What Can Integrated Analysis of Morphological and Genetic Data Still Reveal about the Anastrepha fraterculus (Diptera: Tephritidae) Cryptic Species Complex?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Analysis of ITS1 Sequences
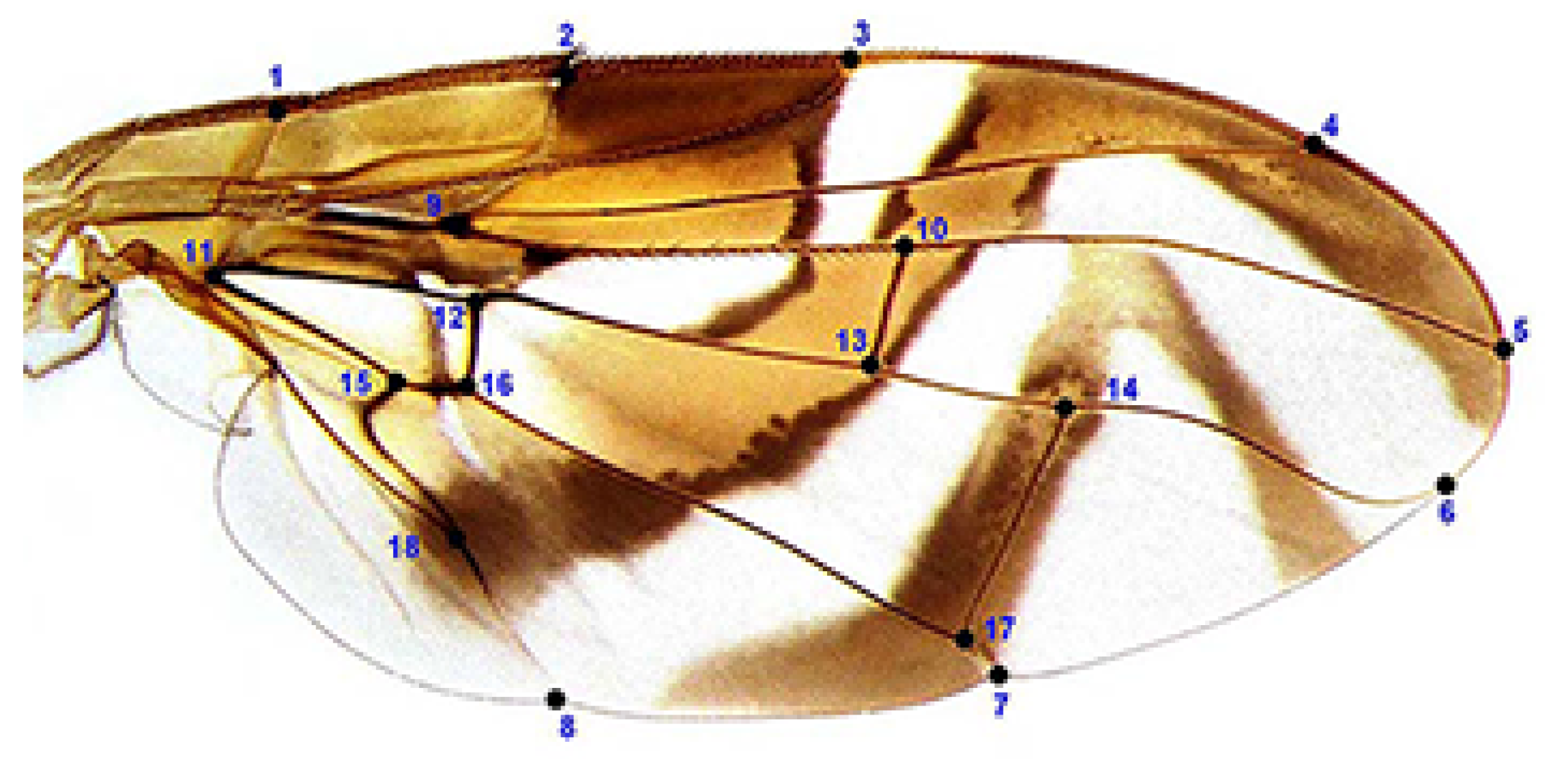
2.3. Morphometric Assessment
3. Results
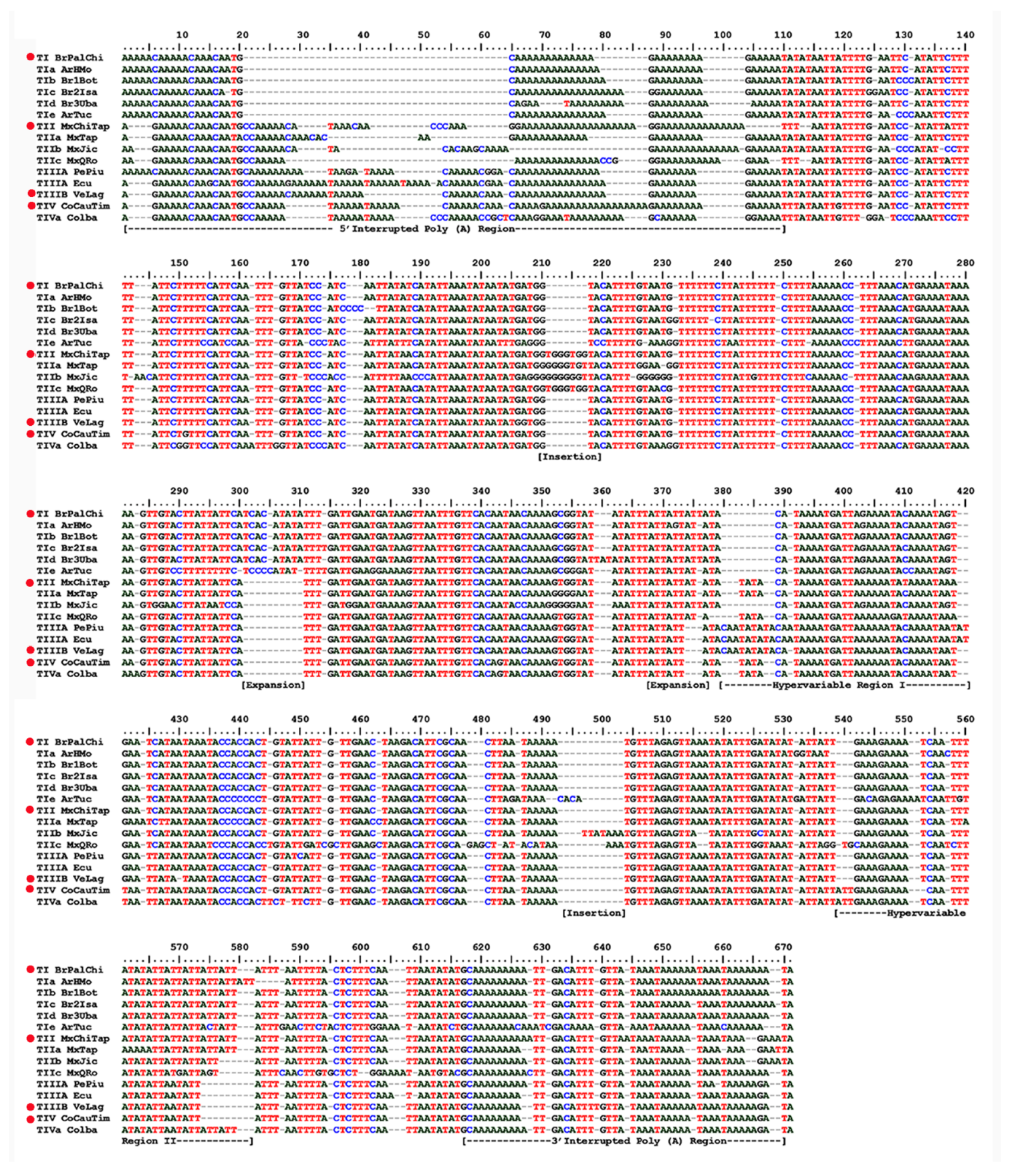
3.1. Analyses of ITS1 Sequences
3.2. Characterization of the ITS1 Sequence Types
3.2.1. Sequence Type TI
3.2.2. Sequence Type TII
3.2.3. Sequence Types TIII A, B, C
3.2.4. Sequence Type TIV
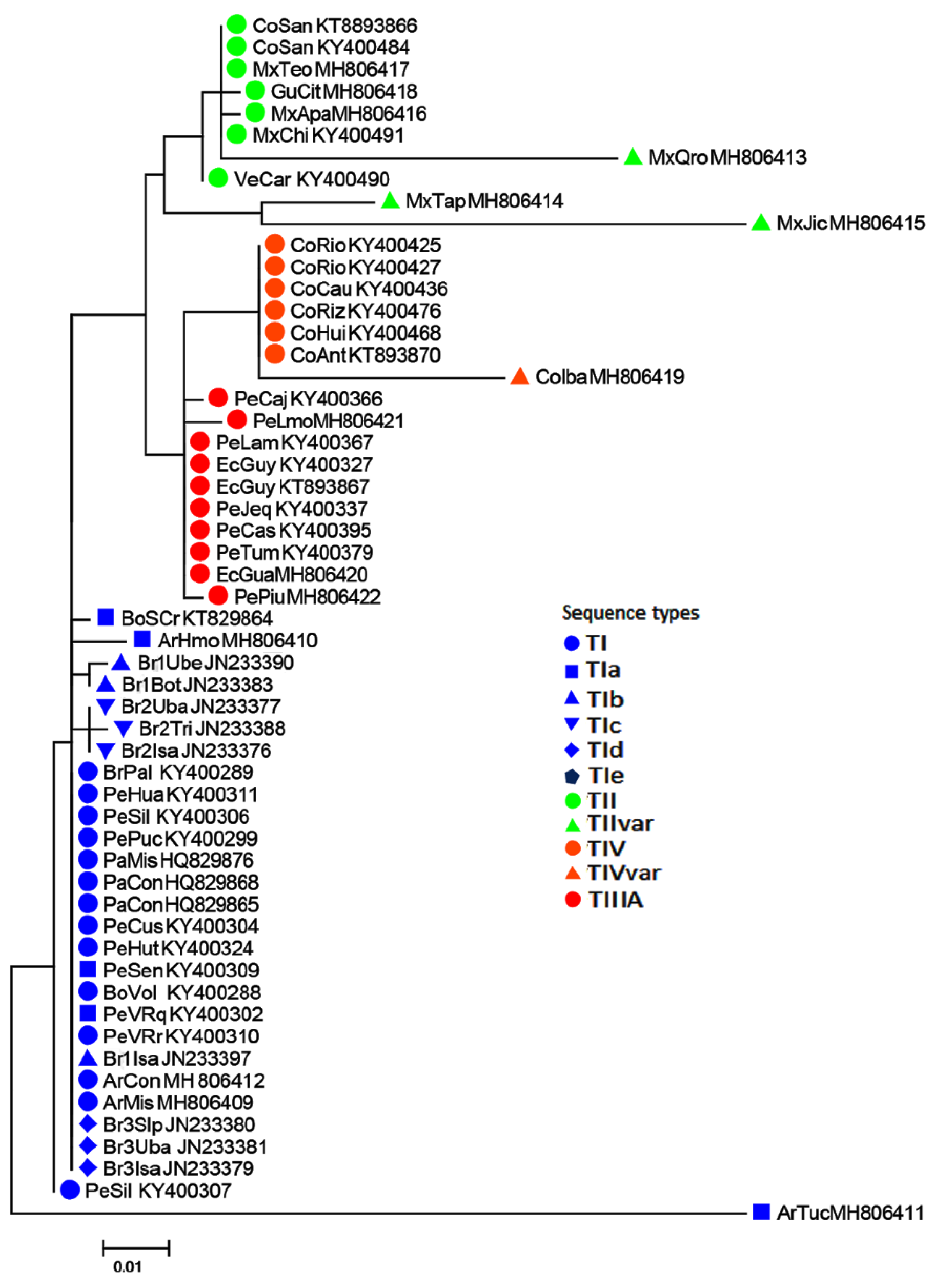
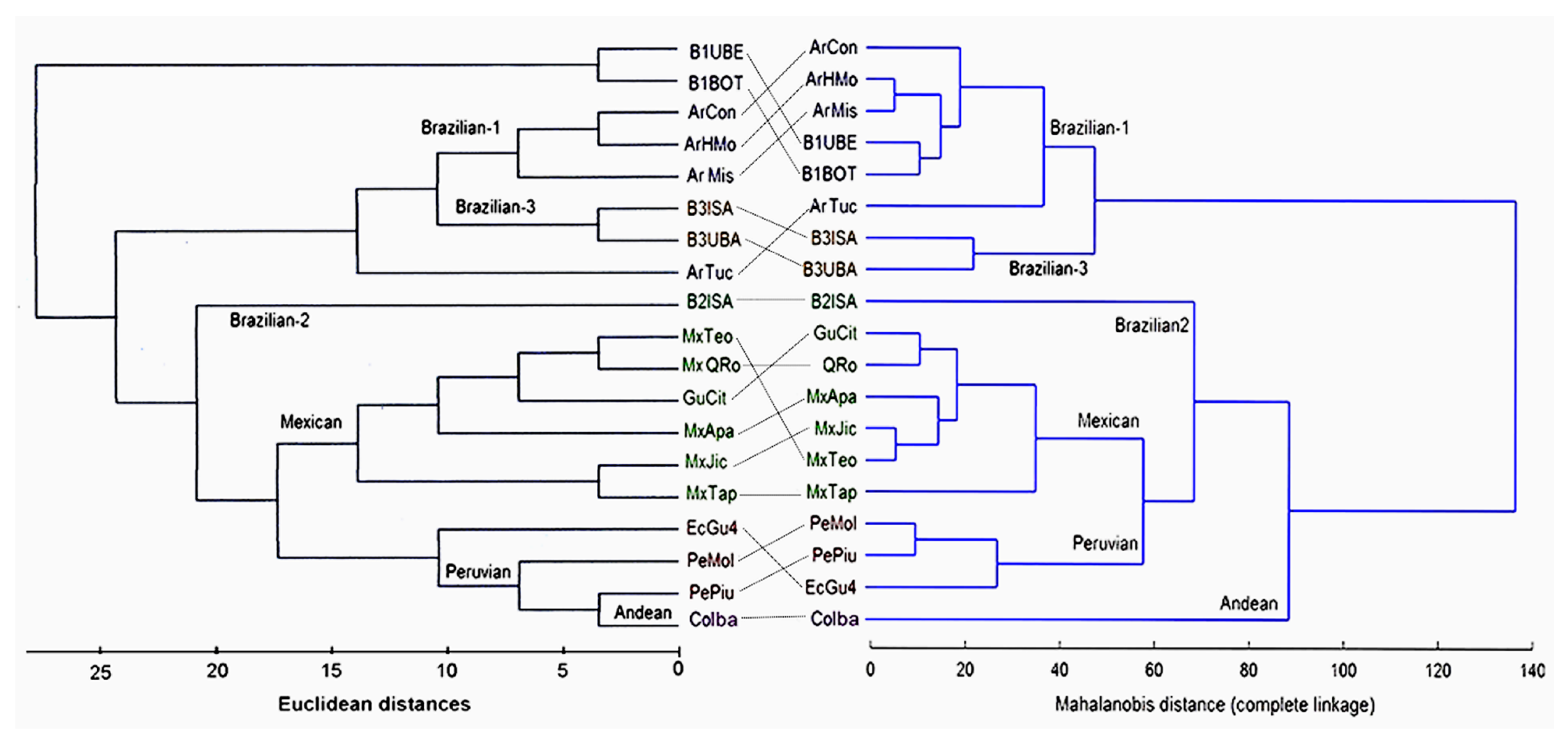
3.3. Relationships of Sequence Types among Samples of the AF Complex
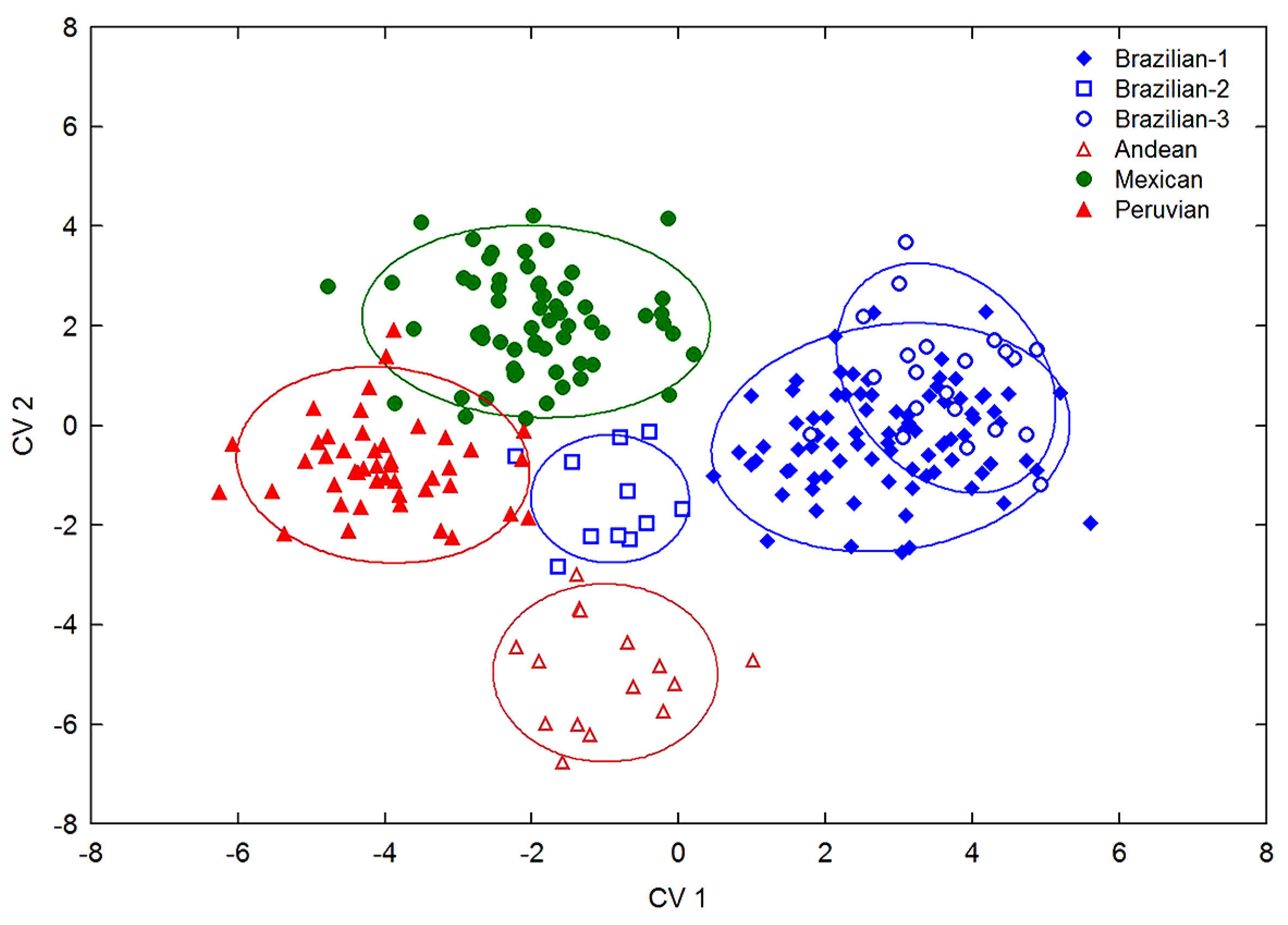
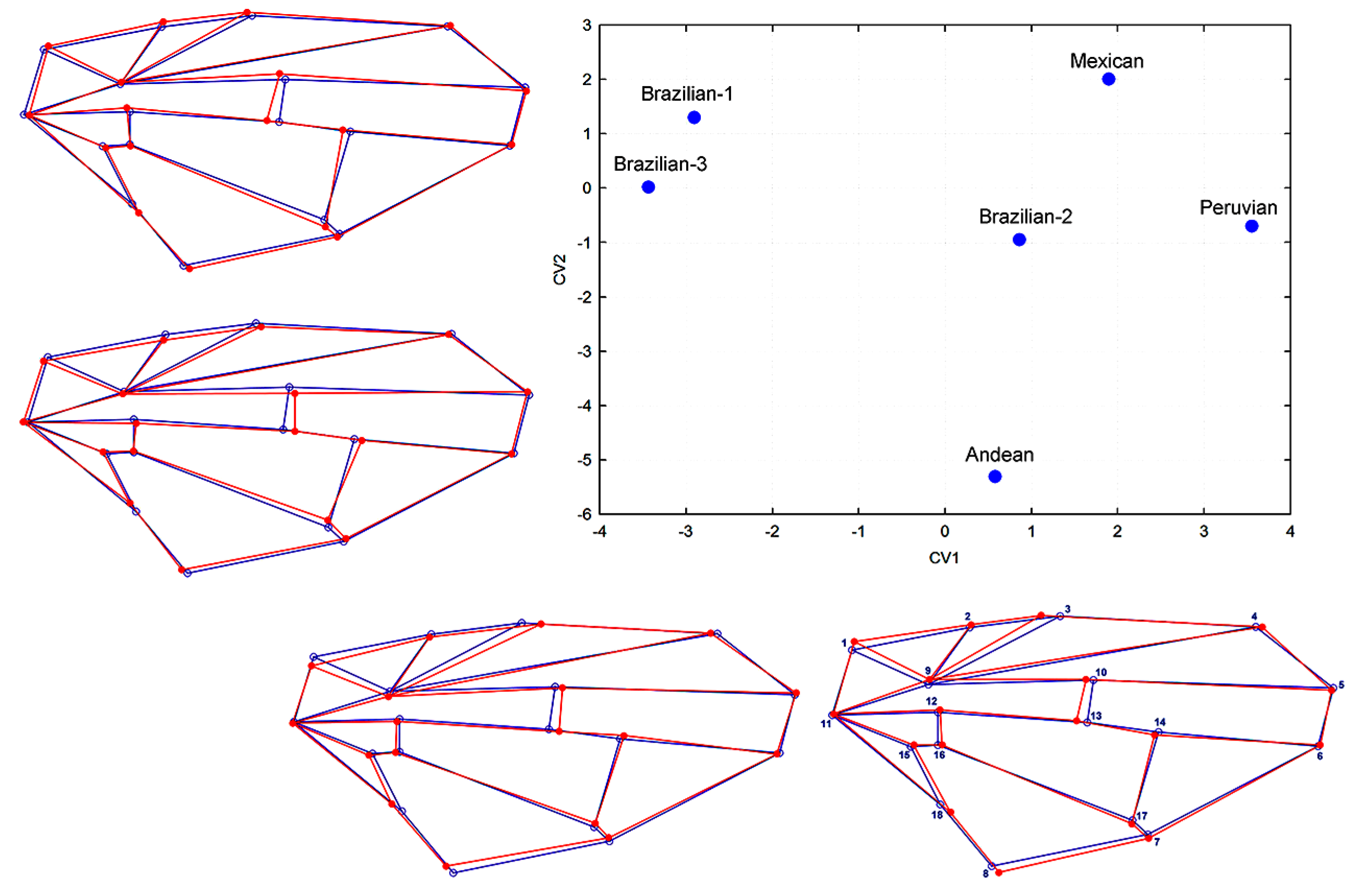
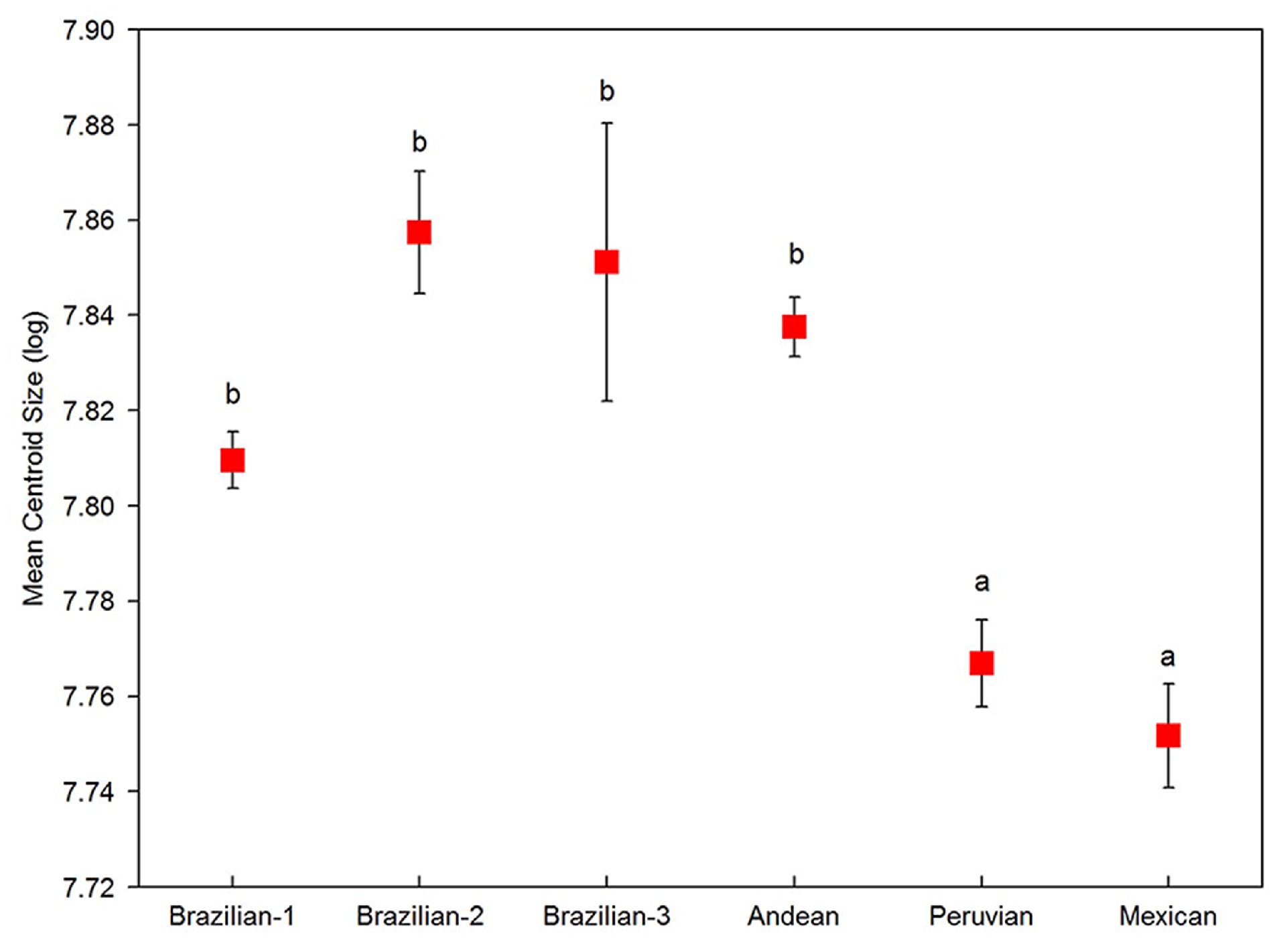
3.4. Morphological Relationships
3.5. Wings and ITS1 Data Relationships
4. Discussion
4.1. ITS1 as a Molecular Marker
4.2. ITS1 and Morphotype Relationships
4.3. Scenarios of Divergences Within the AF Complex
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, UK, 1992. [Google Scholar]
- Norrbom, A.L.; Carroll, L.E.; Thompson, C.F.; White, I.M.; Freidberg, A. Systematic database of names. In Fruit Fly Expert Identification System and Systematic Information Database, Myia 9; Thompson, F.C., Ed.; Backhuys Publishers: Slikweg, The Netherlands, 1998; pp. 65–251. [Google Scholar]
- Norrbom, A.L.; Zucchi, R.A.; Hernández-Ortiz, V. Phylogeny of the genera Anastrepha and Toxotrypana (Trypetinae: Toxotrypanini) based on morphology. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Ratón, FL, USA, 1999; pp. 299–342. [Google Scholar]
- Norrbom, A.L.; Korytkowiski, C. New species of and taxonomic notes on Anastrepha (Diptera, Tephritidae). Zootaxa 2011, 2740, 1–23. [Google Scholar] [CrossRef]
- Norrbom, A.L.; Korytkowski, C.A.; Zucchi, R.A.; Uramoto, K.; Venable, G.L.; McCormick, J.; Dallwitz, M.J. Anastrepha and Toxotrypana: Descriptions, Illustrations, and Interactive Keys. Version: 28th. 2013. Available online: http://delta-intkey.com (accessed on 9 April 2017).
