P.; Eisne, R.J. Modeling the Environmental Suitability for Aedes (Stegomyia) Aegypti and Aedes (Stegomyia) Albopictus (Diptera: Culicidae) in the Contiguous United States. J. Med. Entomol. 2017, 54, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Fu, J.; Jiang, D.; Hao, M.; Lin, G. Mapping the Spatial Distribution of Aedes Aegypti and Aedes Albopictus. Acta Tropica 2018, 178, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.; Moore, C.G. Aedes (Stegomyia) Aegypti in the Continental United States: A Vector at the Cool Margin of Its Geographic Range. J. Med. Entomol. 2013, 50, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.P.; Luther, C.; Moo-Llanes, D.; Ramsey, J.M.; Danis-Lozano, R.; Peterson, A.T. Climate change influences on global distributions of dengue and chikungunya virus vectors. Philos. Trans. R. Soc. Lond. Ser. B. Biol. Sci. 2015, 370, 20140135. [Google Scholar] [CrossRef] [PubMed]
- Proestos, Y.; Christophides, G.K.; Ergüler, K.; Tanarhte, M.; Waldock, J.; Lelieveld, J. Present and Future Projections of Habitat Suitability of the Asian Tiger Mosquito, a Vector of Viral Pathogens, from Global Climate Simulation. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 20130554. [Google Scholar] [CrossRef] [PubMed]
- Peper, S.T.; Wilson-Fallon, A.; Haydett, K.; Greenberg, H.; Presley, S.M. First record of Aedes aegypti and Aedes albopictus in thirteen Panhandle region counties of Texas, U.S.A. J. Vector Ecol. 2017, 42, 352–354. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, H.S.; Wilson-Fallon, A.; Peper, S.T.; Haydett, K.M.; Presley, S.M. New records of Aedes aegypti and Aedes albopictus in eight Texas counties, U.S.A. J. Vector Ecol. 2019, 44, 199–200. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.; Shearer, F.M.; Brady, O.J.; Messina, J.P.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; et al. The Global Compendium of Aedes Aegypti and Ae. Albopictus Occurrence. Sci. Data 2015, 2, 150035. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Kraemer, M.U.G.; Brady, O.J.; Pigott, D.M.; Shearer, F.M.; Weiss, D.L.; Golding, N.; Ruktanonchai, C.W.; Gething, P.W.; Cohn, E.; et al. Mapping Global Environmental Suitability for Zika Virus. eLife 2016, 5, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Mordecai, E.A.; Cohen, J.M.; Evans, M.V.; Gudapati, P.; Johnson, L.R.; Lippi, C.A.; Miazgowicz, K.; Murdock, C.C.; Rohr, J.R.; Ryan, S.J.; et al. Detecting the Impact of Temperature on Transmission of Zika, Dengue, and Chikungunya Using Mechanistic Models. PLoS Negl. Trop. Dis. 2017, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Leta, S.; Beyene, T.J.; De Clercq, E.M.; Amenu, K.; Revie, C. Global Risk Mapping for Major Diseases Transmitted by Aedes Aegypti and Aedes Albopictus. Int. J. Infect. Dis. 2017, 67, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Cárdenas, G.; López-Mata, L.; Villaseñor, J.L.; Ortiz, E. Potential Species Distribution Modeling and the Use of Principal Component Analysis as Predictor Variables. Rev. Mex. Biodivers. 2014, 85, 189–199. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. 2016. Available online: https://www.R-project.org/ (accessed on 26 March 2018).
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Medone, P.; Ceccarelli, S.; Parham, P.E.; Figuera, A.; Rabinovich, J.E. The Impact of Climate Change on the Geographical Distribution of Two Vectors of Chagas Disease: Implications for the Force of Infection. Philos. Trans. R. Soc. B: Biol. Sci. 2015, 370, 20130560. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Elith, J. Species Distribution Modeling with R. Available online: https://cran.r-project.org/web/packages/dismo/vignettes/sdm.pdf (accessed on 19 February 2018).