- Stone, A. The Fruit Flies of the Genus Anastrepha; US Department of Agriculture, Miscellaneous Publication: Washington, DC, USA, 1942; Volume 439, pp. 1–112. [Google Scholar]
- Baker, A.C.; Stone, W.E.; Plummer, C.C.; MacPhail, H. A Review of Studies on the Mexican Fruit Fly and Related Mexican Species; US Department of Agriculture, Miscellaneous Publication: Washington, DC, USA, 1944; Volume 531, pp. 1–155. [Google Scholar]
- Mendes, L.O.T. Observações citológicas em “moscas-das-frutas”. Bragantia 1958, 17, 29–39. [Google Scholar] [CrossRef]
- Bush, G.L. The cytotaxonomy of the larvae of some Mexican fruit flies in the genus Anastrepha. (Tephritidae: Diptera). Psyche 1962, 6, 87–101. [Google Scholar] [CrossRef]
- Solferini, V.N.; Morgante, J.S. Karyotype study of eight species of Anastrepha (Diptera: Tephritidae). Caryologia 1987, 40, 229–241. [Google Scholar] [CrossRef]
- Morgante, S.; Malavasi, A.; Bush, G.L. Biochemical systematics and evolutionary relationships of Neotropical Anastrepha. Ann. Entomol. Soc. Am. 1980, 73, 622–630. [Google Scholar] [CrossRef]
- Steck, G.J. Biochemical systematics and population genetic structure of Anastrepha fraterculus and related species (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 1991, 84, 10–28. [Google Scholar] [CrossRef]
- Steck, G.J.; Sheppard, W.S. Mitochondrial DNA variation in Anastrepha fraterculus. In. Fruit Flies: Biology and Management; Aluja, M., Liedo, P., Eds.; Springer: New York, NY, USA, 1993; pp. 9–14. [Google Scholar]
- Santos, P. Variabilidade do DNA Mitocondrial em Populações de Três Espécies de Anastrepha (Diptera, Tephritidae). Master’s Thesis, Universidade de São Paulo, São Paulo, Brazil, 1994. [Google Scholar]
- McPheron, B.A.; Han, H.Y.; Silva, J.G.; Norrbom, A.L. Phylogeny of the genus Anastrepha (Trypetinae: Toxotrypanini) based upon 16S rDNA mitochrondial sequences. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Ratón, FL, USA, 1999; pp. 343–361. [Google Scholar]
- Smith-Caldas, M.R.B.; McPheron, B.A.; Silva, J.G.; Zucchi, R.A. Phylogenetic relationships among species of the fraterculus group (Anastrepha: Diptera: Tephritidae) inferred from DNA sequences of mitochondrial cytochrome oxidade 1. Neotrop. Entomol. 2001, 30, 565–573. [Google Scholar] [CrossRef]
- Barr, N.B.; Cui, L.; McPheron, B.A. Molecular systematics of nuclear gene period in genus Anastrepha (Tephritidae). Ann. Entomol. Soc. Am. 2005, 98, 173–180. [Google Scholar] [CrossRef]
- Selivon, D. Estudo Sobre a Diferenciação Populacional em Anastrepha fraterculus (Wiedemann) (Diptera: Tephritidae). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 1996. [Google Scholar]
- Selivon, D.; Vretos, C.; Fontes, L.; Perondini, A.L.P. New variant forms in the Anastrepha fraterculus complex. In Proceedings of the 6th International Symposium on Fruit Flies of Economic Importance, Stellenbosh, South Africa, 6–10 May 2002; pp. 253–258. [Google Scholar]
- Selivon, D.; Perondini, A.L.P.; Morgante, J.S. A genetic-morphological characterization of two cryptic species of the Anastrepha fraterculus complex (Diptera, Tephritidae). Ann. Entomol. Soc. Am. 2005, 98, 367–381. [Google Scholar] [CrossRef]
- Goday, C.; Selivon, D.; Perondini, A.L.P.; Greciano, P.G.; Ruiz, M.F. Cytological characterization of sex chromosomes and ribosomal DNA location in Anastrepha species (Diptera, Tephtiridae). Cytogent. Genome Res. 2006, 114, 70–76. [Google Scholar] [CrossRef]
- Selivon, D.; Perondini, A.L.P. Eggshell morphology in two cryptic species of Anastrepha fraterculus (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 1998, 91, 473–478. [Google Scholar] [CrossRef]
- Rocha, L.S.; Selivon, D. Studies on highly repetitive DNA in cryptic species of the Anastrepha fraterculus complex (Diptera: Tephritidae). In Proceedings of the 6th International Symposium on Fruit Flies of Economic Importance, Stelllenbosch, South Africa, 6–10 May 2002; pp. 415–418. [Google Scholar]
- Selivon, D.; Perondini, A.L.P. Extrusion of yolk masses by hybrid embryos of two cryptic species of Anastrepha fraterculus (Diptera, Tephritidae). Brazil. J. Genet. 1997, 20, 253–255. [Google Scholar]
- Selivon, D.; Morgante, J.S.; Perondini, A.L.P. Egg size, yolk mass extrusion and hatching behavior in two cryptic species of Anastrepha fraterculus (Wied.) (Diptera, Tephritidae). Brazil. J. Genet. 1997, 20, 587–594. [Google Scholar] [CrossRef]