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and Thermal Tolerance of Eggs and the Coexistence of Competing Mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Reiskind, M.H.; Lounibos, L.P. Spatial and Temporal Patterns of Abundance of Aedes Aegypti, L. (Stegomyia Aegypti) and Aedes Albopictus (Skuse) [Stegomyia Albopictus (Skuse)] in Southern Florida. Med. Vet. Entomol. 2013, 27, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Faull, K.J.; Williams, C.R. Intraspecific Variation in Desiccation Survival Time of Aedes Aegypti (L.) Mosquito Eggs of Australian Origin. J. Vector Ecol. 2015, 40, 292–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, A.G.; Miniota, J.; Joseph, H.A.; Brady, O.J.; Kraemer, M.U.G.; Grills, A.W.; Morrison, S.; Esposito, D.H.; Nicolucci, A.; German, M.; et al. Elevation as a proxy for mosquito-borne Zika virus transmission in the Americas. PLoS ONE 2017, 12, e0178211. [Google Scholar] [CrossRef] [PubMed]
- Aedes Aegypti and Aedes Albopictus Mosquitoes. Available online: https://www.cdph.ca.gov/Programs/CID/DCDC/CDPH%20Document%20Library/AedesDistributionMap.pdf (accessed on 3 August 2019).
- Kraemer, M.U.G.; Hay, S.I.; Pigott, D.M.; Smith, D.L.; Wint, G.R.W.; Golding, N. Progress and Challenges in Infectious Disease Cartography. Trends Parasitol. 2016, 32, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Gastón, A.; García-Viñas, J.I. Modelling Species Distributions with Penalised Logistic Regressions: A Comparison with Maximum Entropy Models. Ecol. Model. 2011, 222, 2037–2041. [Google Scholar] [CrossRef]
- Hay, S.I.; George, D.B.; Moyes, C.L.; Brownstein, J.S. Big Data Opportunities for Global Infectious Disease Surveillance. PLoS Med. 2013, 10, 2–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Percent Contribution | Permutation Importance |
|---|---|---|
| Min. Temperature of Coldest Month (Bio6) | 92.1 | 88.4 |
| Precipitation of Wettest Month (Bio13) | 5.1 | 9.3 |
| Max. Temperature of Warmest Month (Bio5) | 2.7 | 2.3 |
| Variable | Percent Contribution | Permutation Importance |
|---|---|---|
| Temperature Annual Range (Bio7) | 39.4 | 41.7 |
| Max. Temperature of Warmest Month (Bio5) | 16 | 27.2 |
| Precipitation of Wettest Month (Bio13) | 40.7 | 26.6 |
| Mean Diurnal Range (Bio2) | 3.8 | 4.6 |
| Variable | Percent Contribution | Permutation Importance |
|---|---|---|
| Min. Temperature of Coldest Month (Bio6) | 92.6 | 92.0 |
| Precipitation of Wettest Month (Bio13) | 2.7 | 4.5 |
| Max. Temperature of Warmest Month (Bio5) | 4.7 | 3.4 |
| Variable | Percent Contribution | Permutation Importance |
|---|---|---|
| Temperature Annual Range (Bio7) | 31.8 | 33.9 |
| Max. Temperature of Warmest Month (Bio5) | 16.7 | 29.4 |
| Precipitation of Wettest Month (Bio13) | 43.1 | 19.6 |
| Mean Diurnal Range (Bio2) | 8.5 | 17.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tiffin, H.S.; Peper, S.T.; Wilson-Fallon, A.N.; Haydett, K.M.; Cao, G.; Presley, S.M. The Influence of New Surveillance Data on Predictive Species Distribution Modeling of Aedes aegypti and Aedes albopictus in the United States. Insects 2019, 10, 400. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10110400
Tiffin HS, Peper ST, Wilson-Fallon AN, Haydett KM, Cao G, Presley SM. The Influence of New Surveillance Data on Predictive Species Distribution Modeling of Aedes aegypti and Aedes albopictus in the United States. Insects. 2019; 10(11):400. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10110400
Chicago/Turabian StyleTiffin, Hannah S., Steven T. Peper, Alexander N. Wilson-Fallon, Katelyn M. Haydett, Guofeng Cao, and Steven M. Presley. 2019. "The Influence of New Surveillance Data on Predictive Species Distribution Modeling of Aedes aegypti and Aedes albopictus in the United States" Insects 10, no. 11: 400. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10110400