- Selivon, D.; Perondini, A.L.P.; Morgante, J.S. Haldane’s rule and other aspects of reproductive isolation observed in the Anastrepha fraterculus complex (Diptera, Tephhritidae). Genet. Mol. Biol. 1999, 22, 507–510. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V.; Gomez-Anaya, J.A.; Sanchez, A.; McPheron, B.A.; Aluja, M. Morphometric analysis of Mexican and South American populations of the Anastrepha fraterculus complex (Diptera:Tephritidae) and recognition of a distinct Mexican morphotype. Bull. Entomol. Res. 2004, 94, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ortiz, V.; Bartolucci, A.; Morales-Valles, P.; Frias, D.; Selivon, D. Cryptic species of the Anastrepha fraterculus complex (Diptera, Tephritidae): A multivariate approach for the recognition of South American morphotypes. Ann. Entomol. Soc. Am. 2012, 105, 305–318. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V.; Canal, N.A.; Tigrero Salas, J.O.; Ruíz-Hurtado, F.M.; Dzul-Cauich, J.F. Taxonomy and phenotypic relationships of the Anastrepha fraterculus complex in the Mesoamerican and Pacific Neotropical dominions (Diptera, Tephritidae). ZooKeys 2015, 540, 95–124. [Google Scholar] [CrossRef] [PubMed]
- Hendrichs, J.; Vera, M.T.; De Meyer, M.; Clarke, A.R. Resolving cryptic species complexes of major tephritid pests. ZooKeys 2015, 540, 5–39. [Google Scholar] [CrossRef]
- Prezotto, L.F. Análise do ITS1 do DNA ribossômico em espécies do complexo Anastrepha fraterculus (Diptera, Tephritidae). Master’s Thesis, Universidade de São Paulo, São Paulo, Brazil, 2008. [Google Scholar]
- Sutton, B.D.; Steck, G.J.; Norrbom, A.L.; Rodriguez, E.J.; Srivastava, P.; Alvarado, N.N.; Colque, F.; Landa, E.Y.; Sánchez, J.J.; Quisberth, E.; et al. Nuclear ribosomal internal transcribed spacer 1 (ITS1) variation in the Anastrepha fraterculus cryptic species complex (Diptera, Tephritidae) of the Andean region. ZooKeys 2015, 540, 175–191. [Google Scholar] [CrossRef]
- Manni, M.; Lima, K.M.; Guglielmino, C.R.; Lanzavecchia, S.B.; Juri, M.; Vera, T.; Cladera, J.; Scolari, F.; Gomulski, L.; Bonizzoni, M.; et al. Relevant genetic differentiation among Brazilian populations of Anastrepha fraterculus (Diptera, Tephritidae). ZooKeys 2015, 540, 157–173. [Google Scholar]
- Sánchez, L.; Perondini, A.L.P.; Selivon, D. Understanding the dynamics underpinning the geographical distribution of the Anastrepha fraterculus complex in Eastern Brazil. Trends Entomol. 2018, 14, 17–31. [Google Scholar] [CrossRef]
- Morrone, J.J. Biogeographical areas and transition zones of Latin America and the Caribbean Island based on panbiogeographical and cladistics analyses of the entomofauna. Ann. Rev. Entomol. 2006, 51, 467–494. [Google Scholar] [CrossRef] [PubMed]
- Jowett, T.; Rizki, T.M.; Rizki, R.M. Regulation of synthesis of larval serum proteins after transplantation of larval fat body into adult Drosophila melanogaster. Dev. Biol. 1986, 116, 23–30. [Google Scholar] [CrossRef]
- Togawa, R.C.; Brigido, M.M. PHPH: Web based Tool for Simple Electropherogram Quality Analysis. Available online: http://asparagin.cenargen.embrapa.br/ phph/references.html (assessed on 15 March 2017).
- Lopes, G.N.; Arias, O.R.; Cônsoli, F.L.; Zucchi, R.A. The identity of specimens of the Anastrepha fraterculus complex (Diptera, Tephritidae) with atypical aculeus tip. Neotrop. Entomol. 2013, 42, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, S.; Kumar, S. MEGA 6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Sneath, P.H.A. Numerical Taxonomy: The Principles and Practice of Numerical Classification; Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
- Neil, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using th neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 1103–11035. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Marcus, L.F. A revolution in morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Rohlf, F.J. TpsDig Version 2.16; State University of New York: Stony Brook, NY, USA, 2010; Available online: http://life.bio.sunysb.edu/morph/ (accessed on April 2014).
- Rohlf, F.J. Tps Utility Program Version 1.52; State University of New York: Stone Brook, NY, USA, 2012; Available online: http://life.bio.sunysb.edu/morph/ (accessed on April 2014).
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks. Package (PMCMR)R, 2014. Available online: http://CRAN.R-project.org/package=PMCMR (accessed on April 2014).
- Hillis, D.H.; Dixon, M.T. Ribosomal DNA: Molecular evolution and phylogenetic inference. Q. Rev. Biol. 1991, 66, 411–453. [Google Scholar] [CrossRef]
- Elder, J.F.; Turner, B.J. Concerted evolution of repetitive DNA sequences in Eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Douglas, L.J.; Haymer, D.S. Ribosomal ITS1 polymorphisms in Ceratitis capitata and Ceratitis rosa (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2001, 94, 726–731. [Google Scholar] [CrossRef]
- Basso, A.; Sonvico, A.; Quesada-Allue, L.A.; Manso, F. Karyotypic and molecular identification of laboratory stocks of the South American fruit fly Anastrepha fraterculus (Wied) (Diptera: Tephritidae). J. Econ. Entomol. 2003, 96, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Haymer, D.S. Random amplified polymorphic DNAs and microsatellites: What are they, and can they tell us anything we don’t know? Ann. Entomol. Soc. Am. 1994, 87, 717–722. [Google Scholar] [CrossRef]
- Schlötterer, C.; Hauser, M.T.; von Haeseler, A.; Tautz, D. Comparative evolutionary analysis of rDNA ITS region in Drosophila. Mol. Biol. Evol. 1994, 11, 513–522. [Google Scholar]
- Malavasi, A.; Morgante, J.S. Population genetics of Anastrepha fraterculus (Diptera, Teprhitidae) in different hosts: Genetic differentiation and heterozygosity. Genetica 1983, 60, 207–211. [Google Scholar] [CrossRef]
- Morgante, J.S.; Malavasi, A. Genetic variability in populations of South American fruit fly Anastrepha fraterculus (Tephritidae). Rev. Bras. Genet. 1985, 8, 241–247. [Google Scholar]
- Alberti, A.C.; Calcagno, G.; Saidman, B.O.; Vilardi, J.C. Analysis of the genetic structure of a natural population of Anastrepha fraterculus (Diptera, Tephritidae). Ann. Entomol. Soc. Am. 1999, 92, 731–736. [Google Scholar] [CrossRef]
- Alberti, A.C.; Confalonieri, V.A.; Zandomeni, R.O.; Vilardi, J.C. Phylogeographic studies on natural populations of the South American fruit fly, Anastrepha fraterculus (Diptera: Tephritidae). Genetica 2008, 132, 1–8. [Google Scholar] [CrossRef]
- Basso, A.; Manso, F. Are Anastrepha fraterculus chromosomal polymorphism an isolation barrier? Cytobios 1998, 93, 103–111. [Google Scholar]
- Alberti, A.C.; Rodriguero, M.S.; Cendra, P.G.; Saidman, B.O.; Vilardi, J.C. Evidence indicating that Argentine populations of Anastrepha fraterculus (Díptera: Tephritidae) belong to a single biological species. Ann. Entomol. Soc. Am. 2002, 95, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Petit-Marty, N.; Vera, M.T.; Calcagno, G.; Cladera, J.L.; Segura, D.F.; Allinghi, A.; Rodriguero, M.; Cendra, P.G.; Viscarret, M.M.; Vilardi, J.C. Sexual behavior and mating compatibility among four populations of Anastrepha fraterculus (Diptera: Tephritidae) from Argentina. Ann. Entomol. Soc. Am. 2004, 97, 1320–1327. [Google Scholar] [CrossRef] [Green Version]
- Giardini, M.R.; Milla, F.H.; Lanzavechia, M.N.; Cladera, J.L. Sex chromosomes in mitotic and polytene tissue of Anastrepha fraterculus (Diptera: Tephritidae) from Argentina: A review. ZooKeys 2015, 540, 83–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schutze, M.K.; Virgilio, M.; Norrbom, A.; Clarke, A.R. Tephritid integrative taxonomy: Where we are now, with a focus on the resolution of three tropical fruit fly species complexes. Ann. Rev. Entomol. 2017, 62, 147–164. [Google Scholar] [CrossRef]
- Rull, J.; Abraham, S.; Kovaleski, A.; Segura, D.F.; Mendonza, M.; Liendo, M.C.; Vera, M.T. Evolution of prezygotic and postzygotic barriers to gene flow among three cryptic species within the Anastrepha fraterculus complex. Entomol. Exp. Appl. 2013, 148, 213–222. [Google Scholar] [CrossRef]
- Devescovi, F.; Abraham, S.; Roriz, A.K.; Nolazco, N.; Castañeda, R.; Tadeo, E.; Caceres, C.; Segura, D.F.; Vera, M.T.; Joachim-Bravo, I.; et al. Ongoing speciation within the Anastrepha fraterculus cryptic species complex: The case of the Andean morphotype. Entomol. Exp. Appl. 2014, 152, 238–247. [Google Scholar] [CrossRef]
- Abraham, S.; Rull, J.; Mendoza, M.; Liendo, M.C.; Devescovi, F.; Roriz, A.K.; Kovaleski, A.; Segura, D.F.; Vera, M.T. Differences in sperm storage and remating propensity between adult females of two morphotypes of the Anastrepha fraterculus (Diptera: Tephritidae) cryptic species complex. Bull. Entomol. Res. 2014, 104, 376–382. [Google Scholar] [CrossRef]
- Roriz, A.K.P.; Japyassú, H.F.; Joachin-Bravo, I.S. Incipient speciation in the Anastrepha fraterculus cryptic species complex: Reproductive compatibility between A. sp.1 aff. fraterculus and A. sp.3 aff. fraterculus. Entomol. Exp. Appl. 2017, 162, 346–357. [Google Scholar]
- Diaz, F.; Lima, A.L.A.; Nakamura, A.L.; Fernandes, F.; Sobrinho, I., Jr.; Brito, R.A. Evidence for introgression among three species of the Anastrepha fraterculus group, a radiating species complex of fruit flies. Front. Genet. 2018, 9, 359. [Google Scholar] [CrossRef]
- Rull, J.; Abraham, S.; Kovaleski, A.; Segura, D.F.; Islam, A.; Wornoayporn, V.; Dammalage, T.; Santo Tomas, U.; Vera, M.T. Random mating and reproductive compatibility among Argentinean and southern Brazilian populations of Anastrepha fraterculus (Diptera: Tephritidae). Bull. Entomol. Res. 2012, 102, 435–443. [Google Scholar] [CrossRef]
- Kovaleski, A.; Uramoto, K.; Sugayama, R.L.; Canal, N.A.; Malavasi, A. A survey of Anastrepha Schiner (Diptera, Tephritidae) species in the apple growing area of the state of Rio Grande do Sul, Brazil. Rev. Bras. Entomol. 1999, 43, 229–234. [Google Scholar]
- Vaníčková, L.; Hernández-Ortiz, V.; Bravo, I.S.; Dias, V.; Roriz, A.K.; Laumann, R.A.; de Lima Mendonça, A.; Paranhos, B.A.; do Nascimento, R.R. Current knowledge of the species complex Anastrepha fraterculus (Diptera, Tephritidae) in Brazil. Zookeys 2015, 540, 211–237. [Google Scholar] [CrossRef] [PubMed]
- Dias, V.S.; Silva, J.G.; Lima, K.M.; Petitinga, C.S.; Hernández-Ortiz, V.; Laumann, R.A.; Paranhos, B.J.; Uramoto, K.; Zucchi, R.A.; Joachim-Bravo, I.S. An integrative multidisciplinary approach to understanding cryptic divergence in Brazilian species of the Anastrepha fraterculus complex (Diptera: Tephritidae). Biol. J. Linn. Soc. 2016, 117, 725–746. [Google Scholar] [CrossRef] [Green Version]
- Schulter, D. Ecology and the origion of species. Trends Ecol. Evol. 2001, 16, 372–380. [Google Scholar] [CrossRef]
- Feder, J.L.; Berlocher, S.H.; Roethele, J.B.; Dambroski, H.; Smith, J.J.; Perry, W.L.; Gavrilovic, V.; Filchak, K.E.; Rull, J.; Aluja, M. Allopatric genetic origins for sympatric host-plant shifts and race formation in Rhagoletis. Proc. Nat. Acad. Sci. USA 2003, 100, 10314–10319. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
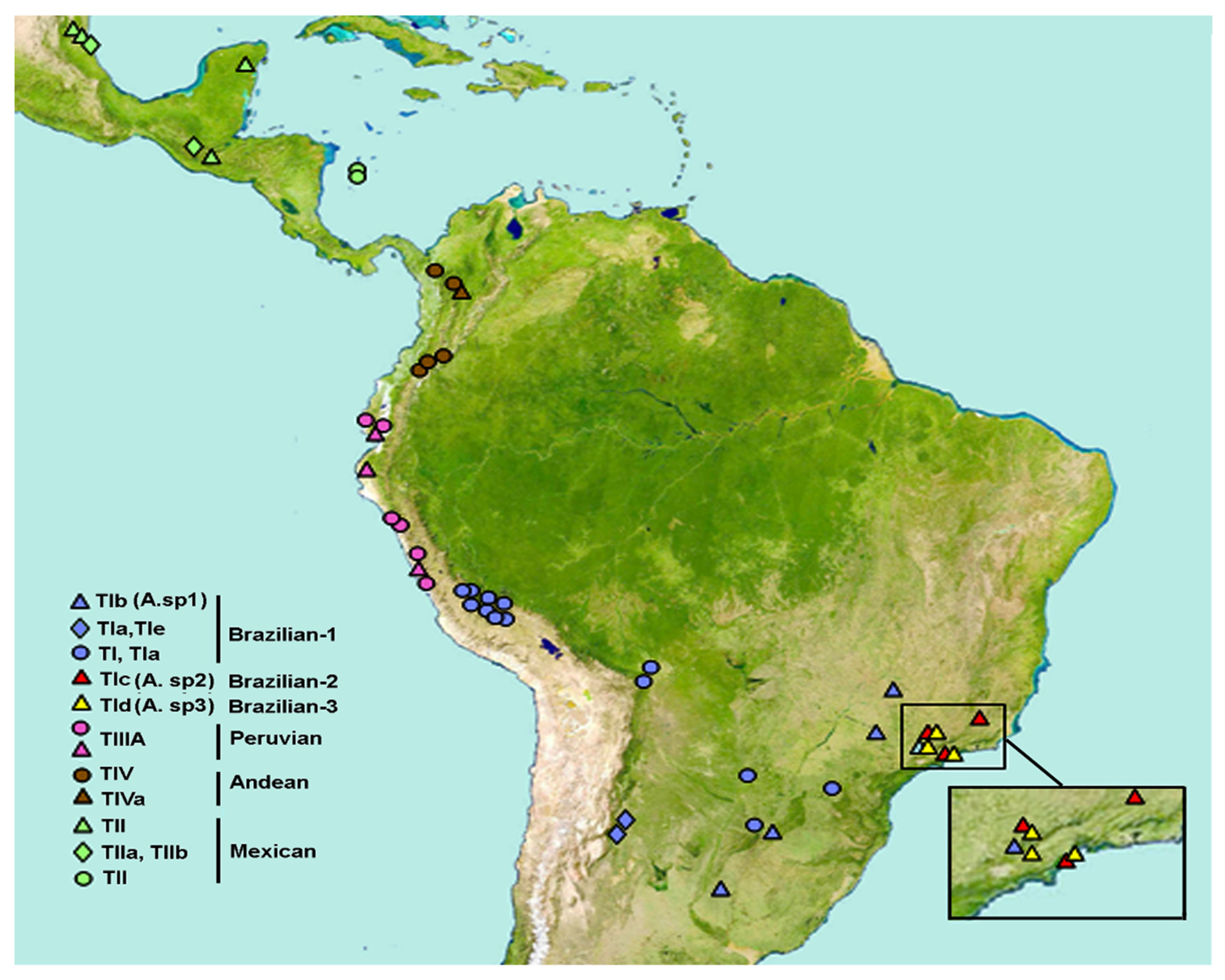{kind=link}
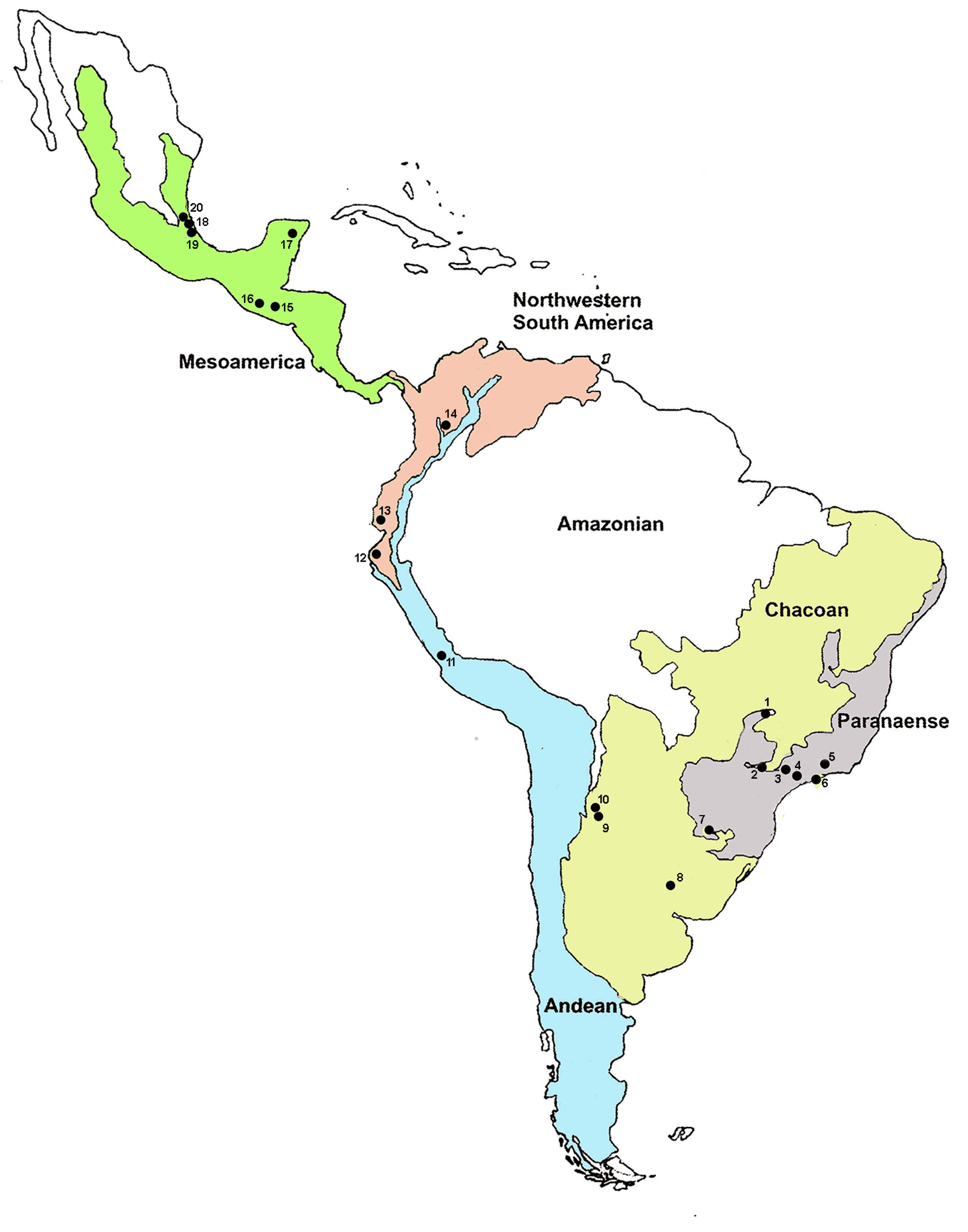
| Country | Provinces * | Locality | Map | Coordinates | Host Fruit |
|---|---|---|---|---|---|
| Brazil | Parana | Uberlândia | 1 | 18°56′46″ S, 48°13′55″ W | Guava (Psidium guajava) |
| Botucatu | 2 | 22°56′18″ S, 48°18′25″ W | Guava | ||
| Santa Isabel ** | 3 | 23°19′00″ S, 46°13′25″ W | Guava; Orange (Citrus sp.) | ||
| Parana Florest | Três Rios ** | 5 | 22°07′21″ S, 45°41′52″ W | Guava; Orange | |
| São Luis do Paraitinga ** | 4 | 23°13′24″ S, 45°18′47″ W | Guava | ||
| Atlantic | Ubatuba | 6 | 22°46′24″ S, 48°18′25″ W | Tropical almond (Teminalia cattapa) | |
| Argentina | Parana | Misiones | 7 | 27°23′59″ S, 55°56′01″ W | Guava |
| Pampeana | Concordia | 8 | 31°23′13″ S, 58°01′12″ W | Guava | |
| Chaco | Tucumán | 9 | 27°02′18″ S, 65°19′13″ W | Guava | |
| Horco Molle | 10 | 26°46′37″ S, 65°19′49″ W | Guava | ||
| Peru | Desert | La Molina | 11 | 12°14′15″ S, 76°31′50″ W | Lab colony (Annona cherimola) |
| Ecuadorian | Piura | 12 | 07°40′23″ S, 79°12′40″ W | Guava | |
| Ecuador | West Ecuadorian | Guayaquil | 13 | 02°2′13″ S, 79°53′50″ W | Guava |
| Colombia | Paramo | Ibagué | 14 | 04°26′11″ N, 5°11′29″ W | Lab colony (Coffea arabica) |
| Guatemala | High Altitude Chiapas | Guatemala City | 15 | 14°36′51″ N, 0°32′22″ W | Guava |
| Mexico | Pacific Low Land | Tapachula | 16 | 14°53′47″ N, 2°10′30″ W | Guava; Loquat (Eriobotrya japonica) |
| Yucatan | Quintana Roo | 17 | 19°37′39″ N, 8°38′56″ W | McPhail trap | |
| Veracruz | Teocelo | 18 | 19°23′14″ N, 6°57′23″ W | Guava | |
| Apazapan | 19 | 19°17′00″ N, 6°36′59″ W | McPhail trap | ||
| La Jicayana | 20 | 19°21′44″ N, 6°39′23″ W | Guava |
| Country | Locality | Code | Sample | NCBI Accession * |
|---|---|---|---|---|
| Brazil | Palmas, PR | BrPal | 9 | KY400289 |
| Uberlândia, MG | BrUbe | 1 | JN223390 | |
| Botucatú, SP | BrBot | 2 | JN223383 | |
| Santa Isabel, SP | BrIsa | 3 | JN223376; JN1223379; JN223385 | |
| S. L. Paraitinga, SP | BrSlp | 4 | JN223382 | |
| Três Rios, RJ | BrTri | 5 | JN223378 | |
| Ubatuba, SP | BrUba | 6 | JN223377; JN223380 | |
| Argentina | Misiones | ArMis | 7 | MH806409 |
| Concordia | ArCon | 8 | MH806412 | |
| Tucumán | ArTuc | 9 | MH806411 | |
| Horco Molle | ArHmo | 10 | MH806410 | |
| Bolivia | Santa Cruz | BoSCr | 3 | KT829864 |
| Los Volcanes | BoVol | 4 | KT400288 | |
| Peru (highlands) | Huatatas | PeHut | 63 | KY400324 |
| Pucahuasi | PePuc | 65 | KY400299 | |
| Huanta | PeHua | 66 | KY400311 | |
| Cusco | PeCus | 68 | KY400304 | |
| Sillacancha | PeSil | 70 | KY400306; KY400307 | |
| VRAE Qanan | PeVRq | 95 | KY400302 | |
| SENASA | PeSen | 96 | KY400309 | |
| VRAERegion | PeVRr | 97 | KY400310 | |
| Peru (lowlands) | La Molina | PeLmo | 11 | MH806421 |
| Piura | PePiu | 12 | MH806422 | |
| Casma | PeCas | 61 | KY400395 | |
| Cajamarca | PeCaj | 67 | KY400366 | |
| Lambayeque | PeLam | 80 | KY400367 | |
| Jequetepeque | PeJeq | 83 | KY400337 | |
| Tumbes | PeTum | 92 | KY400379 | |
| Ecuador | Guayas | EcGuy | 49 | KY400327; KT893867 |
| Guayaquil | EcGua | 13 | MH806420 | |
| Colombia (island) (Highlands) | San Andres | CoSan | 43; 44 | KY400484; KT893866 |
| Ibagué | CoIba | 14 | MH806419 | |
| Rionegro | CoAnt | 17 | KT893870 | |
| CoRio | 18;19 | KY400427; KY400425 | ||
| CoCau | 20 | KY400436 | ||
| Huila | CoHui | 24 | KY400468 | |
| Risaralda | CoRis | 40 | KY400476 | |
| Mexico | Chiapas | MxChi | 54 | KY400491 |
| Tapachula | MxTap | 16 | MH806414 | |
| Quintana Roo | MxQro | 17 | MH806413 | |
| Teocelo | MxTeo | 18 | MH806417 | |
| Apazapan | MxApa | 19 | MH806416 | |
| La Jicayana | MxJic | 20 | MH806415 | |
| Guatemala | Guatemala City | GuCit | 15 | MH806418 |
| Collection | Sequence | |||
|---|---|---|---|---|
| Morphotype | Country | Code | N | |
| Brazilian-1 | Brazil | Uberlândia | B1Ube | 10 |
| Botucatu | B1Bot | 10 | ||
| Santa Isabel | B1Isa | 10 | ||
| Argentina | Misiones | ArMis | 2 | |
| Concordia | ArCon | 3 | ||
| Horco-Molle | ArHmo | 3 | ||
| Tucumán | ArTuc | 3 | ||
| Brazilian-2 | Brazil | Santa Isabel | B2Isa | 10 |
| Tres Rios | B2Tri | 10 | ||
| Ubatuba | B2Uba | 10 | ||
| Brazilian-3 | Brazil | Santa Isabel | B3Isa | 10 |
| S.L.Paraitinga | B3Slp | 10 | ||
| Ubatuba | B3Uba | 10 | ||
| Peruvian | Peru | La Molina | PeLmo | 3 |
| Piura | PePiu | 2 | ||
| Ecuador | Guayaquil | EcGua | 10 | |
| Andean | Colombia | Ibagué | CoIba | 2 |
| Mexican | Guatemala | Guatemala City | GuCit | 2 |
| Mexico | Tapachula | MxTap | 2 | |
| Quintana-Roo | MxQro | 3 | ||
| Teocelo | MxTeo | 4 | ||
| Apazapan | MxApa | 2 | ||
| La Jicayana | MxJic | 2 | ||
| % Correct Classification | Bra-1 | Bra-2 | Bra-3 | And | Mex | Per | Observed | |
|---|---|---|---|---|---|---|---|---|
| Brazilian-1 | 98.8 | 85 | 0 | 1 | 0 | 0 | 0 | 86 |
| Brazilian-2 | 100.0 | 0 | 11 | 0 | 0 | 0 | 0 | 11 |
| Brazilian-3 | 100.0 | 0 | 0 | 21 | 0 | 0 | 0 | 21 |
| Andean | 100.0 | 0 | 0 | 0 | 15 | 0 | 0 | 15 |
| Mexican | 98.4 | 0 | 0 | 0 | 0 | 60 | 1 | 61 |
| Peruvian | 100.0 | 0 | 0 | 0 | 0 | 0 | 45 | 45 |
| Expected | 99.2 | 85 | 11 | 22 | 15 | 60 | 46 | 239 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prezotto, L.F.; Perondini, A.L.P.; Hernández-Ortiz, V.; Frías, D.; Selivon, D. What Can Integrated Analysis of Morphological and Genetic Data Still Reveal about the Anastrepha fraterculus (Diptera: Tephritidae) Cryptic Species Complex? Insects 2019, 10, 408. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10110408
Prezotto LF, Perondini ALP, Hernández-Ortiz V, Frías D, Selivon D. What Can Integrated Analysis of Morphological and Genetic Data Still Reveal about the Anastrepha fraterculus (Diptera: Tephritidae) Cryptic Species Complex? Insects. 2019; 10(11):408. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10110408
Chicago/Turabian StylePrezotto, Leandro F., André L.P. Perondini, Vicente Hernández-Ortiz, Daniel Frías, and Denise Selivon. 2019. "What Can Integrated Analysis of Morphological and Genetic Data Still Reveal about the Anastrepha fraterculus (Diptera: Tephritidae) Cryptic Species Complex?" Insects 10, no. 11: 408. https://0-doi-org.brum.beds.ac.uk/10.3390/insects10110408